Polygenic Adaptation in a Population of Finite Size


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Model and Analysis
2.1. Deterministic Model of a Single Quantitative Trait
2.2. Stochastic Analysis
3. Results
3.1. Stochastic Equilibrium between Drift, Mutation and Selection
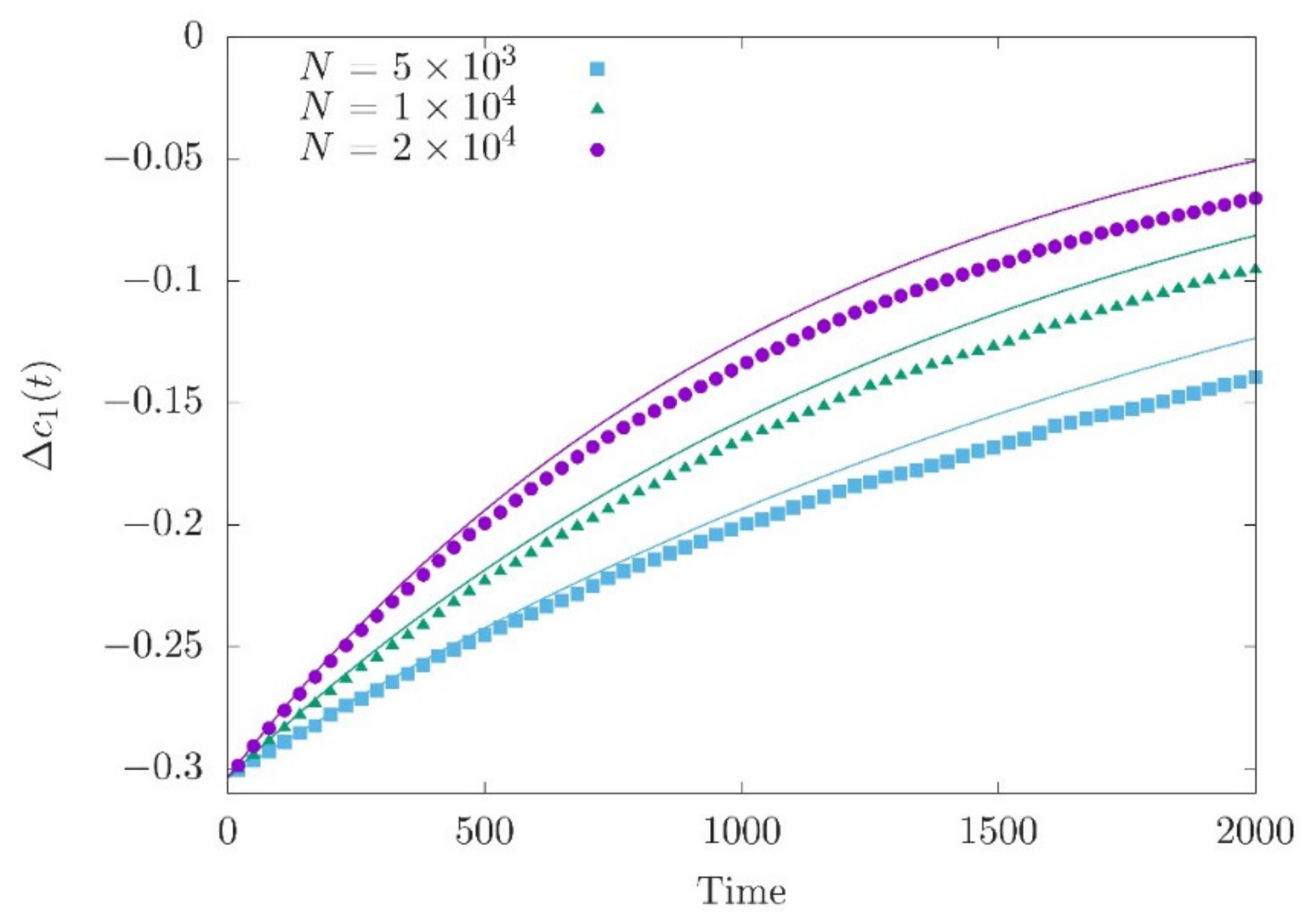
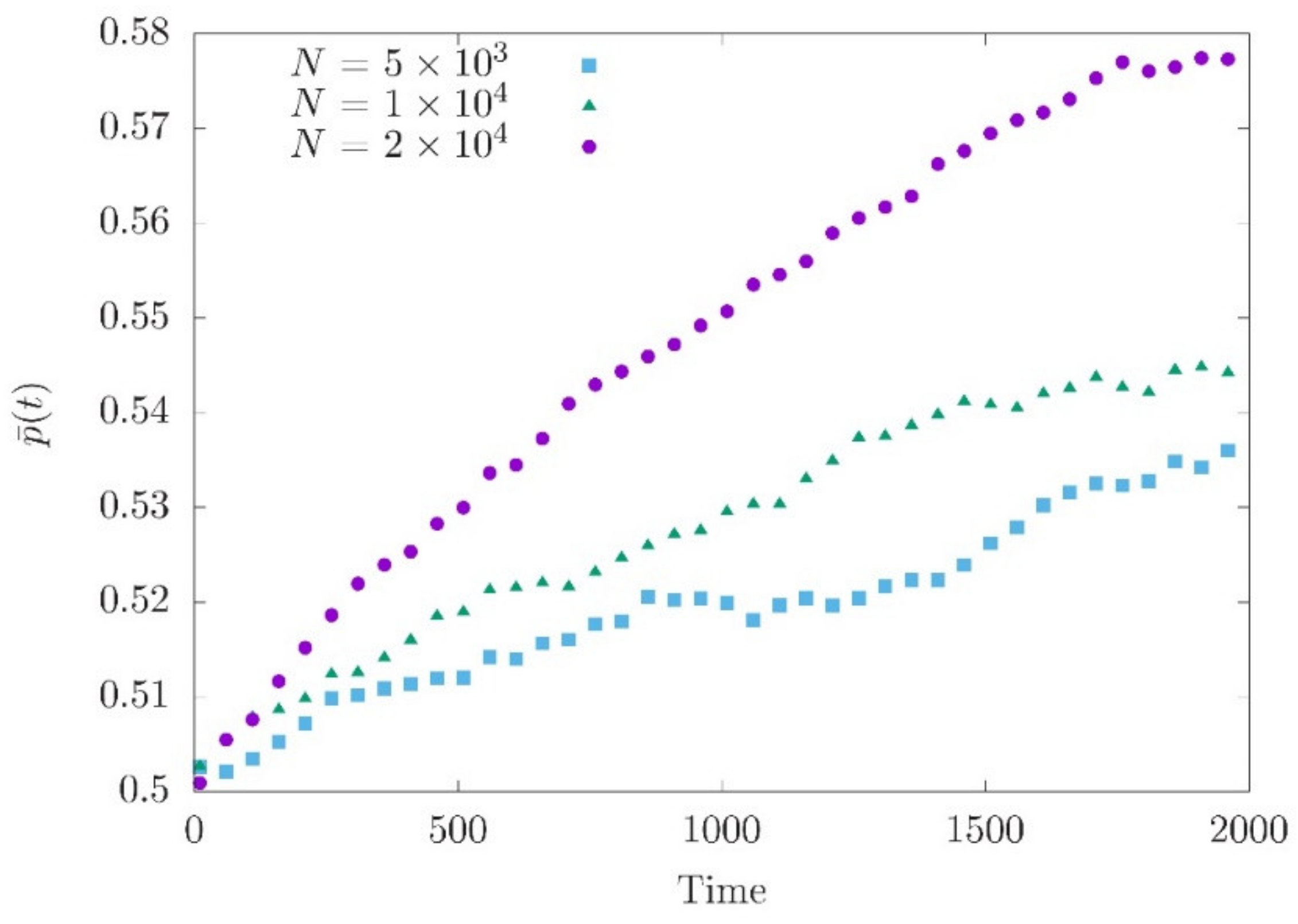
3.2. Adaptation after a Sudden Shift of the Fitness Optimum
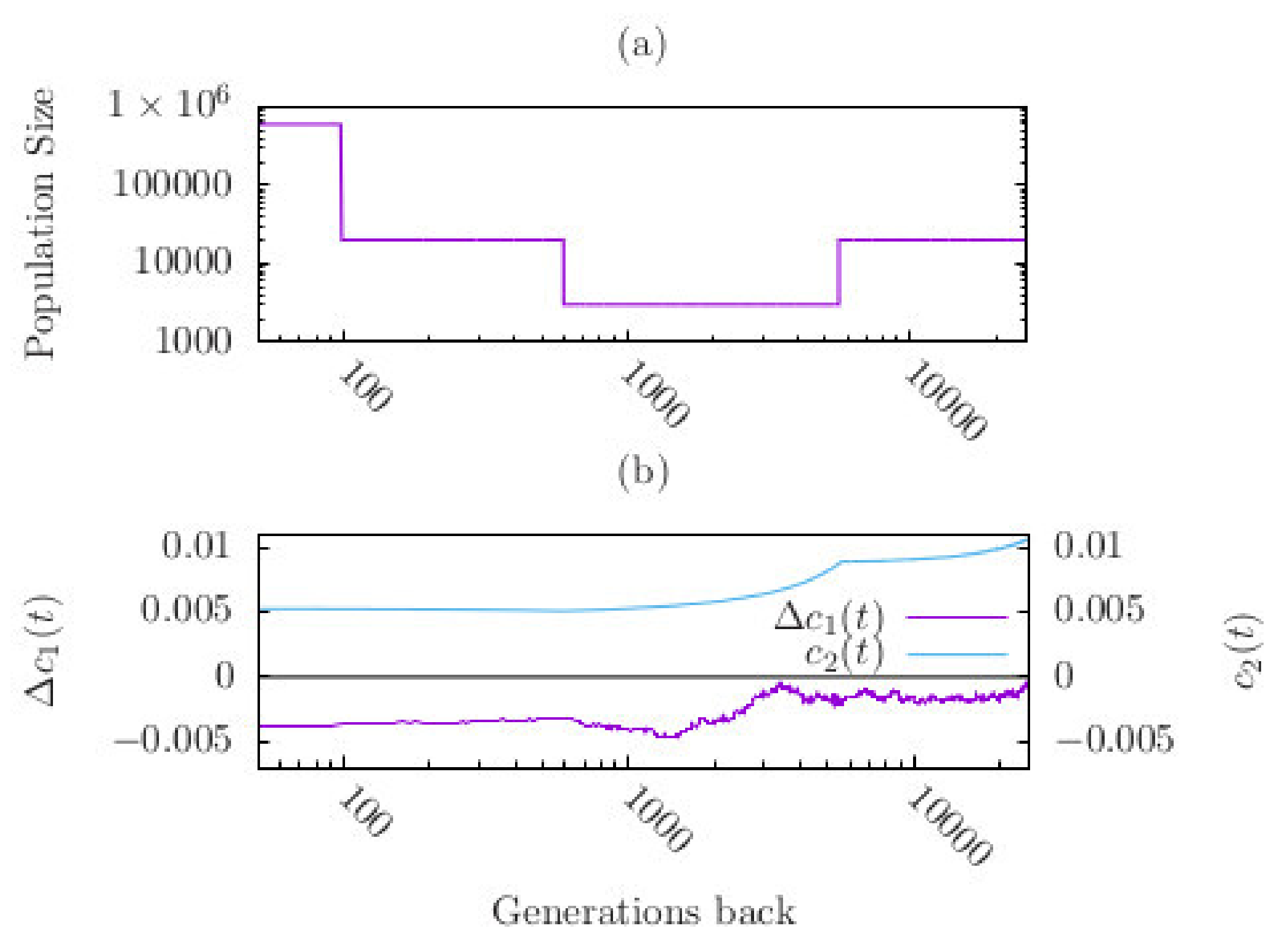
3.3. Effects of Demography on Polygenic Adaptation
4. Discussion
4.1. Summary
4.2. Impact of Large-Effect Alleles
4.3. Do Selective Sweeps Occur in Polygenic Adaptation?
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Akey, J.M.; Eberle, M.A.; Rieder, M.J.; Carlson, C.S.; Shriver, M.D.A.; Nickerson, D.; Kruglyak, L. Population history and natural selection shape patterns of genetic variation in 132 Genes. PLoS Biol. 2004, 2, e286. [Google Scholar] [CrossRef]
- Berg, J.J.; Coop, G. A population genetic signal of polygenic adaptation. PLoS Genet. 2014, 10, e1004412. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.; Stephan, W. Detecting a local signature of genetic hitchhiking along a recombining chromosome. Genetics 2002, 160, 765–777. [Google Scholar] [CrossRef]
- Nielsen, R.; Williamson, S.; Kim, Y.; Hubisz, M.J.; Clark, A.G.; Bustamante, C. Genomic scans for selective sweeps using SNP data. Genome Res. 2005, 15, 1566–1575. [Google Scholar] [CrossRef] [Green Version]
- Kaplan, N.L.; Hudson, R.R.; Langley, C.H. The ‘hitchhiking effect’ revisited. Genetics 1989, 123, 887–899. [Google Scholar]
- Maynard Smith, J.; Haigh, J. The hitch-hiking effect of a favourable gene. Genet. Res. 1974, 23, 23–35. [Google Scholar] [CrossRef] [Green Version]
- Stephan, W.; Wiehe, T.H.E.; Lenz, M.W. The effect of strongly selected substitutions on neutral polymorphism: Analytical results based on diffusion theory. Theor. Popul. Biol. 1992, 41, 237–254. [Google Scholar] [CrossRef]
- Barton, N.H. Linkage and the limits to natural selection. Genetics 1995, 140, 821–841. [Google Scholar]
- Chevin, L.M.; Billiard, S.; Hospital, F. Hitchhiking both ways: Effect of two interfering selective sweeps on linked neutral variation. Genetics 2008, 180, 301–316. [Google Scholar] [CrossRef]
- Kirby, D.A.; Stephan, W. Multi-locus selection and the structure of variation at the white gene of Drosophila melanogaster. Genetics 1996, 144, 635–645. [Google Scholar]
- Mackay, T.F.C. The genetic architecture of quantitative traits: Lessons from Drosophila. Curr. Opin. Genet. Dev. 2004, 14, 253–257. [Google Scholar] [CrossRef] [PubMed]
- Catalan, A.; Glaser-Schmitt, A.; Argyridou, E.; Duchen, P.; Parsch, J. An indel polymorphism in the MtnA 3′ untranslated region is associated with gene expression variation and local adaptation in Drosophila melanogaster. PLoS Genet. 2016, 12, e1005987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lange, B.W.; Langley, C.H.; Stephan, W. Molecular evolution of Drosophila metallothionein genes. Genetics 1990, 126, 921–932. [Google Scholar] [PubMed]
- Pfeifer, S.P.; Laurent, S.; Sousa, V.C.; Linnen, C.R.; Foll, M.; Excoffier, L.; Hoekstra, H.E.; Jensen, J.D. The evolutionary history of Nebraska deer mice: Local adaptation in the face of strong gene flow. Mol. Biol. Evol. 2018, 35, 792–806. [Google Scholar] [CrossRef]
- Slatkin, M.; Wiehe, T. Genetic hitch-hiking in a subdivided population. Genet. Res. 1998, 71, 155–160. [Google Scholar] [CrossRef] [Green Version]
- Stephan, W. Selective sweeps. Genetics 2019, 211, 5–13. [Google Scholar] [CrossRef] [Green Version]
- Pritchard, J.K.; Pickrell, J.K.; Coop, G. The genetics of human adaptation: Hard sweeps, soft sweeps, and polygenic adaptation. Curr. Biol. 2010, 20, R208–R215. [Google Scholar] [CrossRef] [Green Version]
- Pritchard, J.K.; Di Rienzo, A. Adaptation—not by sweeps alone. Nat. Rev. Genet. 2010, 11, 665–667. [Google Scholar] [CrossRef] [Green Version]
- Reznick, D.N. The Origin Then and Now: An Interpretive Guide to the Origin of Species; Princeton University Press: Princeton, NJ, USA, 2009. [Google Scholar]
- Vignieri, S.N.; Larson, J.G.; Hoekstra, H.E. The selective advantage of crypsis in mice. Evolution 2010, 64, 2153–2158. [Google Scholar] [CrossRef]
- Cook, L.M.; Grant, B.S.; Saccheri, I.J.; Mallet, J. Selective bird predation on the peppered moth: The last experiment of Michael Majerus. Biol. Lett. 2012, 8, 609–612. [Google Scholar] [CrossRef]
- Ffrench-Constant, R.H.; Bogwitz, M.; Daborne, P.; Yen, J. A single P450 allele associated with insecticide resistance in Drosophila. Science 2002, 297, 2253–2256. [Google Scholar] [CrossRef]
- Grant, P.R.; Grant, B.R. How and Why Species Multiply: The Radiation of Darwin’s Finches; Princeton University Press: Princeton, NJ, USA, 2008. [Google Scholar]
- Losos, J.B. Lizards in an Evolutionary Tree: Ecology and Adaptive Radiation of Anoles; University of California Press: San Francisco, CA, USA, 2009. [Google Scholar]
- Van’t Hof, A.E.; Edmonds, N.; Dalíková, M.; Marec, F.; Saccheri, I.J. Industrial melanism in British peppered moths has a singular and recent mutational origin. Science 2011, 332, 958–960. [Google Scholar] [CrossRef] [Green Version]
- Turchin, M.C.; Chiang, C.W.K.; Palmer, C.D.; Sankararaman, S.; Reich, D.; Genetic Investigation of ANthropometric Traits (GIANT) Consortium; Hirschhorn, J.N. Evidence of widespread selection on standing variation in Europe at height-associated SNPs. Nat. Genet. 2012, 44, 1015–1019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bürger, R.; Lynch, M. Evolution and extinction in a changing environment: A quantitative-genetic analysis. Evolution 1995, 49, 151–163. [Google Scholar] [CrossRef]
- Lande, R. Natural selection and random genetic drift in phenotypic evolution. Evolution 1976, 30, 314–334. [Google Scholar] [CrossRef]
- De Vladar, H.P.; Barton, N. Stability and response of polygenic traits to stabilizing selection and mutation. Genetics 2014, 197, 749–767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jain, K.; Stephan, W. Response of polygenic traits under stabilizing selection and mutation when loci have unequal effects. G3 Genes Genomes Genet. 2015, 5, 1065–1074. [Google Scholar] [CrossRef] [Green Version]
- Jain, K.; Stephan, W. Rapid adaptation of a polygenic trait after a sudden environmental shift. Genetics 2017, 206, 389–406. [Google Scholar] [CrossRef]
- Jain, K.; Stephan, W. Modes of rapid polygenic adaptation. Mol. Biol. Evol. 2017, 34, 3169–3175. [Google Scholar] [CrossRef]
- Höllinger, I.; Pennings, P.S.; Hermisson, J. Polygenic adaptation: From sweeps to subtle frequency shifts. PLoS Genet. 2019, 15, e1008035. [Google Scholar] [CrossRef] [Green Version]
- John, S.; Stephan, W. Important role of genetic drift in rapid polygenic adaptation. Ecol. Evol. 2020, 10, 1278–1287. [Google Scholar] [CrossRef] [PubMed]
- Simons, Y.B.; Bullaughey, K.; Hudson, R.R.; Sella, G. A population genetic interpretation of GWAS findings for human quantitative traits. PLoS Biol. 2018, 16, e2002985. [Google Scholar] [CrossRef] [PubMed]
- Stetter, M.G.; Thornton, K.R.; Ross-Ibarra, J. Genetic architecture and selective sweeps after polygenic adaptation to distant trait optima. PLoS Genet. 2018, 14, e1007794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thornton, K.R. Polygenic adaptation to an environmental shift: Temporal dynamics of variation under Gaussian stabilizing selection and additive effects on a single trait. Genetics 2019, 213, 1513–1530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, S. Evolution in populations in approximate equilibrium. J. Genet. 1935, 30, 257–266. [Google Scholar] [CrossRef]
- Barton, N.H. The maintenance of polygenic variation through a balance between mutation and stabilizing selection. Genet. Res. 1986, 47, 209–216. [Google Scholar] [CrossRef] [Green Version]
- Bulmer, M.G. The genetic variability of polygenic characters under optimizing selection, mutation and drift. Genet. Res. 1972, 19, 17–25. [Google Scholar] [CrossRef]
- Ewens, W.J. Mathematical Population Genetics. I. Theoretical Introduction, 2nd ed.; Springer: Berlin, Germany, 2004. [Google Scholar]
- Gardiner, C.W. Handbook of Stochastic Methods, 7th ed.; Springer: Berlin, Germany, 1990. [Google Scholar]
- Schiffels, S.; Durbin, R. Inferring human population size and separation history from multiple genome sequences. Nat. Genet. 2014, 46, 919–925. [Google Scholar] [CrossRef] [Green Version]
- Hayward, L.K.; Sella, G. Polygenic adaptation after a sudden change in environment. bioRxiv 2019. [Google Scholar] [CrossRef]
- Chevin, L.M.; Hospital, F. Selective sweep at a quantitative trait locus in the presence of background genetic variation. Genetics 2008, 180, 1645–1660. [Google Scholar] [CrossRef] [Green Version]
- Pavlidis, P.; Metzler, D.; Stephan, W. Selective sweeps in multi-locus models of quantitative traits. Genetics 2012, 192, 225–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wollstein, A.; Stephan, W. Adaptive fixation in two-locus models of stabilizing selection and genetic drift. Genetics 2014, 198, 685–697. [Google Scholar] [CrossRef] [PubMed]
- Stephan, W. Signatures of positive selection: From selective sweeps at individual loci to subtle allele frequency changes in polygenic adaptation. Mol. Ecol. 2015, 25, 79–88. [Google Scholar] [CrossRef] [PubMed]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stephan, W.; John, S. Polygenic Adaptation in a Population of Finite Size. Entropy 2020, 22, 907. https://doi.org/10.3390/e22080907
Stephan W, John S. Polygenic Adaptation in a Population of Finite Size. Entropy. 2020; 22(8):907. https://doi.org/10.3390/e22080907
Chicago/Turabian StyleStephan, Wolfgang, and Sona John. 2020. "Polygenic Adaptation in a Population of Finite Size" Entropy 22, no. 8: 907. https://doi.org/10.3390/e22080907
APA StyleStephan, W., & John, S. (2020). Polygenic Adaptation in a Population of Finite Size. Entropy, 22(8), 907. https://doi.org/10.3390/e22080907