In this section, we will summarize the results of the comparison of adaptive mechanisms tested in the three presented tasks, focusing on the performance differences induced by robots controlled by Boolean networks taken from ordered, critical and chaotic ensembles.
Comparison of the Adaptive Mechanisms
As stated before, we run 50 replicas for each configuration of parameters and collected the following statistics:
- Mean of max objective function per epoch
To represent the robots’ ability to adapt as the length of the adaptation phase increases, we take for each epoch the maximum score (returned value by the objective function) achieved so far by the robot, taking into account all previous epochs. Subsequently, we calculate the average so as to obtain for each epoch, 500 in total, a point that represents the average of the maximum score obtained so far by all robots. The trend will always be increasing.
- Area under the curve
To provide quantitative information about the differential performances of all the combinations of adaptive mechanisms and dynamical regimes, we calculate and rank from highest to lowest the area under the curve of the trend traced by the mean of the maximum objective function along the epochs.
- Box-plot of max objective function scores
In this type of statistic, each box-plot is populated by the maximum objective function score of each robot. The maximum score is the highest score that a robot was able to reach in one of the 500 epochs available.
- Mean rank of max scores
For this analysis, we sort all the robot’s max scores from highest to lowest and assign a rank. Afterwards, we compute the mean of the rank of every experimental case by grouping them by the mechanism and dynamical regime. For this statistic, the lower, the better.
In
Figure 4, we present, for each task, the trend of the mean score per epoch, pointing out the relationship between the type of Boolean network and the adaptive mechanism.
First of all, we observe that Boolean networks generated with (blue ones) obtain, except for Task II, a lower average score regardless of the adaptive technique used. Secondly, we can also notice that the critical dynamical regime (green curves), in general, tends to dominate the others, with a slightly better performance of in–out mapping with respect to other adaptation schemas. To better characterize the contributions made, respectively, by the adaptive mechanism and the dynamical regime of the Boolean network in the critical controller case, we have to note the relative difference in performance obtained by the mutation mechanism compared to the other two. Indeed, the latter produces lower values than the other two mechanisms at each epoch, leading us to conclude that the results of the hybrid mechanism in the critical case were more positively affected by the change in input and output mapping than by the component concerning the Boolean network modifications. In contrast, the configuration of (initially) ordered controller (red curves) with hybrid technique appears to take advantage of the mutational component of the hybrid mechanism, except in Task I, which is the simplest one. However, how much these variations affect the dynamical regime of these networks is unclear, and in the following, we will analyze this aspect by analyzing their Derrida values.
As previously stated, to quantitatively evaluate the differences in performance, we also calculated the area under each curve shown in
Figure 4. We then ranked them and reported the 5 best for each task in
Table 2. Interestingly, the first 4 positions are occupied by critical RBNs, for all tasks, and on only one occasion does the mechanism of mutation appear. Furthermore, we note that in
of the tasks, the
in–out mapping turns out to be the best for this type of statistic.
The overall picture seems to point toward the initial postulated hypothesis that the critical RBNs already have all the computational characteristics to perform the tasks, and therefore, the in–out mapping technique is sufficient to obtain good results.
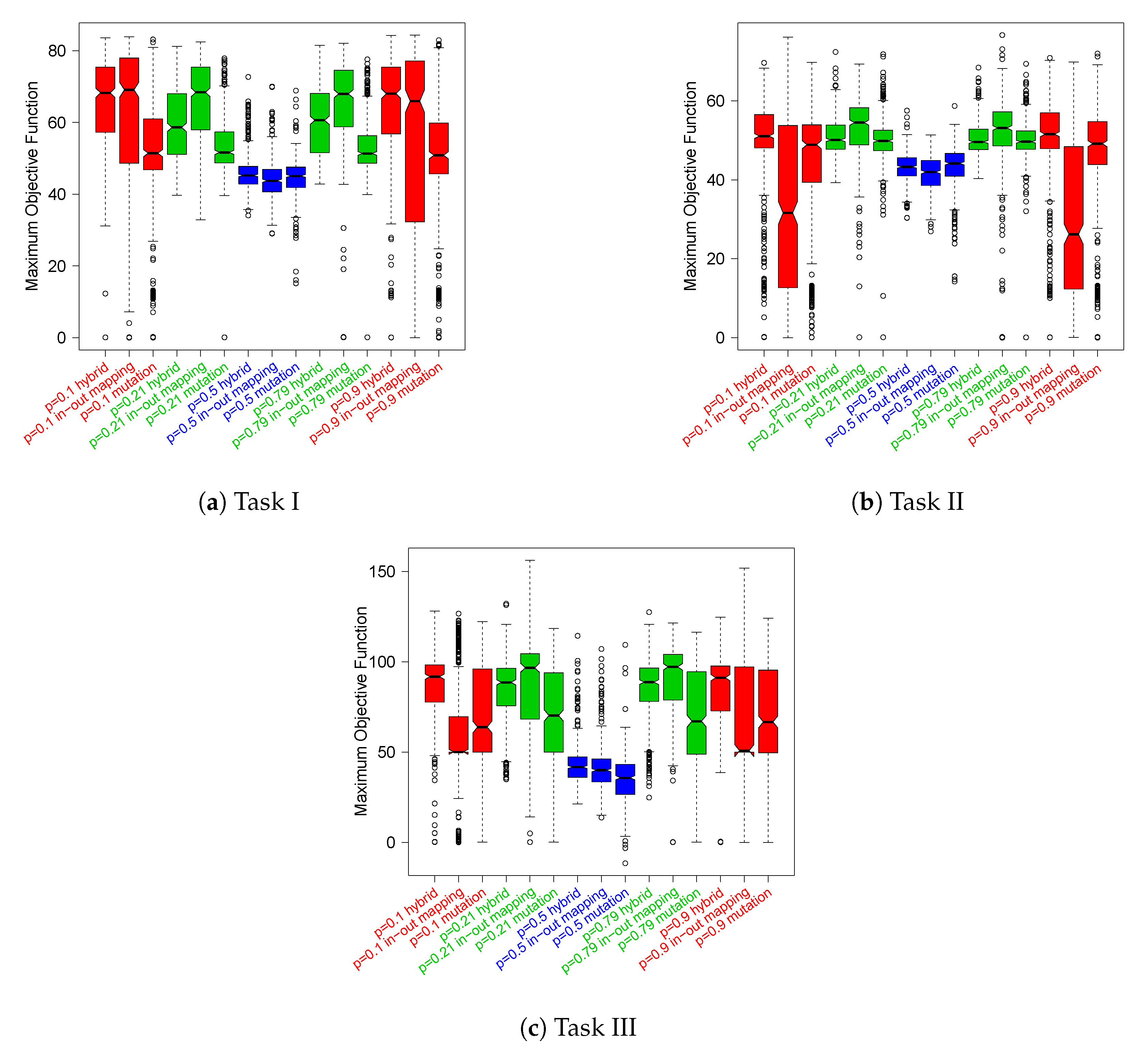
In
Figure 5, we report the distributions of the maximum objective function scores obtained by the robots along their epochs of adaptation, which are grouped by task. These data also confirm that Boolean networks generated with
perform poorly on average. Indeed, the best robots in this parameter setup fail to achieve high values of the objective function across all the tasks. In addition, we can state that at least for this kind of analysis, there is no clear winner between the critical and ordered networks, which are the best configurations for the previous analysis. However, the box-plots seem to suggest that as the task difficulty increases, i.e., from
I to
III, the relationships between the two distributions gradually change in favor of the critical configuration. As regards the adaptation mechanism, both cases of critical RBNs (namely,
and
) with
in–out mapping present median values comparable to (in Task I) or higher to (Task II and III) other adaptation mechanisms.
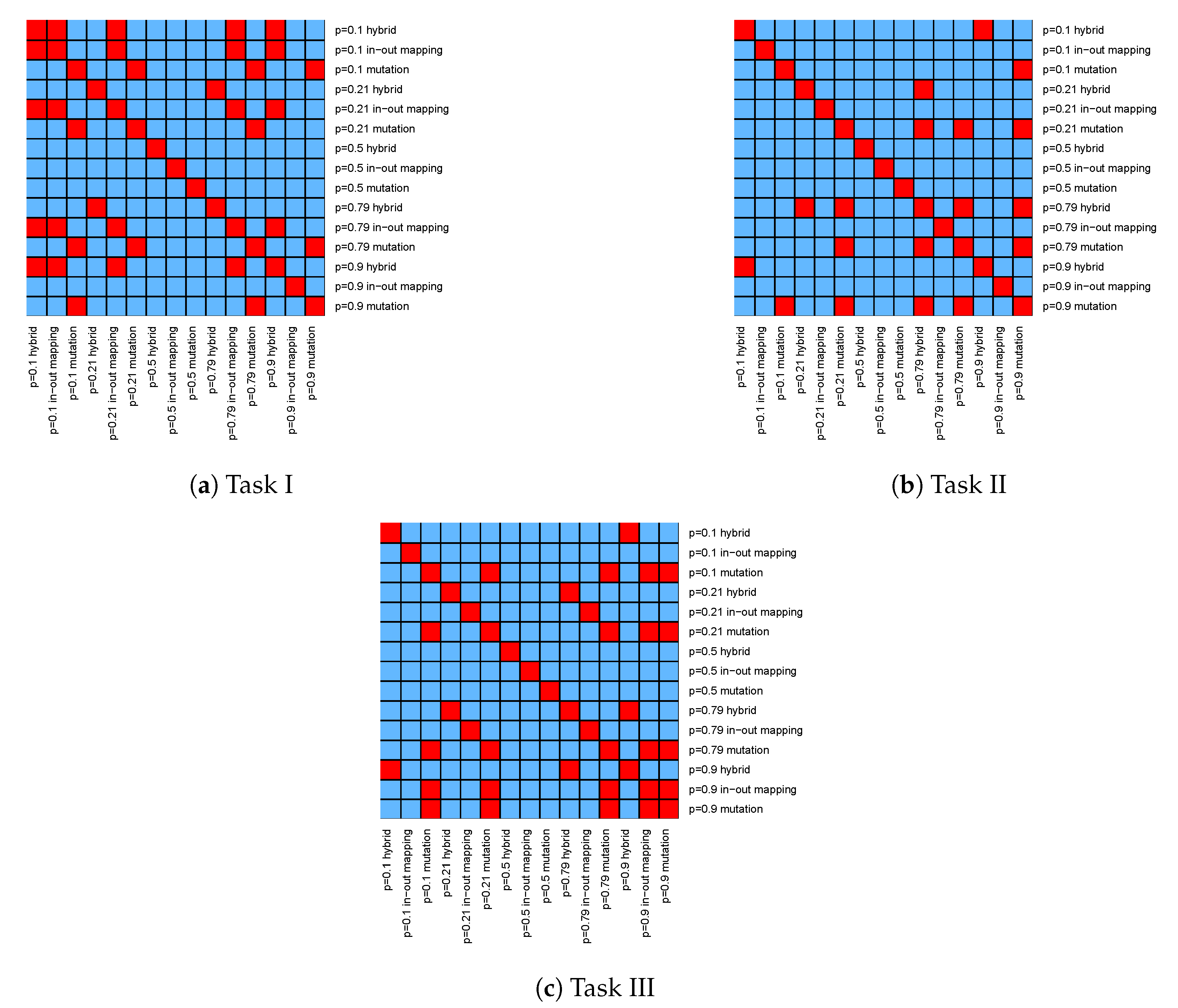
The unpaired Wilcoxon statistical hypothesis tests, reported in
Appendix C in
Figure A2, confirm that the medians of the distributions of
and
in–out mapping are significantly different from the other distributions’ medians in Tasks II and III, with some exceptions in Task I.
As briefly introduced above, another prominent analysis is that concerning the
nominal dynamical regime, i.e., the one statistically possessed by the networks generated randomly at the beginning of the adaptation phase, against the
actual one, i.e., the operating dynamical regime expressed by the BNs of the best robots. The
actual dynamical regime is the result of the particular adaptive pressure exerted by the objective function on the environment–robot system. Indeed, having undergone modifications to the internal structure through the mutation and hybrid mechanisms, Boolean networks belonging to the
configuration that have performance comparable to the critical ones, initially ordered, may have undergone a change to their dynamical regime of operation, which, arguably, could have led them to resemble, dynamically, those in the critical regime. In general, structural modifications to the Boolean networks introduced by the
mutation and
hybrid adaptive techniques no longer allow us to consider the networks as part of the random Boolean network ensemble from which they were initially sampled, since the bias selected by the adaptation techniques could have undergone substantial changes. So, a precise characterization of the dynamical regime of the networks that have been subjected to adaptation is needed. In this regard, we study their Derrida parameter [
34]
at one step.
The procedure for computing the Derrida value for a single state of a Boolean network is as follows: a copy of the state of the Boolean network is created, and the value of a randomly chosen variable is negated; then, a synchronous update is performed for both states, and the Hamming distance between the two resulting states is measured. Usually, this procedure is repeated several times, for different variables, to determine an average value. The result measures the average propagation level of a perturbation and is statistically greater than 1 in chaotic networks, and it is less than 1 in ordered and 1 in critical networks. Since in our experiments, Boolean networks are embedded in robot bodies, and thus regions of their state spaces are not equally accessible due to physical constraints and the control program they are actually implementing, we compute, for each robot, the Derrida value using the following procedure:
We identify the epoch in which the robot obtained the best score;
We extrapolate the state of the network for all the 800 steps composing the epoch;
We calculate the Derrida value for each of the 800 states and on all the possible perturbations, . Then, an average value of all states belonging to the robot is computed;
Finally, the average value is used to generate a point on the scatter plot.
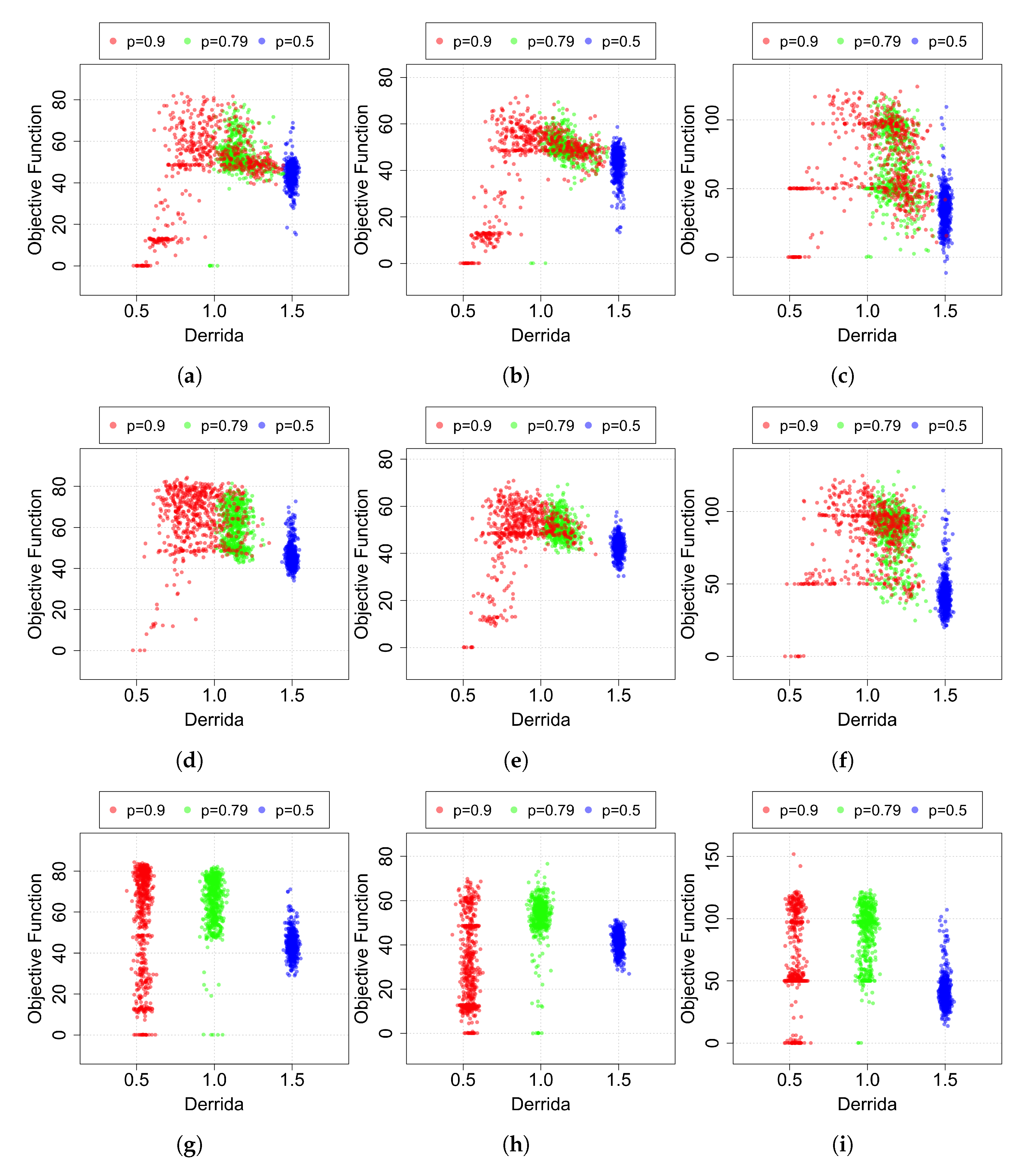
The value thus calculated represents the average propagation level of a perturbation only on the states in which the robot has transited during its best epoch. Then, these values are reported in
Figure 6 with respect to their relative scores. For reasons of clarity,
Figure 6 reports the comparison of initially ordered, critical and chaotic networks generated using the following bias values:
. The other bias values taken into account, namely
, which show a similar trend, are reported in
Appendix B and in particular in
Figure A1.
In the cases where only an
in–out mapping adaptive mechanism is applied, it is possible to notice well-divided clusters in relation to the Derrida value and to the parameter
p used to generate the RBNs. When the mutation is introduced (
hybrid and
mutation techniques), the regions start to overlap. In particular, it is possible to notice that the green and red regions (
and
, and in the same way for
and
in
Figure A1) move to the right, obtaining a higher average Derrida value. Remarkably, the effect is particularly marked for the initially ordered networks. To better evaluate the effect of the “dynamical regime shift” observed in
Figure A1) and induced by the mechanisms that change the BNs structure, we perform a further analysis: we compute the mean of the ranking of the maximum scores in both
nominal and
actual dynamical regimes. To do so, for the
nominal dynamical regime, we considered as ordered the BNs generated with
, critical those with
and chaotic with
. Meanwhile, for the
actual dynamical regime, we considered them ordered if
, critical if
and
, and, lastly, chaotic if
, with
. The results are reported in
Table 3 for Task III, while those concerning the dynamical regime shift observed in Tasks I and II are presented in
Appendix D, in
Table A6 and
Table A7. In general, we can note how all the configurations with (initially) ordered networks that are present in the first positions of the “nominal” column (remember that in this analysis, the lower, the better) are superseded by the critical ones when we consider the actual operating dynamical regimes expressed by the BNs that control the robots.
Overall, this result demonstrates how initially ordered networks that undergo adaptation by the hybrid and mutation mechanisms have statistically similar dynamical characteristics to the critical ones and strengthen our initial hypothesis.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}








