
Entropy Rate Maps of Complex Excitable Dynamics in Cardiac Monolayers

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Experimental Setup
2.1. Dataset and Preprocessing
2.1.1. Signal Processing
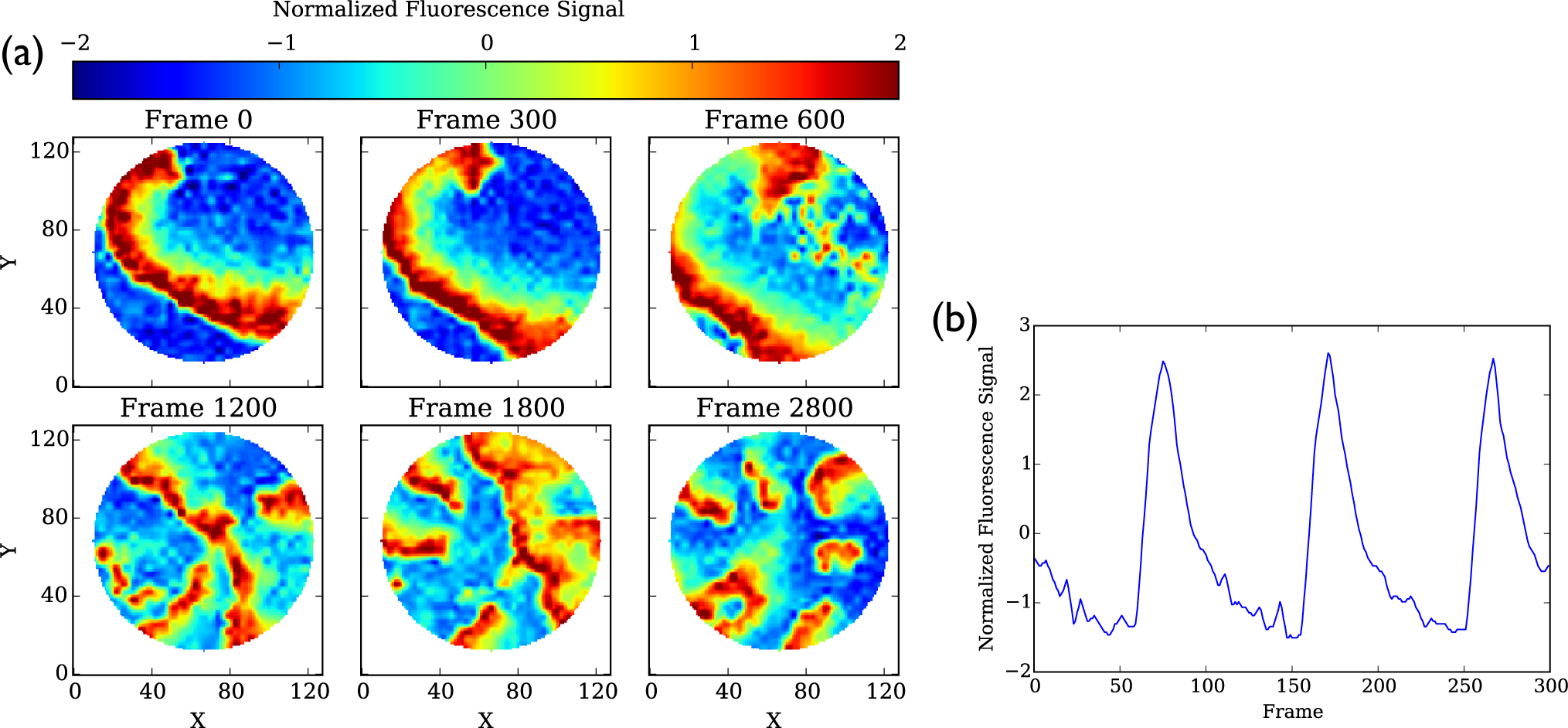
2.2. Normalization
2.3. Dominant Frequency Maps
2.4. Phase Singularity Analysis
2.5. Entropy of Binary Sequences
2.6. Entropy Rate Maps
3. Simulations of Excitable Media
3.1. Aliev–Panfilov Model
3.2. Cubic Barkley Model
4. Results and Discussion
- Simulated data using the Aliev–Panfilov model introduced in Section 3.1.
- Simulated data using the cubic Barkley model introduced in Section 3.2.
- The cell culture dataset described in Section 2.1.
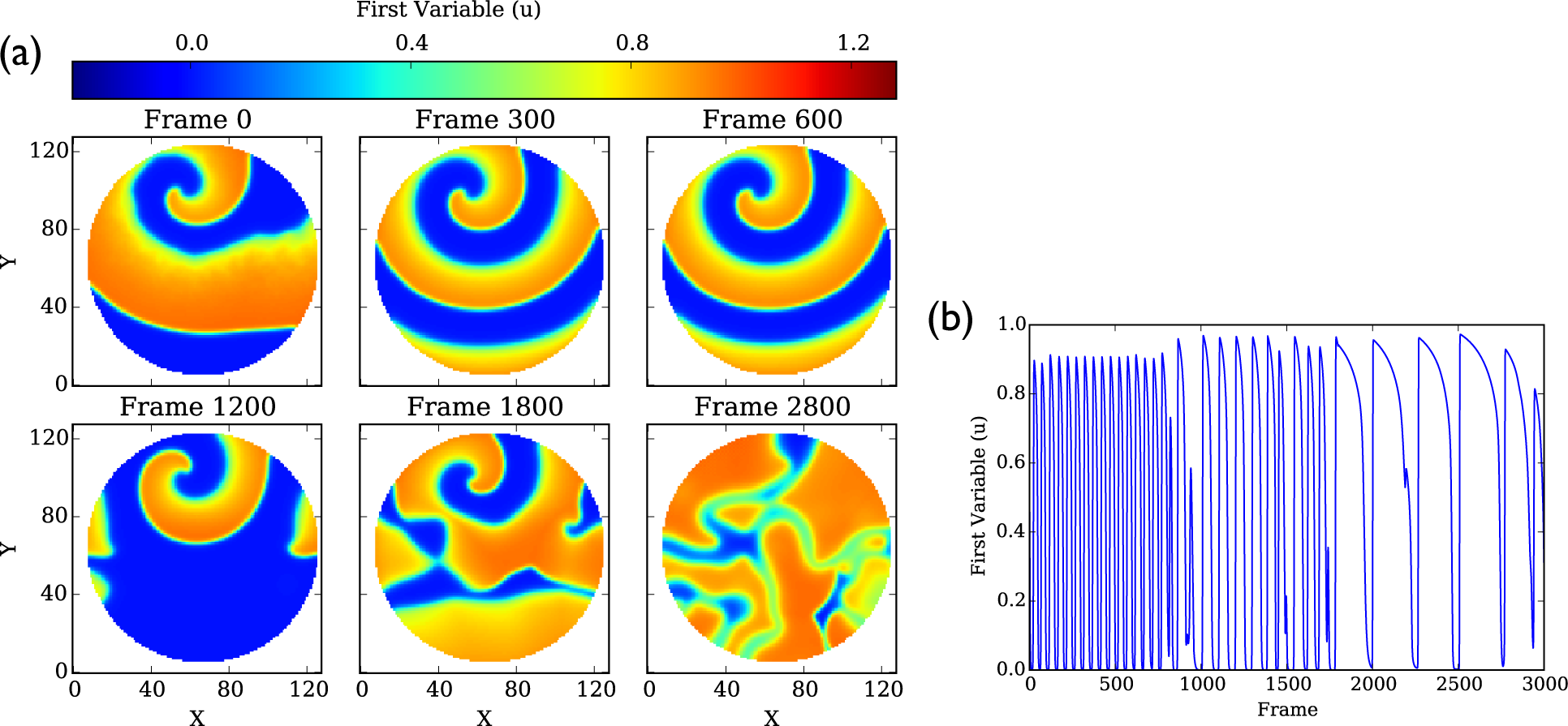
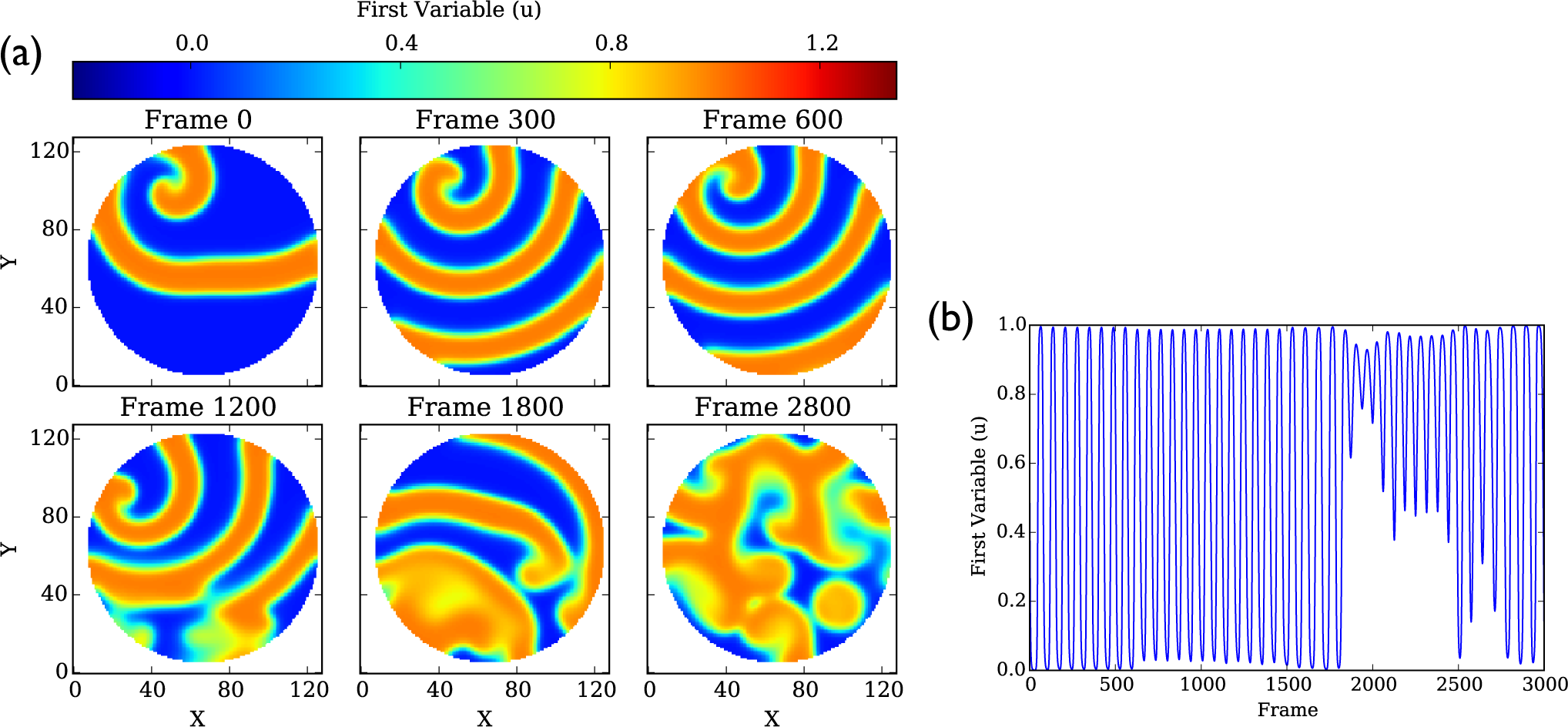
4.1. Aliev–Panfilov Model
4.1.1. Dominant Frequency Maps
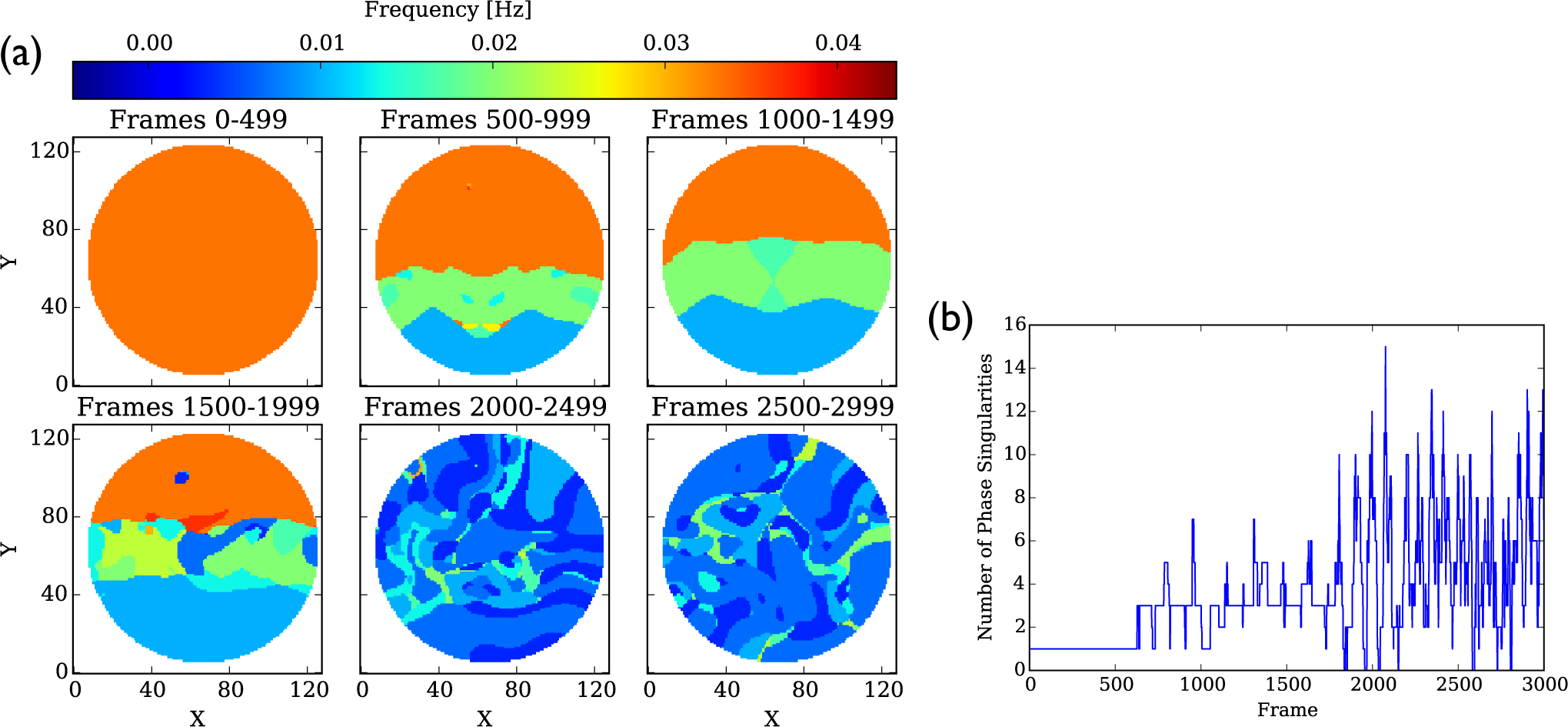
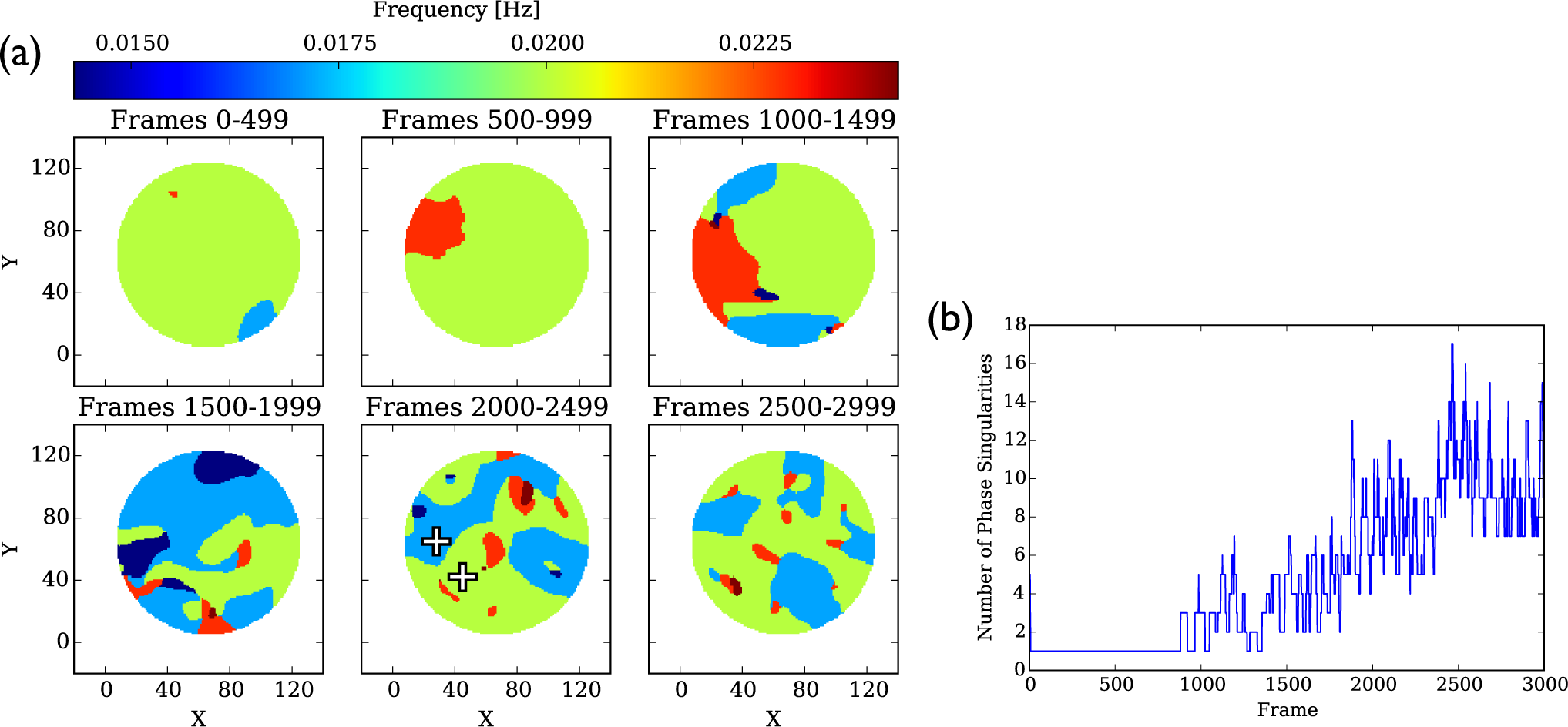
4.1.2. Phase Singularities
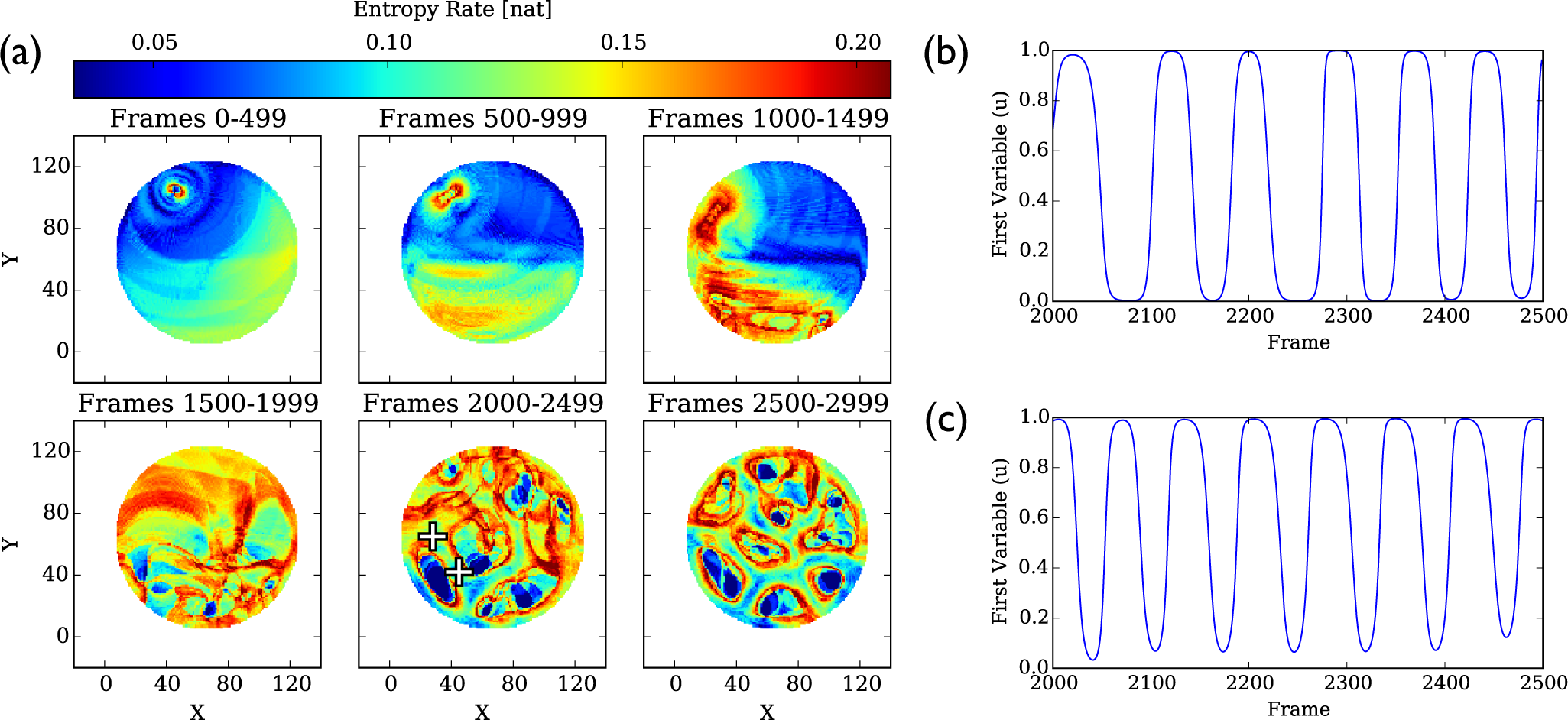
4.1.3. Entropy Rate Maps
4.2. Cubic Barkley Model
4.2.1. Dominant Frequency Maps
4.2.2. Phase Singularities
4.2.3. Entropy Rate Maps
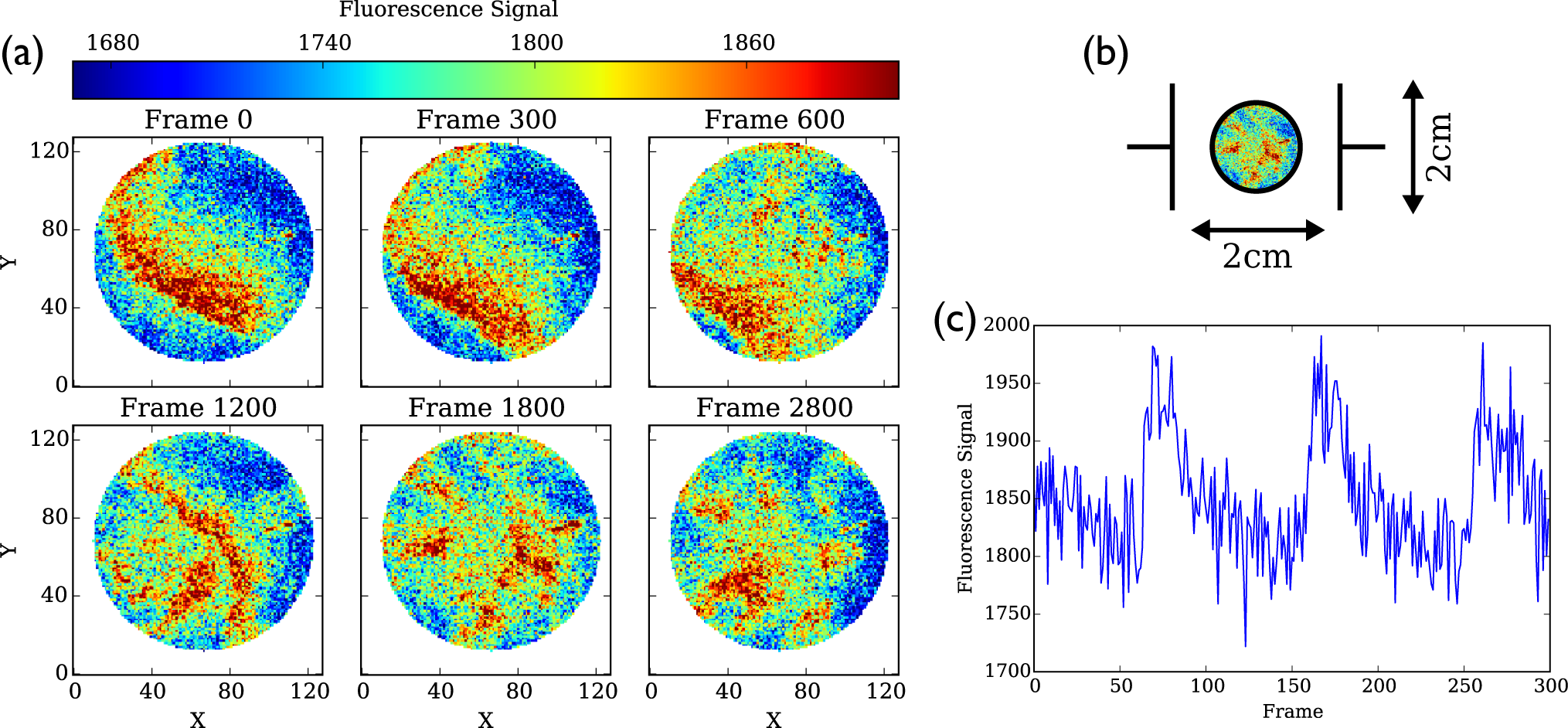
4.3. Cell Culture Data
4.3.1. Dominant Frequency Maps
4.3.2. Phase Singularities
4.3.3. Entropy Rate Maps
4.4. Comparison of Entropy Rate Maps and Dominant Frequency Maps
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
A. Appendix
A.1. Preparation of Cardiac Monolayers
A.1. Optical Mapping
References
- Cherry, E.M.; Fenton, F.H. Visualization of spiral and scroll waves in simulated and experimental cardiac tissue. New J. Phys. 2008, 10, 125016. [Google Scholar] [CrossRef]
- Parlitz, U.; Schlemmer, A.; Luther, S. Synchronization patterns in transient spiral wave dynamics. Phys. Rev. E 2011, 83, 057201. [Google Scholar] [CrossRef]
- Gray, R.A.; Pertsov, A.M.; Jalife, J. Spatial and temporal organization during cardiac fibrillation. Nature 1998, 392, 75–78. [Google Scholar]
- Borek, B.; Shajahan, T.K.; Gabriels, J.; Hodge, A.; Glass, L.; Shrier, A. Pacemaker interactions induce reentrant wave dynamics in engineered cardiac culture. Chaos 2012, 22, 033132. [Google Scholar]
- Shajahan, T.; Krinski, V.; Knyazeva, S.; Luther, S. Eliminating pinned spiral waves in cardiac monolayer by far field pacing. Proceedings of the 2014 8th Conference of the European Study Group on Cardiovascular Oscillations (ESGCO), Trento, Italy, 25–28 May 2014; pp. 151–152.
- Press, W.H.; Teukolsky, S.A.; Vetterling, W.T.; Flannery, B.P. Numerical Recipes: The Art of Scientific Computing, 3rd ed; Cambridge University Press: New York, NY, USA, 2007. [Google Scholar]
- Pandit, S.V.; Jalife, J. Rotors and the Dynamics of Cardiac Fibrillation. Circ. Res. 2013, 112, 849–862. [Google Scholar]
- Herlin, A.; Jacquemet, V. Reconstruction of phase maps from the configuration of phase singularities in two-dimensional manifolds. Phys. Rev. E 2012, 85, 051916. [Google Scholar]
- Iyer, A.N.; Gray, R.A. An Experimentalist’s Approach to Accurate Localization of Phase Singularities during Reentry. Ann. Biomed. Eng. 2001, 29, 47–59. [Google Scholar]
- Rogers, J. Combined phase singularity and wavefront analysis for optical maps of ventricular fibrillation. IEEE Bio-Med. Eng. 2004, 51, 56–65. [Google Scholar]
- Olkkonen, H. Computation of hilbert transform via discrete cosine transform. J. Signal Inf. Process. 2010, 1, 18–23. [Google Scholar]
- Schürmann, T.; Grassberger, P. Entropy estimation of symbol sequences. Chaos 1996, 6, 414–427. [Google Scholar]
- Faure, P.; Lesne, A. Estimating Kolmogorov Entropy from Recurrence Plots. In Recurrence Quantification Analysis; Webber, C.L., Jr., Marwan, N., Eds.; Understanding Complex Systems; Springer: Berlin/Heidelberg, Germany, 2015; pp. 45–63. [Google Scholar]
- pyEntropyRateMap A Python Algorithm for Computing Entropy Rate Maps. Available online: http://www.bmp.ds.mpg.de/software.html accessed on 15 February 2015.
- Jung, P. Coherent structure analysis of spatiotemporal chaos. Phys. Rev. E 2000, 61, 2095–2098. [Google Scholar]
- Bub, G.; Shrier, A.; Glass, L. Global organization of dynamics in oscillatory heterogeneous excitable media. Phys. Rev. Lett. 2005, 94, 028105. [Google Scholar]
- Aliev, R.R.; Panfilov, A.V. A Simple Two-variable Model of Cardiac Excitation. Chaos Solitons Fractals 1996, 7, 293–301. [Google Scholar]
- Fenton, F.H.; Cherry, E.M.; Karma, A.; Rappel, W.J. Modeling wave propagation in realistic heart geometries using the phase-field method. Chaos 2005, 15, 013502. [Google Scholar]
- Barkley, D. A model for fast computer simulation of waves in excitable media. Physica D 1991, 49, 61–70. [Google Scholar]
- Barkley, D. Barkley model. Scholarpedia 2008, 3. [Google Scholar] [CrossRef]
- MultiRecorder, a Multiple-Camera Recording and Analysis Software. Available online: http://www.bmp.ds.mpg.de/multirecorder.html accessed on 6 October 2014.











© 2015 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schlemmer, A.; Berg, S.; Shajahan, T.K.; Luther, S.; Parlitz, U. Entropy Rate Maps of Complex Excitable Dynamics in Cardiac Monolayers. Entropy 2015, 17, 950-967. https://doi.org/10.3390/e17030950
Schlemmer A, Berg S, Shajahan TK, Luther S, Parlitz U. Entropy Rate Maps of Complex Excitable Dynamics in Cardiac Monolayers. Entropy. 2015; 17(3):950-967. https://doi.org/10.3390/e17030950
Chicago/Turabian StyleSchlemmer, Alexander, Sebastian Berg, T. K. Shajahan, Stefan Luther, and Ulrich Parlitz. 2015. "Entropy Rate Maps of Complex Excitable Dynamics in Cardiac Monolayers" Entropy 17, no. 3: 950-967. https://doi.org/10.3390/e17030950
APA StyleSchlemmer, A., Berg, S., Shajahan, T. K., Luther, S., & Parlitz, U. (2015). Entropy Rate Maps of Complex Excitable Dynamics in Cardiac Monolayers. Entropy, 17(3), 950-967. https://doi.org/10.3390/e17030950