
Network Dynamics in Elemental Assimilation and Metabolism


 , , ,
, , ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Laser Ablation-Inductively Coupled Plasma-Mass Spectrometry
2.3. Recurrence Quantification Analysis
2.4. Graph Construction and Network Analysis
2.5. Statistical Analysis
3. Results
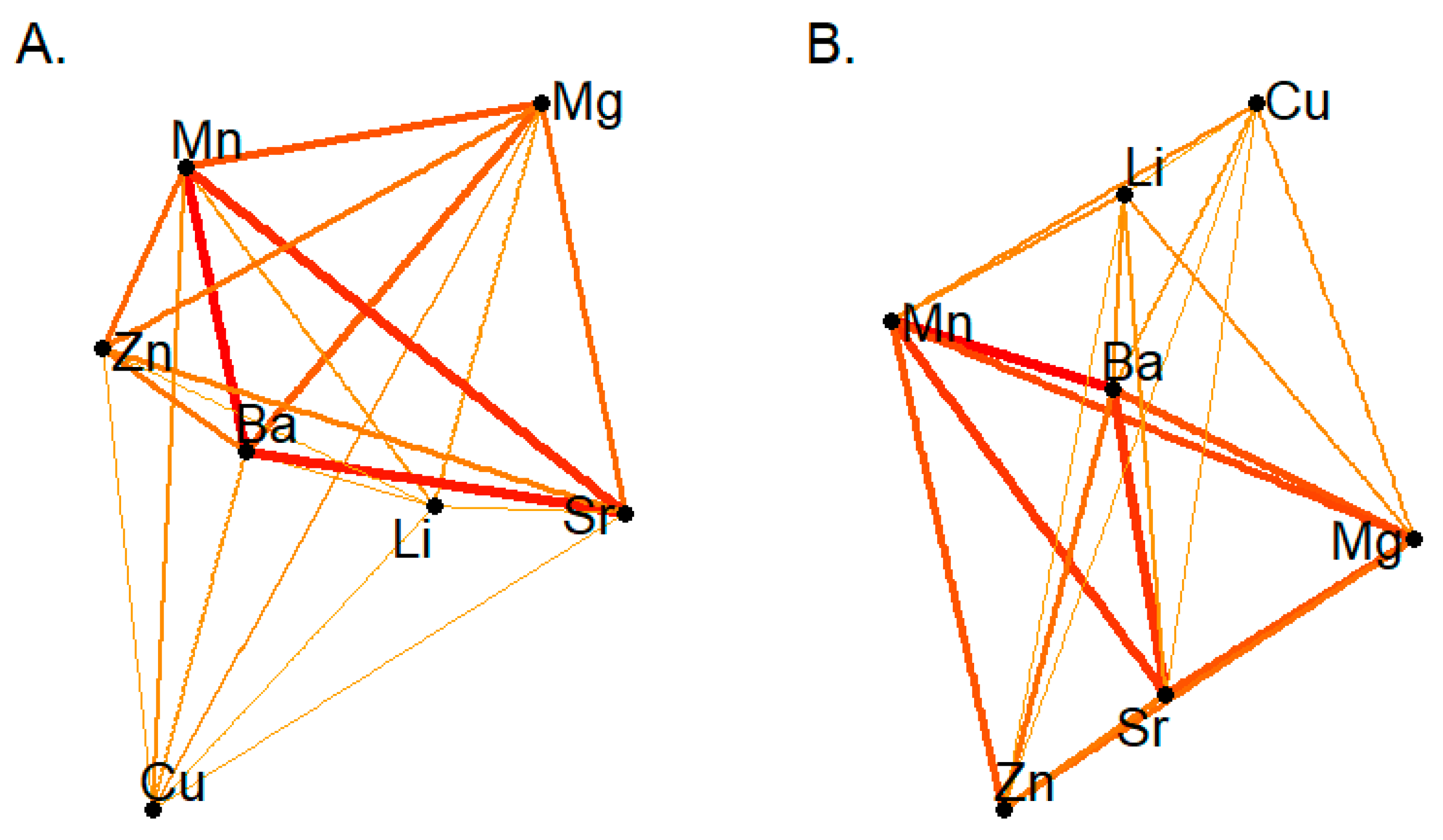
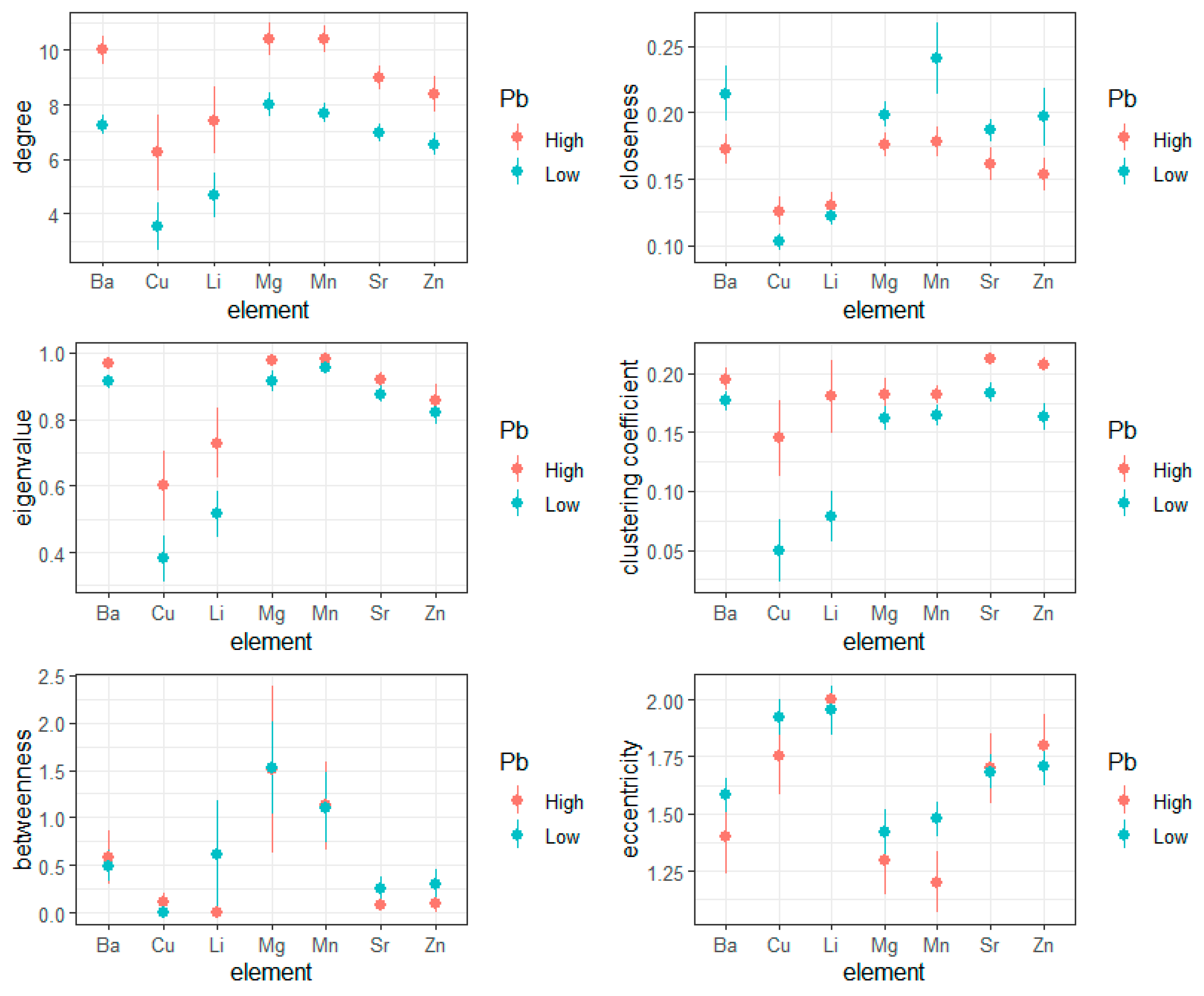
3.1. Network Dynamics in Recurrence Rates
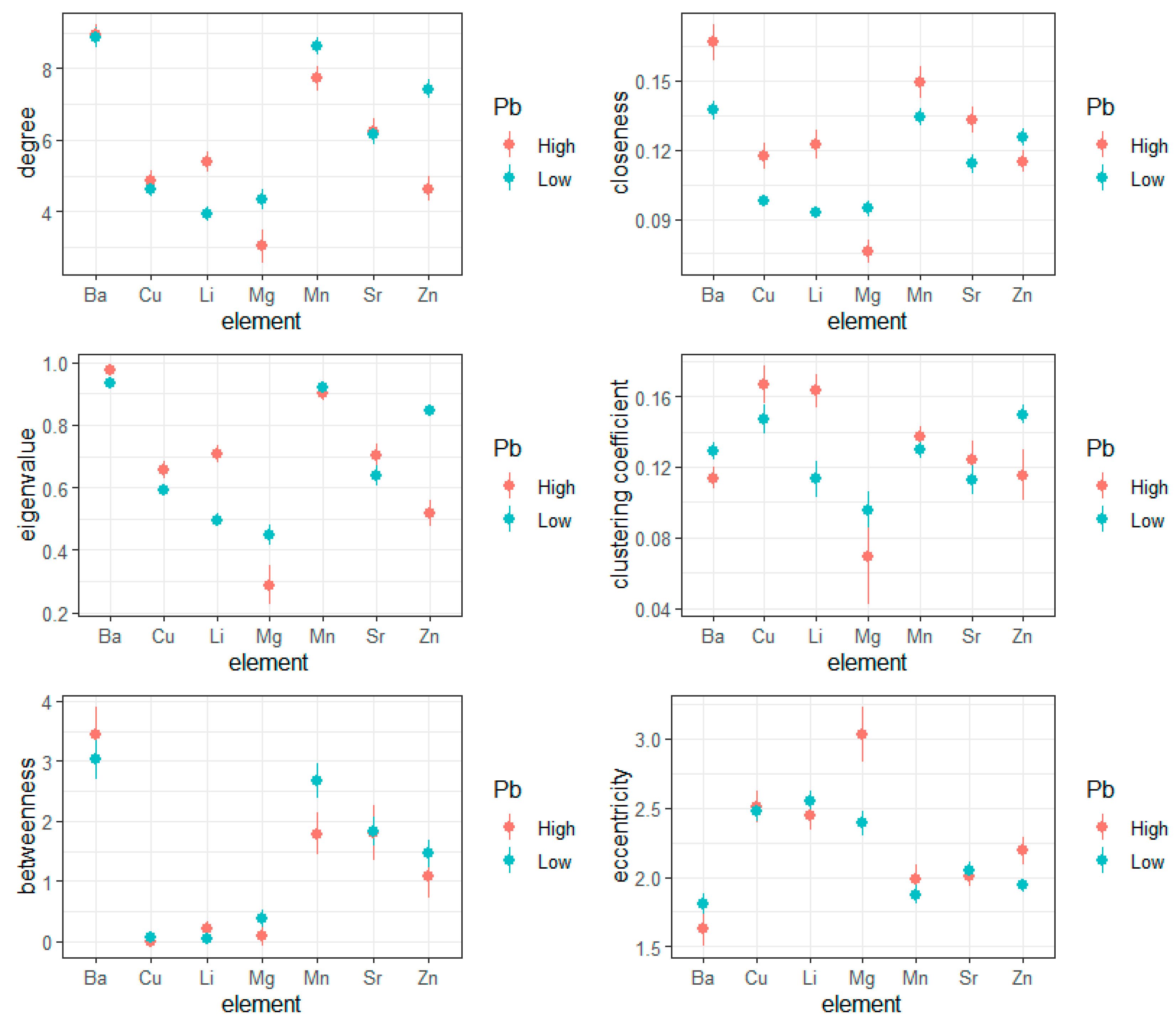
3.2. Entropy in Elemental Metabolic Networks
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Merced-Nieves, F.M.; Arora, M.; Wright, R.O.; Curtin, P. Metal mixtures and neurodevelopment: Recent findings and emerging principles. Curr. Opin. Toxicol. 2021, 26, 28–32. [Google Scholar] [CrossRef]
- Gluckman, P.D.; Hanson, M.A.; Spencer, H.G.; Bateson, P. Environmental influences during development and their later consequences for health and disease: Implications for the interpretation of empirical studies. Proc. Biol. Sci. 2005, 272, 671–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gluckman, P.D.; Hanson, M.A.; Cooper, C.; Thornburg, K.L. Effect of in utero and early-life conditions on adult health and disease. N. Engl. J. Med. 2008, 359, 61–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langley-Evans, S.C. Nutrition in early life and the programming of adult disease: A review. J. Hum. Nutr. Diet. 2015, 28 (Suppl. S1), 1–14. [Google Scholar] [CrossRef] [PubMed]
- Lucas, A.; Fewtrell, M.S.; Cole, T.J. Fetal origins of adult disease-the hypothesis revisited. BMJ 1999, 319, 245–249. [Google Scholar] [CrossRef]
- Arora, M.; Giuliani, A.; Curtin, P. Biodynamic Interfaces Are Essential for Human-Environment Interactions. Bioessays 2020, 42, e2000017. [Google Scholar] [CrossRef]
- Arora, M.; Austin, C. Teeth as a biomarker of past chemical exposure. Curr. Opin. Pediatr. 2013, 25, 261–267. [Google Scholar] [CrossRef]
- Curtin, P.; Curtin, A.; Austin, C.; Gennings, C.; Tammimies, K.; Bolte, S.; Arora, M. Recurrence quantification analysis to characterize cyclical components of environmental elemental exposures during fetal and postnatal development. PLoS ONE 2017, 12, e0187049. [Google Scholar] [CrossRef] [Green Version]
- Austin, C.; Curtin, P.; Curtin, A.; Gennings, C.; Arora, M.; Tammimies, K.; Isaksson, J.; Willfors, C.; Bolte, S. Dynamical properties of elemental metabolism distinguish attention deficit hyperactivity disorder from autism spectrum disorder. Transl. Psychiatry 2019, 9, 238. [Google Scholar] [CrossRef]
- Curtin, P.; Austin, C.; Curtin, A.; Gennings, C.; Arora, M.; Tammimies, K.; Willfors, C.; Berggren, S.; Siper, P.; Rai, D.; et al. Dynamical features in fetal and postnatal zinc-copper metabolic cycles predict the emergence of autism spectrum disorder. Sci. Adv. 2018, 4, eaat1293. [Google Scholar] [CrossRef] [Green Version]
- El-Sayed, A.M.; Scarborough, P.; Seemann, L.; Galea, S. Social network analysis and agent-based modeling in social epidemiology. Epidemiol. Perspect. Innov. 2012, 9, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kestens, Y.; Wasfi, R.; Naud, A.; Chaix, B. “Contextualizing Context”: Reconciling Environmental Exposures, Social Networks, and Location Preferences in Health Research. Curr. Environ. Health Rep. 2017, 4, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Phillips, R.D.; Wilson, S.M.; Sun, D.; Workgroup, V.A.M.-A.M.; Morey, R. Posttraumatic Stress Disorder Symptom Network Analysis in U.S. Military Veterans: Examining the Impact of Combat Exposure. Front. Psychiatry 2018, 9, 608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thakar, J.; Thatcher, T.H.; Smith, M.R.; Woeller, C.F.; Walker, D.I.; Utell, M.J.; Hopke, P.K.; Mallon, T.M.; Krahl, P.L.; Rohrbeck, P.; et al. Integrative Network Analysis Linking Clinical Outcomes With Environmental Exposures and Molecular Variations in Service Personnel Deployed to Balad and Bagram. J. Occup. Environ. Med. 2019, 61 (Suppl. S12), S65–S72. [Google Scholar] [CrossRef]
- Cheong, A.; Zhang, X.; Cheung, Y.Y.; Tang, W.Y.; Chen, J.; Ye, S.H.; Medvedovic, M.; Leung, Y.K.; Prins, G.S.; Ho, S.M. DNA methylome changes by estradiol benzoate and bisphenol A links early-life environmental exposures to prostate cancer risk. Epigenetics 2016, 11, 674–689. [Google Scholar] [CrossRef]
- Motta, V.; Angelici, L.; Nordio, F.; Bollati, V.; Fossati, S.; Frascati, F.; Tinaglia, V.; Bertazzi, P.A.; Battaglia, C.; Baccarelli, A.A. Integrative Analysis of miRNA and inflammatory gene expression after acute particulate matter exposure. Toxicol. Sci. 2013, 132, 307–316. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Cirillo, P.; Hu, X.; Tran, V.; Krigbaum, N.; Yu, S.; Jones, D.P.; Cohn, B. Understanding mixed environmental exposures using metabolomics via a hierarchical community network model in a cohort of California women in 1960’s. Reprod. Toxicol. 2020, 92, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Austin, C.; Smith, T.M.; Bradman, A.; Hinde, K.; Joannes-Boyau, R.; Bishop, D.; Hare, D.J.; Doble, P.; Eskenazi, B.; Arora, M. Barium distributions in teeth reveal early-life dietary transitions in primates. Nature 2013, 498, 216–219. [Google Scholar] [CrossRef] [Green Version]
- Curtin, P.; Austin, C.; Curtin, A.; Gennings, C.; Figueroa-Romero, C.; Mikhail, K.A.; Botero, T.M.; Goutman, S.A.; Feldman, E.L.; Arora, M. Dysregulated biodynamics in metabolic attractor systems precede the emergence of amyotrophic lateral sclerosis. PLoS Comput. Biol. 2020, 16, e1007773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrat, A.; Barthelemy, M.; Pastor-Satorras, R.; Vespignani, A. The architecture of complex weighted networks. Proc. Natl. Acad. Sci. USA 2004, 101, 3747–3752. [Google Scholar] [CrossRef] [Green Version]
- Brandes, U. A faster algorithm for betweenness centrality. J. Math. Sociol. 2001, 25, 163–177. [Google Scholar] [CrossRef]
- Freeman, L. Centrality in Social Networks I: Conceptual Clarification. Soc. Netw. 1979, 1, 215–239. [Google Scholar] [CrossRef] [Green Version]
- Bonacich, P. Power and centrality: A family of measures. Am. J. Sociol. 1987, 92, 1170–1182. [Google Scholar] [CrossRef]
- Harary, F. Graph Theory; Addison-Wesley: Reading, MA, USA, 1969. [Google Scholar]
- Arora, M.; Reichenberg, A.; Willfors, C.; Austin, C.; Gennings, C.; Berggren, S.; Lichtenstein, P.; Anckarsater, H.; Tammimies, K.; Bolte, S. Fetal and postnatal metal dysregulation in autism. Nat. Commun. 2017, 8, 15493. [Google Scholar] [CrossRef] [PubMed]
- de Water, E.; Curtin, P.; Gennings, C.; Chelonis, J.J.; Paule, M.; Bixby, M.; McRae, N.; Svensson, K.; Schnaas, L.; Pantic, I.; et al. Prenatal metal mixture concentrations and reward motivation in children. Neurotoxicology 2021, 88, 124–133. [Google Scholar] [CrossRef] [PubMed]
- Stafoggia, M.; Breitner, S.; Hampel, R.; Basagana, X. Statistical Approaches to Address Multi-Pollutant Mixtures and Multiple Exposures: The State of the Science. Curr. Environ. Health Rep. 2017, 4, 481–490. [Google Scholar] [CrossRef]
- Czarnota, J.; Gennings, C.; Wheeler, D.C. Assessment of weighted quantile sum regression for modeling chemical mixtures and cancer risk. Cancer Inform. 2015, 14, 159–171. [Google Scholar] [CrossRef] [Green Version]
- Carrico, C.; Gennings, C.; Wheeler, D.C.; Factor-Litvak, P. Characterization of Weighted Quantile Sum Regression for Highly Correlated Data in a Risk Analysis Setting. J. Agric. Biol. Environ. Stat. 2015, 20, 100–120. [Google Scholar] [CrossRef] [PubMed]
- Curtin, P.; Kellogg, J.; Cech, N.; Gennings, C. A random subset implementation of weighted quantile sum (WQSrs) regression for analysis of high-dimensional mixtures. Commun. Stat. Simul. Comput. 2019, 50, 1119–1134. [Google Scholar] [CrossRef]
- Li, H.; Deng, W.; Small, R.; Schwartz, J.; Liu, J.; Shi, L. Health effects of air pollutant mixtures on overall mortality among the elderly population using Bayesian kernel machine regression (BKMR). Chemosphere 2022, 286, 131566. [Google Scholar] [CrossRef]
- Bobb, J.F.; Claus Henn, B.; Valeri, L.; Coull, B.A. Statistical software for analyzing the health effects of multiple concurrent exposures via Bayesian kernel machine regression. Environ. Health 2018, 17, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffman, D.J.; Reynolds, R.M.; Hardy, D.B. Developmental origins of health and disease: Current knowledge and potential mechanisms. Nutr. Rev. 2017, 75, 951–970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorban, A.N.; Tyukina, T.A.; Pokidysheva, L.I.; Smirnova, E.V. Dynamic and thermodynamic models of adaptation. Phys. Life Rev. 2021, 37, 17–64. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Measure | p-Value |
|---|---|
| Degree | 0.002 |
| Closeness | 0.301 |
| Betweenness | 0.895 |
| Eigenvalue | 0.372 |
| Clustering Coefficient | 0.375 |
| Eccentricity | 0.330 |
| Element | Degree | Closeness | Betweenness | Eigenvalue | Clustering Coefficient | Eccentricity |
|---|---|---|---|---|---|---|
| Ba | 0.002 | 0.301 | 0.895 | 0.372 | 0.375 | 0.330 |
| Cu | 0.001 | 0.325 | 0.882 | 0.007 | <0.001 | 0.582 |
| Li | 0.002 | 0.334 | 0.432 | 0.006 | <0.001 | 0.424 |
| Mg | 0.003 | 0.273 | 0.987 | 0.314 | 0.234 | 0.532 |
| Mn | 0.003 | 0.279 | 0.974 | 0.632 | 0.438 | 0.120 |
| Sr | 0.015 | 0.271 | 0.782 | 0.510 | 0.153 | 0.872 |
| Zn | 0.027 | 0.261 | 0.741 | 0.539 | 0.041 | 0.555 |
| Measure | p-Value |
|---|---|
| Degree | 0.784 |
| Closeness | 0.000 |
| Betweenness | 0.052 |
| Eigenvalue | 0.057 |
| Clustering Coefficient | 0.034 |
| Eccentricity | 0.007 |
| Element | Degree | Closeness | Betweenness | Eigenvalue | Clustering Coefficient | Eccentricity |
|---|---|---|---|---|---|---|
| Ba | 0.784 | <0.001 | 0.052 | 0.057 | 0.034 | 0.007 |
| Cu | 0.182 | <0.001 | 0.731 | 0.002 | 0.006 | 0.640 |
| Li | <0.001 | <0.001 | 0.434 | <0.001 | <0.001 | 0.137 |
| Mg | <0.001 | 0.341 | 0.399 | <0.001 | 0.044 | 0.001 |
| Mn | <0.001 | <0.001 | <0.001 | 0.341 | 0.321 | 0.107 |
| Sr | 0.734 | <0.001 | 0.906 | 0.002 | 0.118 | 0.503 |
| Zn | <0.001 | 0.022 | 0.066 | <0.001 | <0.001 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Curtin, A.; Austin, C.; Giuliani, A.; Ruiz Marín, M.; Merced-Nieves, F.; Téllez-Rojo, M.M.; Wright, R.O.; Arora, M.; Curtin, P. Network Dynamics in Elemental Assimilation and Metabolism. Entropy 2021, 23, 1633. https://doi.org/10.3390/e23121633
Curtin A, Austin C, Giuliani A, Ruiz Marín M, Merced-Nieves F, Téllez-Rojo MM, Wright RO, Arora M, Curtin P. Network Dynamics in Elemental Assimilation and Metabolism. Entropy. 2021; 23(12):1633. https://doi.org/10.3390/e23121633
Chicago/Turabian StyleCurtin, Austen, Christine Austin, Alessandro Giuliani, Manuel Ruiz Marín, Francheska Merced-Nieves, Martha M. Téllez-Rojo, Robert O. Wright, Manish Arora, and Paul Curtin. 2021. "Network Dynamics in Elemental Assimilation and Metabolism" Entropy 23, no. 12: 1633. https://doi.org/10.3390/e23121633
APA StyleCurtin, A., Austin, C., Giuliani, A., Ruiz Marín, M., Merced-Nieves, F., Téllez-Rojo, M. M., Wright, R. O., Arora, M., & Curtin, P. (2021). Network Dynamics in Elemental Assimilation and Metabolism. Entropy, 23(12), 1633. https://doi.org/10.3390/e23121633