


Noise Enhancement of Neural Information Processing

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
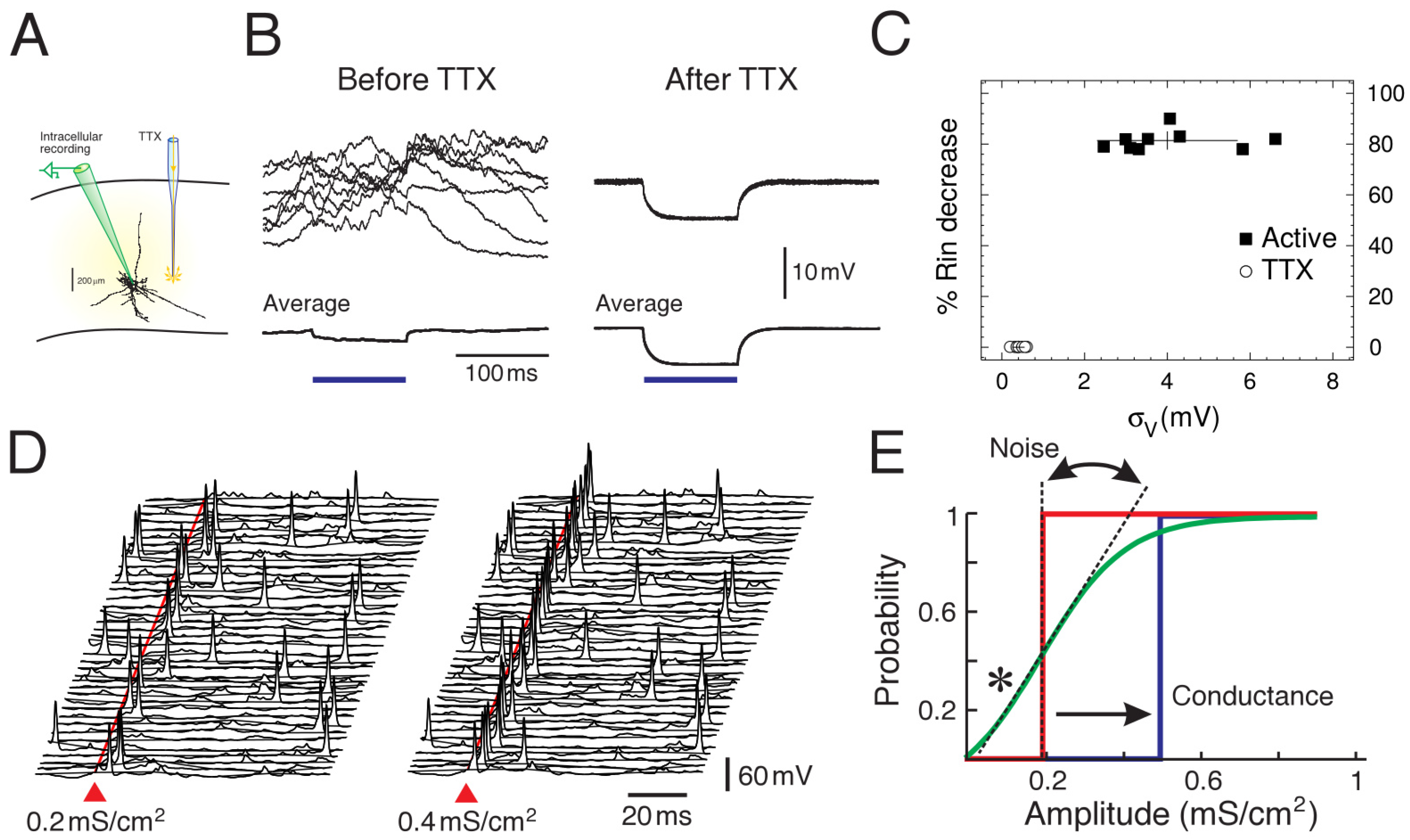
2. Noisy Neurons
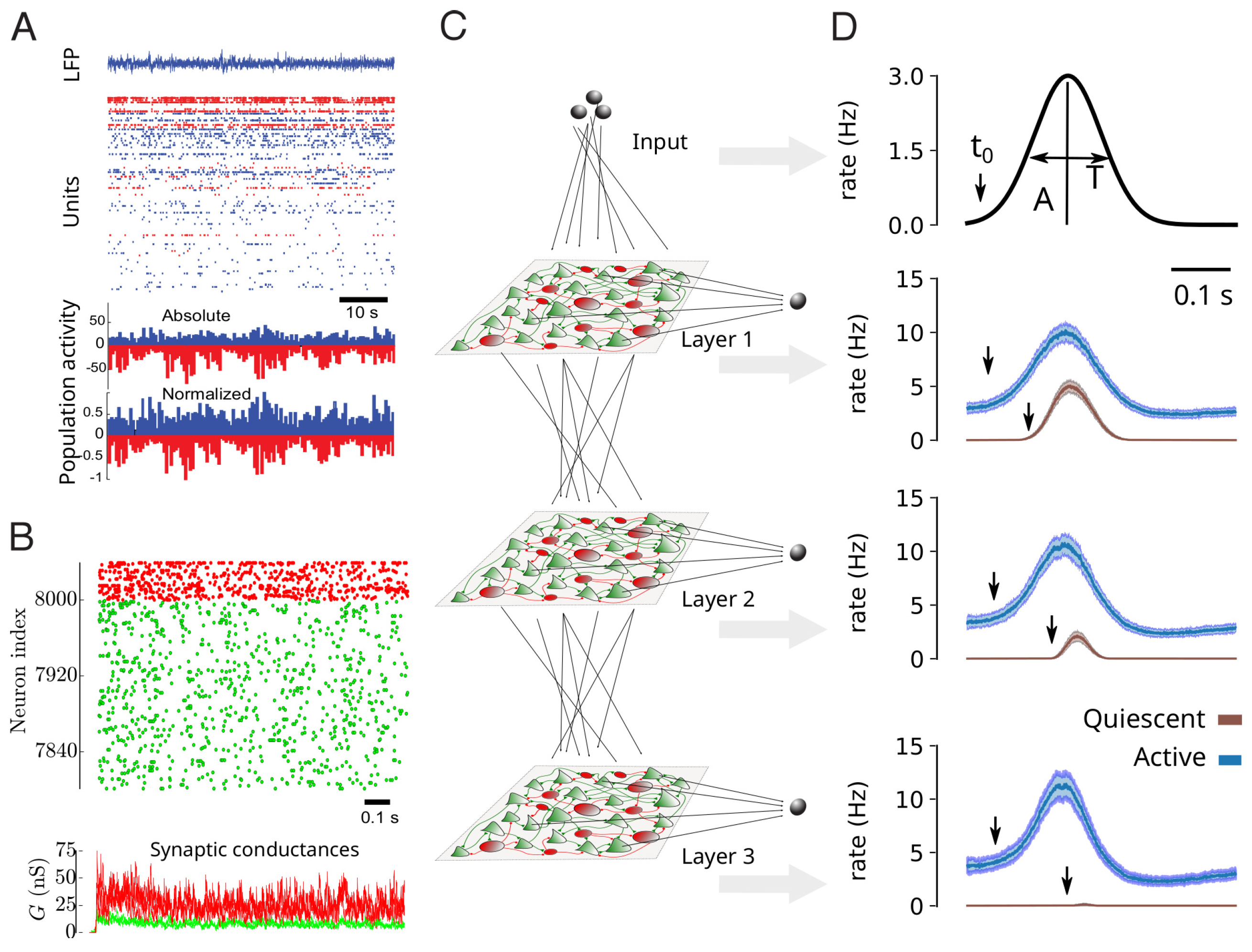
3. Noisy Networks
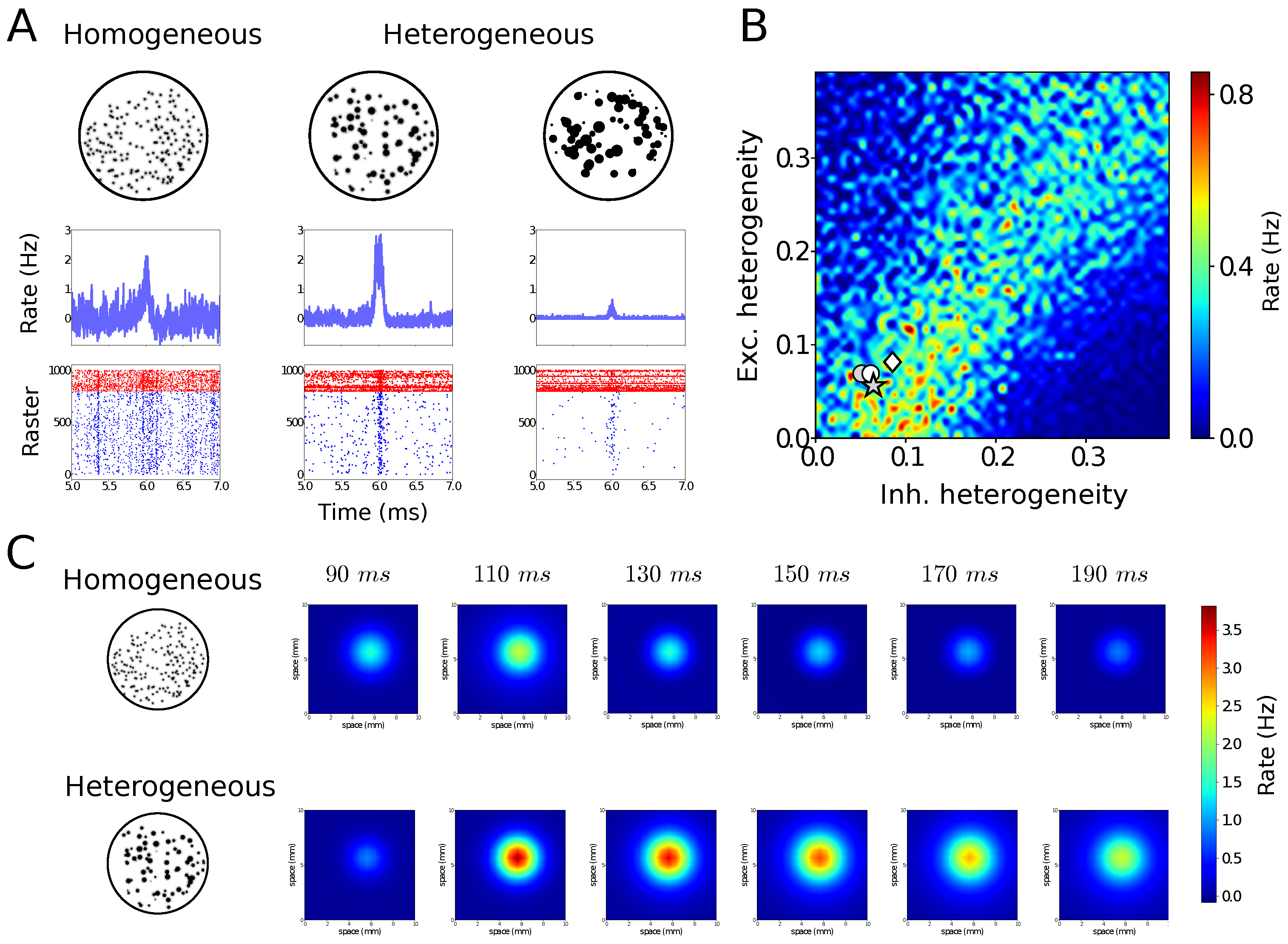
4. Structural Noise
5. Discussion
Funding
Acknowledgments
Conflicts of Interest
References
- Baranyi, A.; Szente, M.B.; Woody, C.D. Electrophysiological characterization of different types of neurons recorded in vivo in the motor cortex of the cat. II. Membrane parameters, action potentials, current-induced voltage responses and electrotonic structures. J. Neurophysiol. 1993, 69, 1865–1879. [Google Scholar] [CrossRef]
- Matsumura, M.; Cope, T.; Fetz, E.E. Sustained excitatory synaptic input to motor cortex neurons in awake animals revealed by intracellular recording of membrane potentials. Exp. Brain Res. 1988, 70, 463–469. [Google Scholar] [CrossRef]
- Steriade, M.; Timofeev, I.; Grenier, F. Natural waking and sleep states: A view from inside neocortical neurons. J. Neurophysiol. 2001, 85, 1969–1985. [Google Scholar] [CrossRef] [Green Version]
- Barrett, J.N. Motoneuron dendrites: role in synaptic integration. Fed. Proc. 1975, 34, 1398–1407. [Google Scholar]
- Bernander, O.; Douglas, R.J.; Martin, K.A.; Koch, C. Synaptic background activity influences spatiotemporal integration in single pyramidal cells. Proc. Natl. Acad. Sci. USA 1991, 88, 11569–11573. [Google Scholar] [CrossRef]
- Destexhe, A.; Paré, D. Impact of network activity on the integrative properties of neocortical pyramidal neurons in vivo. J. Neurophysiol. 1999, 81, 1531–1547. [Google Scholar] [CrossRef] [Green Version]
- Holmes, W.R.; Woody, C.D. Effects of uniform and non-uniform synaptic ’activation-distributions’ on the cable properties of modeled cortical pyramidal neurons. Brain Res. 1989, 505, 12–22. [Google Scholar] [CrossRef]
- Rudolph, M.; Destexhe, A. Tuning neocortical pyramidal neurons between integrators and coincidence detectors. J. Comput. Neurosci. 2003, 14, 239–251. [Google Scholar] [CrossRef]
- Destexhe, A.; Rudolph, M.; Paré, D. The high-conductance state of neocortical neurons in vivo. Nat. Rev. Neurosci. 2003, 4, 739–751. [Google Scholar] [CrossRef]
- Paré, D.; Shink, E.; Gaudreau, H.; Destexhe, A.; Lang, E.J. Impact of spontaneous synaptic activity on the resting properties of cat neocortical pyramidal neurons in vivo. J. Neurophysiol. 1998, 79, 1450–1460. [Google Scholar] [CrossRef]
- Buzsáki, G. Large-scale recording of neuronal ensembles. Nat. Neurosci. 2004, 7, 446–451. [Google Scholar] [CrossRef]
- Dehghani, N.; Peyrache, A.; Telenczuk, B.; Le Van Quyen, M.; Halgren, E.; Cash, S.S.; Hatsopoulos, N.G.; Destexhe, A. Dynamic balance of excitation and inhibition in human and monkey neocortex. Sci. Rep. 2016, 6, 23176. [Google Scholar] [CrossRef] [Green Version]
- Lin, L.; Chen, G.; Xie, K.; Zaia, K.A.; Zhang, S.; Tsien, J.Z. Large-scale neural ensemble recording in the brains of freely behaving mice. J. Neurosci. Methods 2006, 155, 28–38. [Google Scholar] [CrossRef]
- Hô, N.; Destexhe, A. Synaptic background activity enhances the responsiveness of neocortical pyramidal neurons. J. Neurophysiol. 2000, 84, 1488–1496. [Google Scholar] [CrossRef] [Green Version]
- Zohary, E.; Shadlen, M.N.; Newsome, W.T. Correlated neuronal discharge rate and its implications for psychophysical performance. Nature 1994, 370, 140–143. [Google Scholar] [CrossRef]
- Rudolph, M.; Pospischil, M.; Timofeev, I.; Destexhe, A. Inhibition determines membrane potential dynamics and controls action potential generation in awake and sleeping cat cortex. J. Neurosci. 2007, 27, 5280–5290. [Google Scholar] [CrossRef] [Green Version]
- Reig, R.; Zerlaut, Y.; Vergara, R.; Destexhe, A.; Sanchez-Vives, M.V. Gain modulation of synaptic inputs by network state in auditory cortex in vivo. J. Neurosci. 2015, 35, 2689–2702. [Google Scholar] [CrossRef] [Green Version]
- Waters, J.; Helmchen, F. Background synaptic activity is sparse in neocortex. J. Neurosci. 2006, 26, 8267–8277. [Google Scholar] [CrossRef] [Green Version]
- Chance, F.S.; Abbott, L.F.; Reyes, A.D. Gain modulation from background synaptic input. Neuron 2002, 35, 773–782. [Google Scholar] [CrossRef] [Green Version]
- Shu, Y.; Hasenstaub, A.; Badoual, M.; Bal, T.; McCormick, D.A. Barrages of synaptic activity control the gain and sensitivity of cortical neurons. J. Neurosci. 2003, 23, 10388–10401. [Google Scholar] [CrossRef] [Green Version]
- Wolfart, J.; Debay, D.; Le Masson, G.; Destexhe, A.; Bal, T. Synaptic background activity controls spike transfer from thalamus to cortex. Nat. Neurosci. 2005, 8, 1760–1767. [Google Scholar] [CrossRef]
- Kuhn, A.; Aertsen, A.; Rotter, S. Neuronal integration of synaptic input in the fluctuation-driven regime. J. Neurosci. 2004, 24, 2345–2356. [Google Scholar] [CrossRef] [Green Version]
- Zerlaut, Y.; Teleńczuk, B.; Deleuze, C.; Bal, T.; Ouanounou, G.; Destexhe, A. Heterogeneous firing rate response of mouse layer v pyramidal neurons in the fluctuation-driven regime. J. Physiol. 2016, 594, 3791–3808. [Google Scholar] [CrossRef] [Green Version]
- Zerlaut, Y.; Chemla, S.; Chavane, F.; Destexhe, A. Modeling mesoscopic cortical dynamics using a mean-field model of conductance-based networks of adaptive exponential integrate-and-fire neurons. J. Comput. Neurosci. 2018, 44, 45–61. [Google Scholar] [CrossRef]
- Zerlaut, Y.; Destexhe, A. Enhanced responsiveness and low-level awareness in stochastic network states. Neuron 2017, 94, 1002–1009. [Google Scholar] [CrossRef] [Green Version]
- Brunel, N. Dynamics of sparsely connected networks of excitatory and inhibitory spiking neurons. J. Comput. Neurosci. 2000, 8, 183–208. [Google Scholar] [CrossRef]
- van Vreeswijk, C.; Sompolinsky, H. Chaos in neuronal networks with balanced excitatory and inhibitory activity. Science 1996, 274, 1724–1726. [Google Scholar] [CrossRef] [Green Version]
- Balog, I.; Uzelac, K. Quenched disorder: demixing thermal and disorder fluctuations. Phys. Rev. E 2012, 86, 061124. [Google Scholar] [CrossRef]
- Radzihovsky, L. Introduction to Quenched Disorder; University of Colorado: Boulder, CO, USA, 2015. [Google Scholar]
- Braitenberg, V.; Schüz, A. Cortex: Statistics and Geometry of Neuronal Connectivity; Springer: Berlin/Heidelberg, Germany, 1998. [Google Scholar]
- Di Volo, M.; Destexhe, A. Optimal responsiveness and information flow in networks of heterogeneous neurons. Sci. Rep. 2021, 11, 17611. [Google Scholar] [CrossRef]
- Mejias, J.; Longtin, A. Optimal heterogeneity for coding in spiking neural networks. Phys. Rev. Lett. 2012, 108, 228102. [Google Scholar] [CrossRef] [Green Version]
- Wiesenfeld, K.; Moss, F. Stochastic resonance and the benefits of noise: from ice ages to crayfish and SQUIDs. Nature 1995, 373, 33–36. [Google Scholar] [CrossRef]
- Casali, A.G.; Gosseries, O.; Rosanova, M.; Boly, M.; Sarasso, S.; Casali, K.R.; Casarotto, S.; Bruno, M.A.; Laureys, S.; Tononi, G.; et al. A theoretically based index of consciousness independent of sensory processing and behavior. Sci. Transl. Med. 2013, 5, 198ra105. [Google Scholar] [CrossRef]
- Massimini, M.; Ferrarelli, F.; Huber, R.; Esser, S.K.; Singh, H.; Tononi, G. Breakdown of cortical effective connectivity during sleep. Science 2005, 309, 2228–2232. [Google Scholar] [CrossRef]
- Sarasso, S.; Boly, M.; Napolitani, M.; Gosseries, O.; Charland-Verville, V.; Casarotto, S.; Rosanova, M.; Casali, A.G.; Brichant, J.F.; Boveroux, P.; et al. Consciousness and Complexity during Unresponsiveness Induced by Propofol, Xenon, and Ketamine. Curr. Biol. 2015, 25, 3099–3105. [Google Scholar] [CrossRef] [Green Version]
- Goldman, J.S.; Kusch, L.; Yalcinkaya, B.H.; Depannemaecker, D.; Nghiem, T.A.E.; Jirsa, V.; Destexhe, A. Brain-scale emergence of slow-wave synchrony and highly responsive asynchronous states based on biologically realistic population models simulated in the virtual brain. bioRxiv 2020. [Google Scholar] [CrossRef]
- Goldman, J.S.; Kusch, L.; Yalcinkaya, B.H.; Nghiem, T.A.E.; Jirsa, V.; Destexhe, A. A comprehensive neural simulation of slow-wave 1 sleep and highly responsive wakefulness dynamics 2. bioRxiv 2021. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Destexhe, A. Noise Enhancement of Neural Information Processing. Entropy 2022, 24, 1837. https://doi.org/10.3390/e24121837
Destexhe A. Noise Enhancement of Neural Information Processing. Entropy. 2022; 24(12):1837. https://doi.org/10.3390/e24121837
Chicago/Turabian StyleDestexhe, Alain. 2022. "Noise Enhancement of Neural Information Processing" Entropy 24, no. 12: 1837. https://doi.org/10.3390/e24121837
APA StyleDestexhe, A. (2022). Noise Enhancement of Neural Information Processing. Entropy, 24(12), 1837. https://doi.org/10.3390/e24121837