1. Introduction
How might aging manifest in the dynamics of biological systems? One hypothesis that has received much attention in recent years is the
loss of complexity hypothesis, which claims that the loss of function associated with aging is the result of a loss of complexity in the underlying physiological systems [
1]. As aging is associated with an increase in the incidence and severity of falls, the complexity of the postural system is of particular relevance [
2]. Moreover, as individuals rarely perform postural tasks in the absence of an additional task (e.g., conversing with others or manipulating nearby objects), this postural complexity should be measured under ecologically valid, multi-task conditions.
Traditionally, however, research on postural behavior has not had as its focus the complexity of the underlying physiological systems. Rather, research has focused primarily on quantifying the
stability of the postural system, often during laboratory-specific paradigms. In assessing stability, linear assessments of the center of pressure (e.g., ellipse area) and relations of the center of mass to the base of support (e.g., proximity) have been widely studied [
3]. While it is true that these measures may provide valuable information in certain contexts, inherent assumptions of linearity restrict the breadth and depth of information that might be gleaned. When measuring the ellipse area of the center of pressure, for example, the traditionally dominant perspective is that an increase in variability is alone indicative of a pathological system. Such a perspective ignores the nonlinear nature of the postural system and conflates variability with pathology [
4].
A more modern approach is to study the
response of the postural system to mechanical perturbation [
5,
6]. This approach is particularly attractive to those who wish to study the stability of the postural system, as stability is often defined as the resistance of a system to perturbation. This approach, however, is not without its own set of limitations. For example, as an external perturbation is a discrete event, the data obtained during such an experiment are limited, naturally, to a discrete response. Moreover, there is difficulty in scaling the degree of perturbation to be applied, as well as in mitigating the risks associated with mechanical perturbation. An alternative approach is to examine the behavior of the postural system while individuals perform perceptual tasks (i.e., tasks in which the primary objective is to perceive ecological information) during mechanically unperturbed stance. This approach, while not unique to the present study, may permit the examination of more natural postural behavior and provide clinically relevant information.
Importantly, the examination of postural behavior must extend beyond traditional assessments aimed at quantifying the stability of the postural system. An assessment of
complexity may provide alternative insight into the dynamics of the postural system across a wide array of temporal scales and under a wide variety of conditions. Note that this distinction between stability and complexity is neither arbitrary nor is it meant to suggest that assessments of stability are unimportant. Rather, it is meant to offer a delineation between two distinct features of biological systems and to suggest that neither feature should be overlooked. As complexity, much like stability, is more frequently intuited than precisely defined, it is unsurprising that there is no widely accepted definition of complexity. Roughly, however, the complexity of a system may be thought of as the degree to which the system exhibits meaningful connectivity across spatial and temporal scales [
1].
Various techniques have been proposed to measure the complexity of experimentally obtained signals. One such technique,
Sample Entropy, provides an estimate of signal complexity by computing the probability that two sufficiently similar segments of equal length taken from a signal will
cease to remain sufficiently similar when the length of each segment is increased by one sample [
7]. It follows that the Sample Entropy of a purely periodic signal will be equal to zero, while the Sample Entropy of a complex, biophysical signal will be greater than zero. If, however, one computes the Sample Entropy of a signal generated purely by stochastic processes (e.g., white noise), the Sample Entropy of that signal will approach some maximal value despite a distinct
lack of complexity. This inability of Sample Entropy to distinguish, reliably, between signals with complex structure and those without renders this approach insufficient when attempting to measure biophysical signal complexity.
Accordingly, Costa and colleagues developed the Multiscale Sample Entropy (MSE) analysis, which, unlike Sample Entropy, considers the
fractal structure inherent in biological systems [
8,
9,
10]. In this approach, a signal is first coarse-grained into a given number of signals pertaining to a given number of timescales. This coarse-graining procedure consists of partitioning the original signal into non-overlapping windows of a given number of samples, averaging the samples within each window, and concatenating each window as to reconstruct the coarse-grained time series. This procedure will yield one coarse-grained signal, or
scale, per window length, with the original signal occupying scale one. From here, Sample Entropy can be computed across timescales, thereby providing a more robust measure of signal complexity.
Assessing such a measure of postural complexity while individuals engage with a listening task may provide unique insight into how the postural system is able to reorganize its degrees of freedom in response to increased perceptual demands. Existing literature on postural behavior during listening suggests that an increase in listening effort is associated with an increase in postural sway area, particularly for middle-aged adults [
11]. While this increase in postural sway area is generally interpreted as a reduction in postural control resulting from the availability of fewer resources, it is not presently clear how aging impacts the underlying dynamics of the postural system during listening.
Here, we apply the MSE analysis to postural data obtained while young and middle-aged adults engaged with a listening task of varying degrees of difficulty. By assessing the complexity of postural behavior during such a task, insight into how the postural system is able to adapt to increased perceptual demands might be gained. Moreover, assessing the complexity of postural behavior across physiologically relevant timescales (e.g., those associated with physiological tremor) might serve to further elucidate age-related changes in complexity. It is expected that the complexity exhibited by the young adults will be greater than the complexity exhibited by the middle-aged adults, and that this difference will increase as a function of task difficulty.
2. Materials and Methods
2.1. Participants
As part of a larger study, 16 young adults (18 to 28 years of age; mean 22 years) and 16 middle-aged adults (48 to 64 years of age; mean 58 years) were recruited, with both groups being comprised of 11 females and 5 males. Note that data from one young participant and two middle-aged participants were corrupted during collection, resulting in groups of 15 and 14 being used for the present study. The young participants all had clinically normal hearing (pure-tone thresholds <20 dB HL from 250 to 8000 Hz), and all but two of the middle-aged participants had clinically normal hearing (pure-tone thresholds <30 dB HL) from 250 to 4000 Hz. Participants were recruited from the general population and were screened for visual, vestibular, and motor impairments prior to collection. For a full description of participant characteristics, see Helfer et al. [
11]. All procedures were approved by the University of Massachusetts Amherst Institutional Review Board (IRB).
2.2. Procedures
Participants stood with their feet shoulder-width apart on a 40 × 60 cm piezoelectric force platform (Kistler Instruments Corporation, Amherst, NY, USA) while listening to and repeating pre-recorded sentences. These sentences were designed to have low predictability while being grammatically feasible (e.g., Theo found the pink menu and the true item here). During the listening task, a loudspeaker was placed 1.2 m in front of the participants and was adjusted to the ear height of each participant. The loudspeaker simultaneously played both a target sentence (i.e., a sentence which the participants were instructed to repeat) as well as two masking sentences (i.e., sentences similar to the target sentence which the participants were instructed to ignore). These masking sentences were played from random starting points as to minimize any grammatical alignment of the masking and target sentences.
For the present study, three conditions were analyzed. The first condition consisted of participants standing on the force platform without any listening task. This condition, referred to as the baseline condition, served to provide the experimenters with the baseline postural dynamics of each participant. The remaining two conditions consisted of participants standing on the force platform while performing the listening task described above. The difficulty of the listening task was modified for these two conditions by adjusting the signal-to-noise ratio (SNR) of the stimuli. In the first of these remaining conditions, the 0 dB condition, the combined energy of the masking sentences was equal to that of the target sentence. In the final condition, the −6 dB condition, the combined energy of the masking sentences was 6 dB greater than that of the target sentence. The order of conditions was randomized for each participant, with each condition lasting approximately 80 s.
2.3. Data Analysis
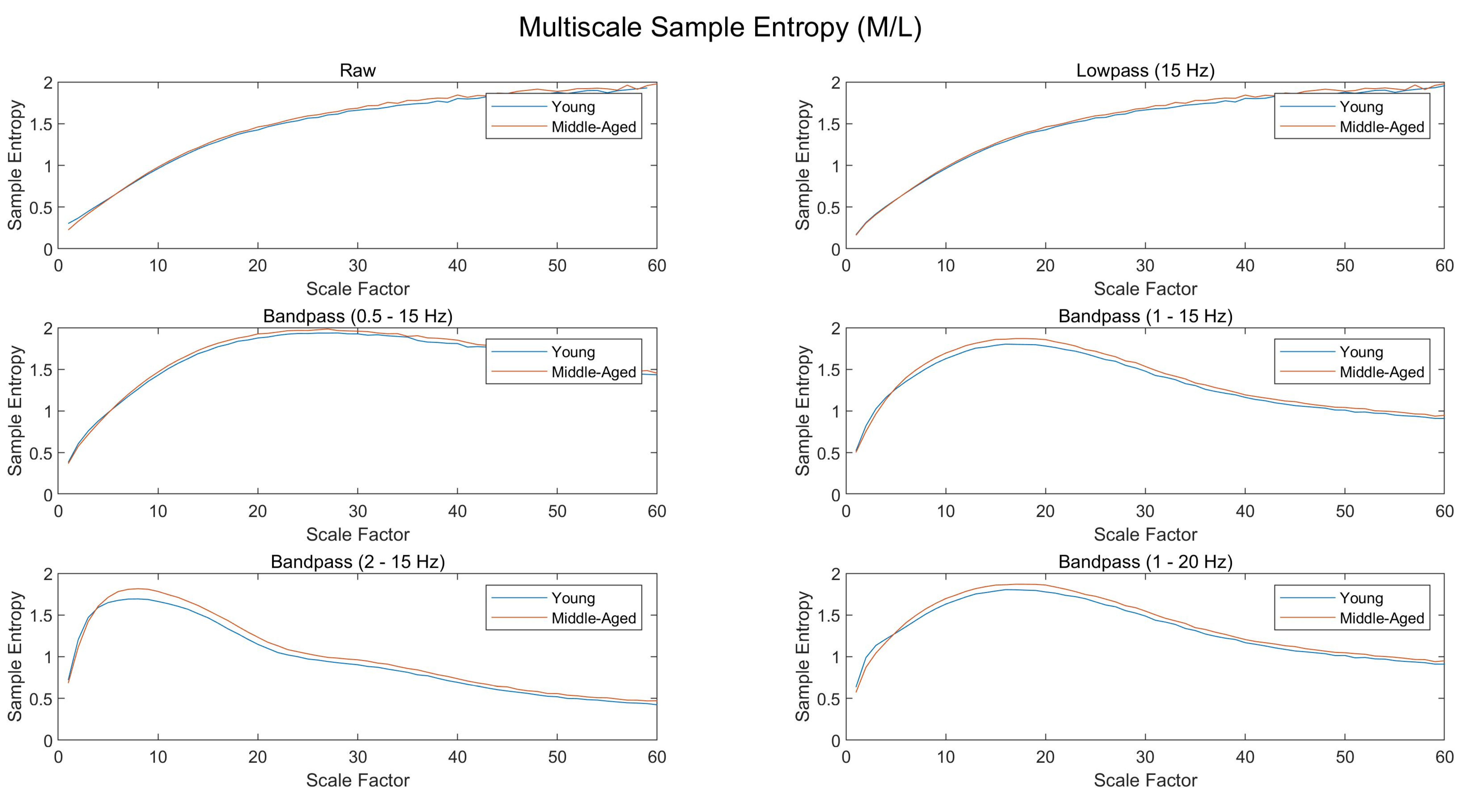
Ground reaction forces were collected continuously during each condition at 100 Hz and a custom written MATLAB script (The MathWorks, Natick, MA, USA) was used to compute the center of pressure (CoP). A second-order, zero-lag, bandpass Butterworth digital filter (1–15 Hz) was then applied to the data based on a sensitivity analysis (see
Appendix B).
Multiscale Sample Entropy was computed for the CoP data in the following manner (
Figure 1). First, the original CoP time series was coarse-grained according to the following equation:
where
is a sample of the coarse-grained signal,
is the scale, and
is a sample of the original signal. This procedure was repeated for a total of 40 scales, with each scale greater than one yielding a new, coarse-grained signal. Sample Entropy was then computed for each of the 40 scales by the following equation:
where
is the number of samples being compared across segments of the signal,
is the radius of similarity, and
is the length of the time series. Here,
is the probability that the segments will be sufficiently similar when comprised of
samples, and
is the probability that the segments will be sufficiently similar when comprised of
samples. Note that in the present study,
was set to 0.15 times the standard deviation of the signal and
was set to 2 [
12].
After computing the Sample Entropy for each scale, Complexity Indices were computed by taking the sum of the Sample Entropy values within a given range of scales, as defined by the following equation:
where
is the Sample Entropy for scale
and the Complexity Index is comprised of scales
b through
. Here, three Complexity Indices were selected based on their corresponding timescales: an
overall Complexity Index comprised of scales 1 through 40 (corresponding to 2.5–100 Hz), a
low-frequency Complexity Index comprised of scales 17 to 40 (corresponding to 2.5–6 Hz), and a
high-frequency Complexity Index comprised of scales 8 to 12 (corresponding to 8–12 Hz). Note that frequencies of 2.5–6 Hz are typically associated with voluntary movement, while frequencies of 8–12 Hz are typically associated with involuntary movement (e.g., physiological tremor).
2.4. Statistical Analysis
Two-way repeated measures analyses of variance (ANOVAs) with condition and age as within- and between-subjects factors, respectively, were performed for the three Complexity Indices (overall, low-frequency, and high-frequency) and for the two CoP directions (anteroposterior (A/P) and mediolateral (M/L)). Greenhouse–Geisser corrections were applied in any instance where the assumption of sphericity was violated (Mauchly’s W; p < 0.05). Post hoc pairwise comparisons were performed using t-tests with Bonferroni corrections. Effect sizes were reported in partial eta-squared (η2p) (0.01 = small; 0.06 = medium; 0.14 = large) for the ANOVAs and in Cohen’s d (0.20 = small; 0.50 = moderate; 0.80 = large) for the post hoc comparisons. A significance threshold of α = 0.05 was used for all inferential statistics. All statistical analyses were performed in JASP (JASP, Amsterdam, The Netherlands).
4. Discussion
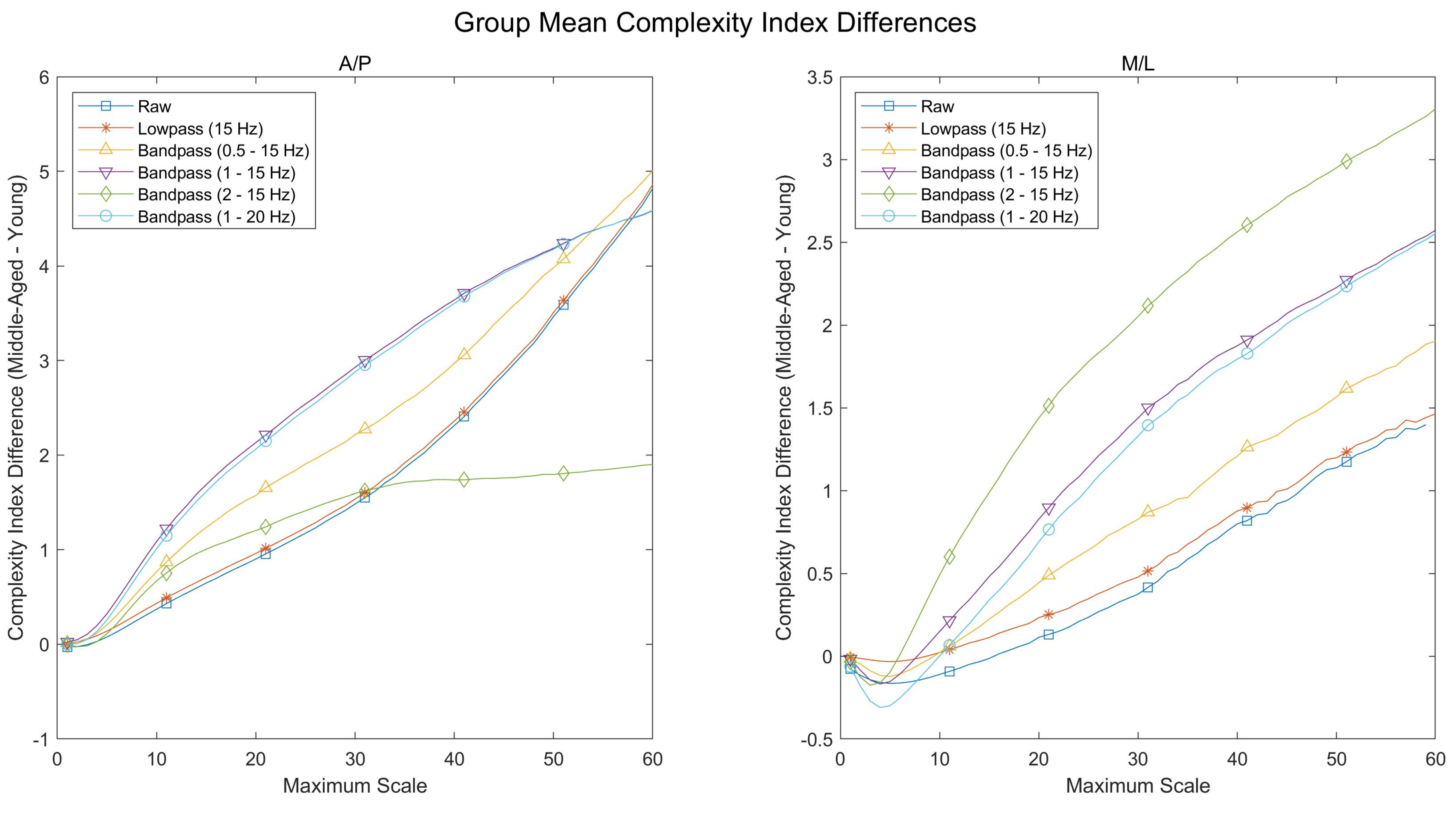
The present study assessed the complexity of postural behavior exhibited by young and middle-aged participants during a listening task of varying degrees of difficulty. Consistent with the loss of complexity hypothesis, it was expected that the young participants would exhibit greater postural complexity than the middle-aged participants, and that this difference would increase as a function of task difficulty. In comparing the postural complexity exhibited by the two groups of participants, however, the opposite result was observed; the middle-aged participants exhibited greater postural complexity across all Complexity Indices and across both CoP directions. Moreover, this difference appeared to increase as a function of task difficulty, with significant age-related differences in postural complexity emerging consistently in the −6 dB condition while not being present in the baseline or 0 dB conditions.
While these results were not expected, they are not entirely without precedent. In 2007, Costa et al. [
13] found that postural complexity did not differ significantly between young and older individuals during quiet standing, provided the individuals in the latter group were of sufficient health (i.e., had no history of falling). Rather, it was found that healthy young and healthy older individuals both exhibited greater postural complexity than did older individuals with a history of falling. As the middle-aged participants in the present study were screened for visual, vestibular, and motor impairments, it is reasonable that no significant age-related differences in postural complexity were observed in the baseline or 0 dB conditions. Indeed, these participants were first recruited by Helfer et al. [
11] in an effort to investigate age-related changes in postural control and speech perception that may appear despite a lack of associated impairments.
Additionally, Duarte and Sternad [
14] found that older individuals exhibited greater postural complexity than did young adults during prolonged standing. Importantly, however, these age-related differences were found to depend upon the radius of similarity,
, expressed in Equation (2). As prolonged standing
is accompanied by natural postural adjustments (e.g., shifting one’s weight),
the resulting signal often contains a substantial number of outliers. If one
group performs more frequent postural adjustments, these outliers may lead to
differences in signal complexity, as
r is
typically multiplied by the standard deviation of the signal. When Duarte and
Sternad corrected for these outliers by using a fixed
r value, these
age-related differences in postural complexity were no longer found.
Does such a finding by Duarte and Sternad suggest
that the age-related differences in the present study are similarly dependent
upon the radius of similarity (
r)?
First, one must consider the nature of the postural tasks in question. While
the prolonged standing task implemented by Duarte and Sternad consisted of
unconstrained standing lasting thirty minutes, the postural task in the present
study lasted only eighty seconds. It is therefore unlikely that the postural
data analyzed in the present study contained a similar number of outliers.
Moreover, the standard deviations in the present study were, on average,
greater in the middle-aged participants [
11].
As a greater standard deviation increases the radius of similarity, using a
fixed
r value in the present
study would serve only to
increase the relative degree of postural complexity observed in the middle-aged participants.
How, then, might one reconcile the nature of these differences with the existing literature on postural complexity and aging? As significant age-related differences in postural complexity emerged consistently in the −6 dB condition, it is possible that the two groups of participants adopted two distinct strategies in response to the listening task and that these strategies became distinguishable only under more difficult acoustic conditions. If, for example, maintaining upright posture while engaging with the listening task proved more difficult for the middle-aged participants, these participants may have been unable to incur any reduction in the degrees of freedom being used for postural control [
15]. In contrast, the young participants may have prioritized the perception of auditory information, even if this prioritization resulted in such a reduction in the degrees of freedom. This explanation is further supported by the finding that the postural complexity exhibited by the middle-aged participants did not differ between conditions, while the postural complexity exhibited by the young participants appeared to decrease as a function of task difficulty (
Figure 2,
Figure 3 and
Figure 4).
Returning to the first study conducted by Helfer et al. [
11], a moderate reduction in listening task performance was observed between the young and middle-aged participants. While this reduction may have been the result of early age-related changes in hearing, it is consistent with the notion that individuals in the latter group may have been unable to prioritize the perception of auditory information. Helfer et al. also reported a moderate increase in the 95% confidence ellipse of the CoP between the young and middle-aged participants during the listening task, further suggesting that maintaining upright posture while engaging with the listening task was more difficult for the middle-aged participants. These results, when combined with those of the present study, suggest that early aging may be accompanied by a decrease in the ability of individuals to regulate their CoP while conversing with others.
One additional finding is that the age-related differences in postural complexity appeared consistent across the three Complexity Indices. Within each Complexity Index, middle-aged participants exhibited significantly greater postural complexity in both the A/P and M/L directions than did young adults in the −6 dB condition (
Figure 2,
Figure 3 and
Figure 4). Likewise, moderate-to-large effect sizes were found within each Complexity Index in the 0 dB condition, suggesting that the middle-aged participants exhibited greater A/P postural complexity than did the young participants, though these differences did not reach statistical significance. Note that direct comparison of postural complexity
between Complexity Indices is not possible given the different number of scales within each Complexity Index (e.g., the overall Complexity Index is comprised of 40 scales while the high-frequency Complexity Index is comprised of only five scales).
While the results presented here provide unique insight into the dynamics of the postural system during listening, there are several limitations that should be noted. Firstly, participants in the present study were restricted to quiet standing, which may not be reflective of the multitude of postures individuals adopt while conversing. Secondly, the physical activity levels of the participants were not assessed despite these levels being potentially relevant to the interpretation of our results. Lastly, as noted by Helfer et al. [
11], the baseline condition was measured in relative silence, which may have altered postural behavior by rendering participants unable to orient themselves towards a reference signal in the surrounding environment [
16].
Overall, the results of the present study do not support the loss of complexity hypothesis put forth by Lipsitz and Goldberger [
1] but rather the notion that age-related differences in complexity are task dependent [
15]. Specifically, it was found that the postural complexity exhibited by young and middle-aged adults may not differ significantly during quiet standing if the individuals in the latter group are of sufficient health. Moreover, it was found that middle-aged adults may exhibit
greater postural complexity than young adults when quiet standing is coupled with a perceptual task. These results should serve to caution against universally associating the degree of complexity exhibited by a physiological system with the functional capacity of that system, as the degree of complexity may differ between tasks and ecological conditions.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}






