
Productivity vs. Evenness in the U.S. Financial Market: A Business Ecosystem Perspective


Abstract
:1. Introduction
2. The Business Ecosystem Perspective: Financial Markets as Ecosystems
| Financial market | Community ecology | Denoted by | |
| ↔ | species | i |
| ↔ | species biomass | vi |
| ↔ | total biomass (all species) | V |
| ↔ | rate of variation of the total biomass | R |
| ↔ | frequency of such species | xi |
| ↔ | evenness (species diversity) | E |
3. Empirical Analysis
3.1. Dataset
3.2. Variables
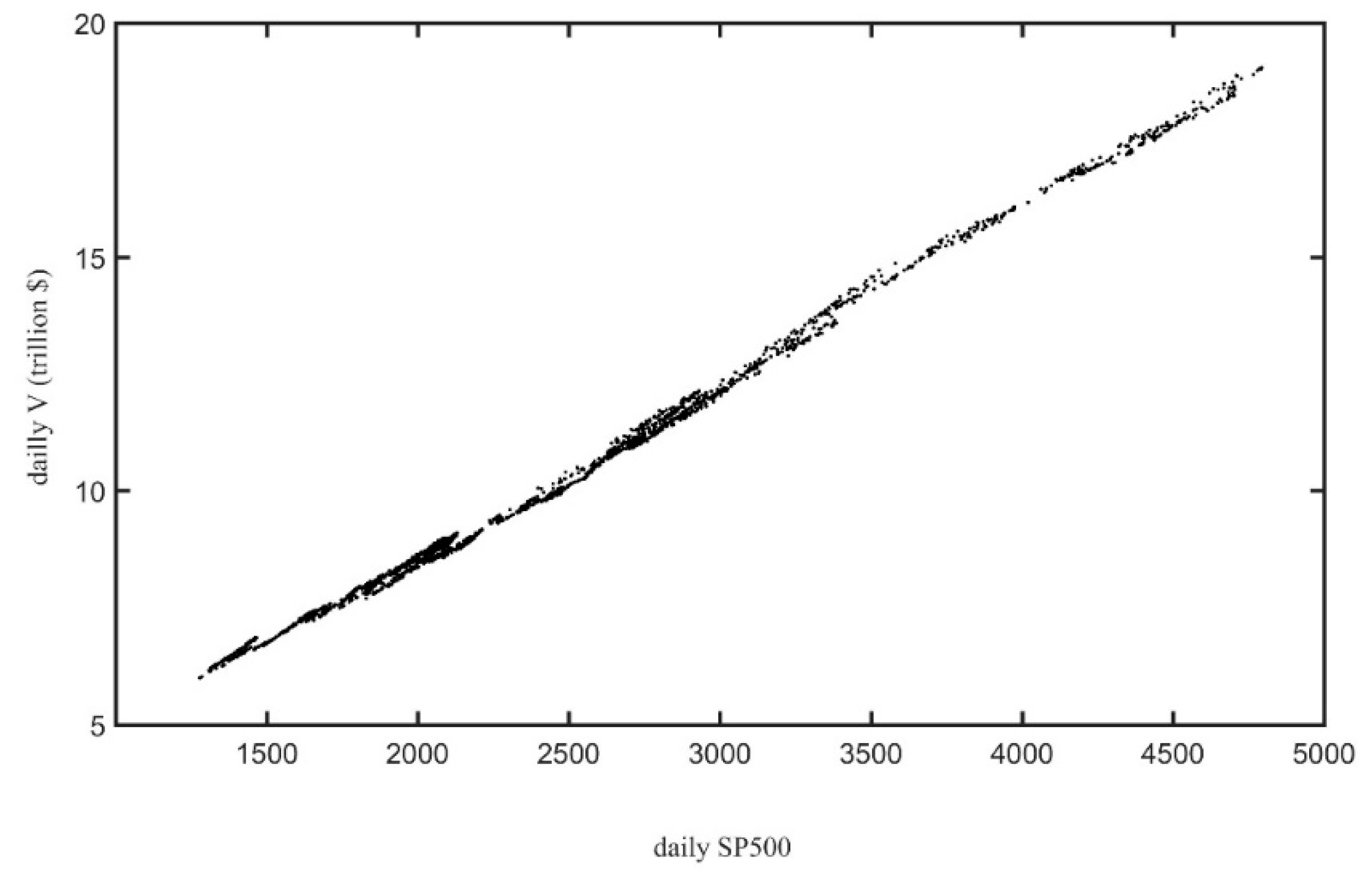
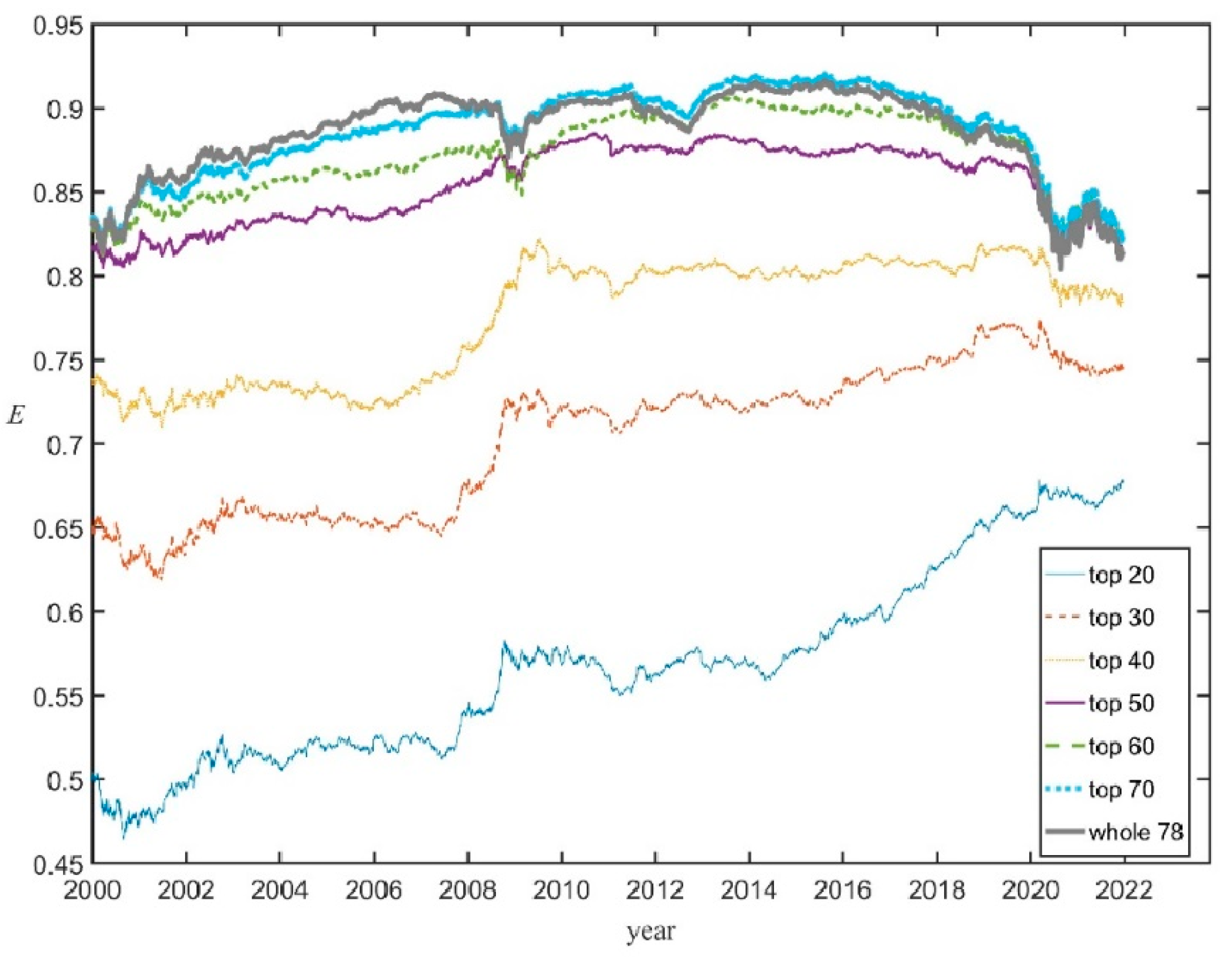
3.3. Productivity vs. Evenness in the U.S. Stock Market
- A period of soaring E(t), from January 2001 to December 2007, (almost exactly coinciding with the first business cycle);
- A period of relatively smooth oscillations of E(t) around a high value, from January 2008 to December 2017;
- A period in which E plunged, from January 2018 to December 2021.

- From the beginning of 2001 until the end of 2007 (portion of the trajectory in blue in Figure 4), in which E steadily increased, while Va ended in a slightly lower value.
- The 2009–2020 expansion (portion of the trajectory in red in Figure 4), which was the longest on record at 128 months—from July 2009 to February 2020—according to the Congressional Research Service (NBER 2022). This was a period in which, after some initial erratic movements, Va grew strongly and E considerable declined.

4. Explaining the Relationship between Productivity and Evenness in Stock Markets from a Community Ecology Perspective
4.1. Ensemble of Synthetic Communities
4.2. Other Ecologically Based Approaches Supporting the Negative Relationship between Productivity and Evenness
5. Conclusions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| EE | Ecological Economics |
| ENT | Ecological Niche Theory |
| GICS | Global Industry Classification Standard |
| LVGT | Lotka–Volterra generalized theory |
| NYSE | New York Stock Exchange |
| PME | Pairwise Maximum-Entropy |
Appendix A. Other Metrics of Evenness

Appendix B. The Evenness of a Randomly Distributed Shares

Appendix C. The Total Market Value as a Function of δ


Appendix D. Effective U.S. FED Rates Compared with the Total Market Value and Evenness

References
- Simon, H.A. A Behavioral Model of Rational Choice. Q. J. Econ. 1955, 69, 99–118. [Google Scholar] [CrossRef]
- Shiller, R.J. Market Volatility; MIT Press: Cambridge, MA, USA, 1997. [Google Scholar]
- Alderson, W. Marketing Behavior and Executive Action: A Functionalist Approach to Marketing Theory; Homewood: Irwin, IL, USA, 1957. [Google Scholar]
- Alderson, W. Dynamic Marketing Behavior: A Functionalist Theory of Marketing; Homewood: Irwin, IL, USA, 1965. [Google Scholar]
- Potters, M.; Cont, R.; Bouchaud, J.P. Financial Markets as Adaptive Ecosystems. Europhys. Lett. 2004, 41, 239. [Google Scholar] [CrossRef] [Green Version]
- Manahov, V.; Hudson, R. A note on the relationship between market efficiency and adaptability. New evidence from artificial stock markets. Expert Syst. Appl. Int. J. 2014, 41, 7436–7454. [Google Scholar] [CrossRef]
- Fort, H. Forecasting with Maximum Entropy: The Interface between Physics, Biology, Economics and Information Theory; Institute of Physics Publishing: Bristol, UK, 2022. [Google Scholar]
- Moore, J.F. Predators and Prey: A new ecology of competition. Harv. Bus. Rev. 1993, 71, 75–86. [Google Scholar]
- Morin, P. Community Ecology; Wiley-Blackwell: Chichester, UK, 2011. [Google Scholar]
- Moore, J.F. The Death of Competition: Leadership and Strategy in the Age of Business Ecosystems; Harper Paperbacks: New York, NY, USA, 1993. [Google Scholar]
- Farmer, D.J.; Lo, A.W. Frontiers of finance: Evolution and efficient markets. Proc. Natl. Acad. Sci. USA 1999, 96, 9991–9992. [Google Scholar] [CrossRef]
- Krebs, C.J. Ecology: The Experimental Analysis of Distribution and Abundance, 2nd ed.; Harper & Row: New York, NY, USA, 1978. [Google Scholar]
- Grime, J.P.; Pierce, S. The Evolutionary Strategies That Shape Ecosystems; Wiley: Chichester, UK, 2012. [Google Scholar]
- Mittelbach, G.G.; Steiner, C.F.; Scheiner, S.M.; Gross, K.L.; Reynolds, H.L.; Waide, R.B.; Willig, M.R.; Dodson, S.I.; Gough, L. What is the observed relationship between species richness and productivity? Ecology 2001, 82, 2381–2396. [Google Scholar] [CrossRef]
- Cardinale, B.J.; Hillebrand, H.; Harpole, W.S.; Gross, K.L.; Ptacnick, R. Separating the influence of resource ‘availability’ from resource ‘imbalance’ on productivity–diversity relationships. Ecol. Lett. 2009, 12, 475–487. [Google Scholar] [CrossRef]
- Allaby, M. (Ed.) A Dictionary of Ecology, 3rd ed.; Oxford University Press: Oxford, UK, 2006. [Google Scholar]
- Vandermeer, J.H. The Ecology of Intercropping; Cambridge University Press: Cambridge, UK, 1989. [Google Scholar]
- Hayes, A. What Is Crop Yield? Investopedia. Available online: https://www.investopedia.com/terms/c/crop-yield.asp (accessed on 14 November 2022).
- Magurran, A.E. Measuring Biological Diversity; Blackwell Science: Malden, MA, USA, 2004. [Google Scholar]
- Chiarucci, A.; Bacaro, G.; Scheiner, S.M. Old and new challenges in using species diversity for assessing biodiversity. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 2426–2437. [Google Scholar] [CrossRef] [Green Version]
- Costanza, R. What is ecological economics? Ecol. Econ. 1989, 1, 1–7. [Google Scholar] [CrossRef]
- Potts, J.; Foster, J.; Straton, A. An entrepreneurial model of economic and environmental co-evolution. Ecol. Econ. 2010, 70, 375–383. [Google Scholar] [CrossRef] [Green Version]
- Farmer, D.J. Market Force, Ecology and Evolution. Ind. Corp. Chang. 2002, 11, 895–953. [Google Scholar] [CrossRef]
- Moore, J.F. Business Ecosystems and the View from the Firm. Antitrust Bull. 2006, 51, 31–75. [Google Scholar] [CrossRef] [Green Version]
- Hileman, J.; Kallstenius, I.; Häyhä, T.; Palm, C.; Cornell, S. Keystone actors do not act alone: A business ecosystem perspective on sustainability in the global clothing industry. PLoS ONE 2020, 15, e0241453. [Google Scholar] [CrossRef]
- Pielou, E. An Introduction to Mathematical Ecology; Wiley: New York, NY, USA, 1969. [Google Scholar]
- Van Dyke, F. Conservation Biology: Foundations, Concepts, Applications, 2nd ed.; Springer: New York, NY, USA, 2008. [Google Scholar]
- Maignan, C.; Ottaviano, G.; Pinelli, D.; Rullani, F. Bio-Ecological Diversity vs. Socio-Economic Diversity: A Comparison of Existing Measures Nota di Lavoro. 2003. Available online: http://www.feem.it/web/activ/_wp.html (accessed on 15 March 2023).
- Baumgärtner, S. Measuring the Diversity of What? And for What Purpose? A Conceptual Comparison of Ecological and Economic Biodiversity Indices. 2006. Available online: https://ssrn.com/abstract=894782 (accessed on 15 March 2023).
- Boyte-White, C. Market Capitalization Versus Market Value: What’s the Difference? Investopedia. Available online: https://www.investopedia.com/ask/answers/122314/what-difference-between-market-capitalization-and-market-value.asp (accessed on 9 November 2022).
- Huston, M. A general hypothesis of species diversity. Am. Nat. 1979, 113, 81–101. [Google Scholar] [CrossRef]
- Huston, M.A.; Aarssen, L.W.; Austin, M.P.; Cade, B.S.; Fridley, J.D.; Garnier, E.; Grime, J.P.; Hodgson, J.; Lauenroth, W.K.; Wardle, D.A. No consistent effect of plant diversity on productivity. Science 2000, 289, 1255. [Google Scholar] [CrossRef] [Green Version]
- Schmid, B. The species richness-productivity controversy. Trends Ecol. Evol. 2002, 17, 113–114. [Google Scholar] [CrossRef]
- Lotka, A.J. Elements of Physical Biology; Williams & Wilkins: New York, NY, USA, 1925. [Google Scholar]
- Volterra, V. Fluctuations in the abundance of a species considered mathematically. Nature 1926, 118, 558–560. [Google Scholar] [CrossRef] [Green Version]
- Hofbauer, M.; Sigmund, K. Evolutionary Games and Population Dynamics; Cambridge University Press: Cambridge, UK, 1998. [Google Scholar]
- Fort, H. Ecological Modelling and Ecophysics: Agricultural and Environmental Applications; Institute of Physics Publishing: Bristol, UK, 2020. [Google Scholar]
- May, R.M. Will a large complex system be stable? Nature 1972, 238, 413–414. [Google Scholar] [CrossRef]
- Chesson, P. Mechanisms of maintenance of species diversity. Ann. Rev. Ecol. Evol. Syst. 2000, 31, 343–366. [Google Scholar] [CrossRef] [Green Version]
- Fortune Magazine. Available online: https://fortune.com/rankings/ (accessed on 9 November 2022).
- Core US Fundamentals Dataset. Available online: http://www.quandl.com/databases/SF1 (accessed on 15 March 2023).
- Wakil, G. Firm size proxies and the value relevance of predictive stock return models. J. Econ. Financ. 2020, 44, 434–457. [Google Scholar] [CrossRef]
- Hamilton, J. Dates of U.S. Recessions as Inferred by GDP-Based Recession Indicator JHDUSRGDPBR, Retrieved from FRED, Federal Reserve Bank of St. Louis. Available online: https://fred.stlouisfed.org/series/JHDUSRGDPBR (accessed on 12 October 2022).
- Money Morning. The Dot-Com Crash of 2000–2002. Available online: https://moneymorning.com/2015/06/12/the-dot-com-crash-of-2000-2002/ (accessed on 23 November 2022).
- NYT 2007. Bear Stearns Reports First Ever Quarterly Loss. Available online: https://archive.nytimes.com/dealbook.nytimes.com/2007/12/20/bear-reports-steep-but-expected-4th-quarter-loss/?_r=0 (accessed on 10 February 2023).
- NYSE 2022. Available online: https://www.nyse.com/market-cap (accessed on 10 July 2022).
- Kenton, W. What Is Trumponomics. Investopedia. Available online: https://www.investopedia.com.cach3.com/terms/t/trumponomics.asp.html (accessed on 14 November 2022).
- Mulder, C.P.; Bazeley-White, E.; Dimitrakopoulos, P.G.; Hector, A.; Scherer-Lorenzen, M.; Schmid, B. Species Evenness and Productivity in Experimental Plant Communities. Oikos 2004, 107, 50–63. [Google Scholar] [CrossRef]
- Halty, V.; Valdés, M.; Tejera, M.; Picasso, V.; Fort, H. Modelling plant interspecific interactions from experiments of perennial crop mixtures to predict optimal combinations. Ecol. Appl. 2017, 27, 2277–2289. [Google Scholar] [CrossRef]
- Vandermeer, J.H. The competitive structure of communities: An experimental approach with protozoa. Ecology 1969, 50, 362–371. [Google Scholar] [CrossRef] [Green Version]
- Huisman, J.; Jonker, R.R.; Zonneveld, C.; Weissing, F.J. Competition for light between phytoplankton species: Experimental tests of mechanistic theory. Ecology 1999, 80, 211–222. [Google Scholar] [CrossRef]
- Connel, J.H. Apparent vs. “rea” competition in plants. In Perspectives on Plant Competition; Grace, J.B., Tilman, D., Eds.; Academic Press: New York, NY, USA, 1990; pp. 9–26. [Google Scholar]
- Miller, T.E. Direct and indirect species interactions in an early old-field plant community. Am. Nat. 1994, 143, 1007–1025. [Google Scholar] [CrossRef]
- Adler, F.R.; Morris, W.F. A general test for interaction modification. Ecology 1994, 75, 1552–1559. [Google Scholar] [CrossRef]
- Fort, H. On predicting species yields in multispecies communities: Quantifying the accuracy of the linear Lotka-Volterra generalized model. Ecol. Model. 2018, 387, 154–162. [Google Scholar] [CrossRef]
- Callaway, R.M.; Walker, L.R. Competition and facilitation: A synthetic approach to interactions in plant communities. Ecology 1997, 78, 1958–1965. [Google Scholar] [CrossRef]
- Silknetter, S.; Creed, R.P.; Brown, B.L.; Frimpong, E.A.; Skelton, J.; Peoples, B.K. Positive biotic interactions in freshwaters: A review and research directive. Freshw. Biol. 2020, 65, 811–832. [Google Scholar] [CrossRef] [Green Version]
- Yodzis, P. The stability of real ecosystems. Nature 1981, 289, 674–676. [Google Scholar] [CrossRef]
- Federal Funds Effective Rate. Retrieved from FRED, Federal Reserve Bank of St. Louis. Available online: https://fred.stlouisfed.org/series/ (accessed on 2 December 2022).
- Gamfeldt, L.; Hillebrand, H. Effects of Total Resources, Resource Ratios, and Species Richness on Algal Productivity and Evenness at Both Metacommunity and Local Scales. PLoS ONE 2011, 6, e21972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chase, J.M.; Leibold, M.A. Ecological Niches; University of Chicago Press: Chicago, IL, USA, 2003. [Google Scholar]
- Fort, H. Community diversity and total abundance: Quantitative predictions from competition niche theory. Ecol. Complex. 2015, 21, 120–127. [Google Scholar] [CrossRef]
- Fort, H. Combining niche and game theories to address interspecific cooperation in ecological communities. Community Ecol. 2020, 21, 13–24. [Google Scholar] [CrossRef]
- Roudi, Y.; Nirenberg, S.; Latham, P.E. Pairwise Maximum Entropy Models for Studying Large Biological Systems: When They Can Work and When They Can’t. PLoS Comput. Biol. 2009, 5, e1000380. [Google Scholar] [CrossRef] [PubMed]
- Jaynes, E.T. Information theory and statistical mechanics I. Phys. Rev. 1957, 106, 620–630. [Google Scholar] [CrossRef]
- Jaynes, E.T. Information theory and statistical mechanics II. Phys. Rev. 1957, 108, 171–190. [Google Scholar] [CrossRef]
- Vinod, H.D. Ranking mutual funds using unconventional utility theory and stochastic dominance. J. Empir. Financ. 2004, 11, 353–377. [Google Scholar] [CrossRef]
- Rompolis, L. Retrieving risk neutral densities from European option prices based on the principle of maximum entropy. J. Empir. Financ. 2010, 17, 918–937. [Google Scholar] [CrossRef]
- Balasubramanyan, L.; Stefanou, S.E.; Stokes, J.R. An entropy approach to size and variance heterogeneity in U.S. commercial banks. J. Econ. Financ. 2012, 36, 728–749. [Google Scholar] [CrossRef]
- Bialek, W.; Cavagna, A.; Giardina, I.; Mora, T.; Silvestri, E.; Viale, M.; Walczak, A.M. Statistical mechanics for natural flocks of birds. Proc. Natl. Acad. Sci. USA 2012, 109, 4786–4791. [Google Scholar] [CrossRef]
- Volkov, I.; Banavar, J.R.; Hubbell, S.P.; Maritan, A. Inferring species interactions in tropical forests. Proc. Natl. Acad. Sci. USA 2009, 106, 13854–13859. [Google Scholar] [CrossRef]
- Fort, H.; Grigera, T. A method for predicting species trajectories tested with trees in Barro Colorado tropical forest. Ecol. Model. 2021, 446, 109504. [Google Scholar] [CrossRef]
- Fort, H.; Grigera, T. A new early warning indicator of tree species crashes from effective intraspecific interactions in tropical forests. Ecol. Indic. 2021, 125, 107506. [Google Scholar] [CrossRef]
- Emary, C.; Fort, H. Markets as ecological networks: Inferring interactions and identifying communities. J. Complex Netw. 2021, 9, cnab022. [Google Scholar] [CrossRef]
- The Global Industry Classification Standard (GICS). Available online: https://www.msci.com/our-solutions/indexes/gics (accessed on 25 May 2023).
- Adomavicius, G.; Bockstedt, J.C.; Gupta, A.; Kauffman, R.J. Technology roles and paths of influence in an ecosystem model of technology evolution. Inf. Technol. Manag. 2007, 8, 185–202. [Google Scholar] [CrossRef]
- Drożdż, S.; Grümmer, F.; Górski, A.Z.; Ruf, F.; Speth, J. Dynamics of competition between collectivity and noise in the stock market. Phys. A 2000, 287, 440–449. [Google Scholar] [CrossRef] [Green Version]
- Triplett, R.; Ozdemir, N.; Mason, P. Structural Change in the Investment Function. J. Econ. Financ. 2022, 46, 220–236. [Google Scholar] [CrossRef]
- Fort, H. A quantitative assessment of the evolutionary dynamics of firms: Estimating the fitnesses from time series of their market caps and forecasting their shares. J. Evol. Econ. 2023; submitted. [Google Scholar]
- Fort, H. Disentangling the effects of selection and the environment in the evolutionary dynamics of financial markets in terms of Malthusian fitness. Work in progress, to be published elsewhere.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Company | Ticker | Market Val (USD Bill) | Rank | Sector | Industry |
|---|---|---|---|---|---|
| Apple | AAPL | 2902 | 1 | Technology | Consumer Electronics |
| Microsoft | MSFT | 2522 | 2 | Technology | Software–Infrastructure |
| Amazon | AMZN | 1697 | 3 | Consumer Cyclical | Internet Retail |
| Berkshire Hathaway | BRK | 662.63 | 4 | Financial Services | Insurance |
| JP Morgan | JPM | 472.51 | 5 | Financial Services | Banks |
| United Health Group | UNH | 466.21 | 6 | Healthcare | Healthcare Plans |
| Johnson & Johnson | JNJ | 450.36 | 7 | Healthcare | Drug Manufacturers |
| Home Depot | HD | 433.37 | 8 | Consumer Cyclical | Home Retail |
| Walmart | WMT | 401.35 | 9 | Consumer Defensive | Discount Stores |
| P&G | PG | 392.11 | 10 | Consumer Defensive | Household |
| Bank of America | BAC | 359.38 | 11 | Financial Services | Banks |
| Pfizer Inc. | PFE | 331.86 | 12 | Healthcare | Drug Manufacturers |
| The Walt Disney Company | DIS | 281.54 | 13 | Comm. Services | Entertainment |
| Cisco Systems, Inc. | CSCO | 267.27 | 14 | Technology | Comm. Equipment |
| Nike | NKE | 263.55 | 15 | Consumer Cyclical | Footwear and Access. |
| Thermo Fisher Scientific Inc. | TMO | 263.18 | 16 | Healthcare | Diagnosis and Research |
| Exxon Mobil | XOM | 259.38 | 17 | Energy | Oil and Gas |
| The Coca-Cola Company | KO | 256.09 | 18 | Consumer Defensive | Beverages |
| Costco | COST | 251.74 | 19 | Consumer Defensive | Discount Stores |
| Abbott Laboratories | ABT | 248.28 | 20 | Healthcare | Medical Devices |
| PepsiCo, Inc. | PEP | 240.24 | 21 | Consumer Defensive | Beverages |
| Oracle | ORCL | 232.89 | 22 | Technology | Software–Infrastructure |
| Comcast | CMCSA | 228.16 | 23 | Comm. Services | Telecom Services |
| Chevron | CVX | 226.46 | 24 | Energy | Oil and Gas |
| Verizon | VZ | 218.12 | 25 | Comm. Services | Telecom Services |
| Intel Corporation | INTC | 209.6 | 26 | Technology | Semiconductors |
| QUALCOMM Incorporated | QCOM | 205.73 | 27 | Technology | Semiconductors |
| Merck & Co., Inc. | MRK | 193.72 | 28 | Healthcare | Drug Manufacturers |
| Wells Fargo | WFC | 186.44 | 29 | Financial Services | Banks |
| Anthem | UPS | 186.41 | 30 | Industrials | Integrated Freight and Logistics |
| Lowe’s | LOW | 174.15 | 31 | Consumer Cyclical | Home Retail |
| Morgan Stanley | MS | 173.96 | 32 | Financial Services | Banks |
| Honeywell International Inc. | HON | 142.79 | 33 | Industrials | Conglomerates |
| CVS Caremark | CVS | 136.38 | 34 | Healthcare | Healthcare Plans |
| Bristol-Myers Squibb Company | BMY | 134.24 | 35 | Healthcare | Drug Manufacturers |
| AT&T | T | 132.58 | 36 | Comm. Services | Telecom Services |
| Raytheon Technologies Corp. | RTX | 128.51 | 37 | Industrials | Aerospace and Defense |
| The Goldman Sachs Group, Inc. | GS | 127.61 | 38 | Financial Services | Banks |
| American Express Company | AXP | 124.5 | 39 | Financial Services | Credit Services |
| IBM | IBM | 120.04 | 40 | Technology | Information Tech. Serv. |
| Citigroup | C | 119.84 | 41 | Financial Services | Banks |
| Boeing | BA | 118.56 | 42 | Industrials | Aerospace and Defense |
| Target | TGT | 110.89 | 43 | Consumer Defensive | Discount Stores |
| Caterpillar Inc. | CAT | 110.79 | 44 | Industrials | Farm and Heavy Constr. |
| Deere & Company | DE | 105.68 | 45 | Industrials | Farm and Heavy Constr. |
| General electrics | GE | 103.83 | 46 | Industrials | Specialty Industr. Machinery |
| 3M Company | MMM | 101.58 | 47 | Industrials | Conglomerates |
| Lockheed Martin Corporation | LMT | 96.32 | 48 | Industrials | Aerospace and Defense |
| ConocoPhillips | COP | 94 | 49 | Energy | Oil and Gas |
| Phillips 66 | TJX | 90.56 | 50 | Energy | Oil and Gas |
| Ford Motors | F | 85.59 | 51 | Consumer Cyclical | Auto Manufacturers |
| Cigna Corporation | CI | 74.16 | 52 | Healthcare | Healthcare Plans |
| FedEx Corporation | FDX | 68.53 | 53 | Industrials | Integrated Freight and Logistics |
| Northrop Grumman Corp. | NOC | 60.49 | 54 | Industrials | Aerospace and Defense |
| Capital One Financial Corp. | COF | 60.05 | 55 | Financial Services | Credit Services |
| The Progressive Corporation | PGR | 59.99 | 56 | Financial Services | Insurance |
| Humana Inc. | HUM | 59.75 | 57 | Healthcare | Healthcare Plans |
| General Dynamics | GD | 57.88 | 58 | Industrials | Aerospace and Defense |
| Enterprise Products Partners L.P. | EPD | 47.79 | 59 | Energy | Oil and Gas |
| AIG | AIG | 46.55 | 60 | Financial Services | Insurance |
| Walgreens Boots Alliance | WBA | 45.03 | 61 | Healthcare | Pharmaceutical Retailers |
| HP Inc. | HPQ | 40.79 | 62 | Technology | Computer Hardware |
| Exelon Corporation | EXC | 40.34 | 63 | Utilities | Utilities-Regulated Electric |
| Sysco Corporation | SYY | 40.27 | 64 | Consumer Defensive | Food Distribution |
| Archer-Daniels-Midland Comp. | ADM | 37.85 | 65 | Consumer Defensive | Farm Products |
| The Travelers Companies, Inc. | TRV | 37.73 | 66 | Financial Services | Insurance |
| McKesson Corp. | MCK | 37.24 | 67 | Healthcare | Medical Distribution |
| The Kroger Co. | KR | 33.28 | 68 | Consumer Defensive | Grocery Stores |
| The Allstate Corporation | ALL | 33.06 | 69 | Financial Services | Insurance |
| Tyson Foods, Inc. | TSN | 31.65 | 70 | Consumer Defensive | Farm Products |
| Nucor Corporation | NUE | 31.1 | 71 | Basic Materials | Steel |
| Valero Energy | VLO | 30.73 | 72 | Energy | Oil and Gas |
| AmerisourceBergen | ABC | 27.78 | 73 | Healthcare | Medical Distribution |
| Best Buy Co., Inc. | BBY | 24.44 | 74 | Consumer Cyclical | Specialty Retail |
| Cardinal Health | CAH | 14.26 | 75 | Healthcare | Medical Distribution |
| Arrow Electronics, Inc. | ARW | 9.14 | 76 | Technology | Electronics Distribution |
| Fannie Mae | FNMA | 0.95 | 77 | Financial Services | Mortgage Finance |
| Chico’s FAS, Inc. | CHS | 0.66 | 78 | Consumer Cyclical | Apparel Retail |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fort, H. Productivity vs. Evenness in the U.S. Financial Market: A Business Ecosystem Perspective. Entropy 2023, 25, 1029. https://doi.org/10.3390/e25071029
Fort H. Productivity vs. Evenness in the U.S. Financial Market: A Business Ecosystem Perspective. Entropy. 2023; 25(7):1029. https://doi.org/10.3390/e25071029
Chicago/Turabian StyleFort, Hugo. 2023. "Productivity vs. Evenness in the U.S. Financial Market: A Business Ecosystem Perspective" Entropy 25, no. 7: 1029. https://doi.org/10.3390/e25071029
APA StyleFort, H. (2023). Productivity vs. Evenness in the U.S. Financial Market: A Business Ecosystem Perspective. Entropy, 25(7), 1029. https://doi.org/10.3390/e25071029