
A Nonlinear Dynamical View of Kleiber’s Law on the Metabolism of Plants and Animals


Abstract
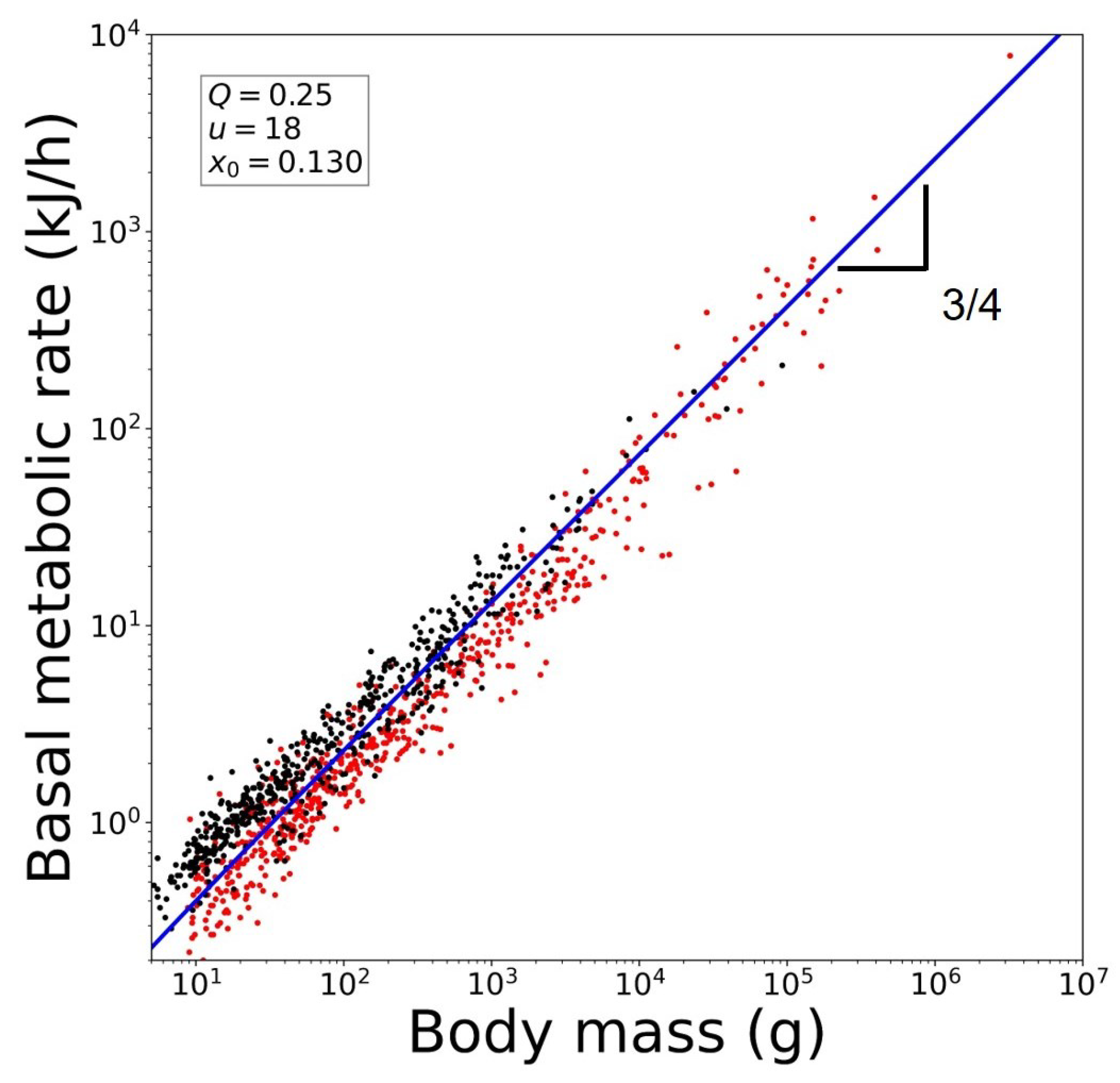
:1. Introduction
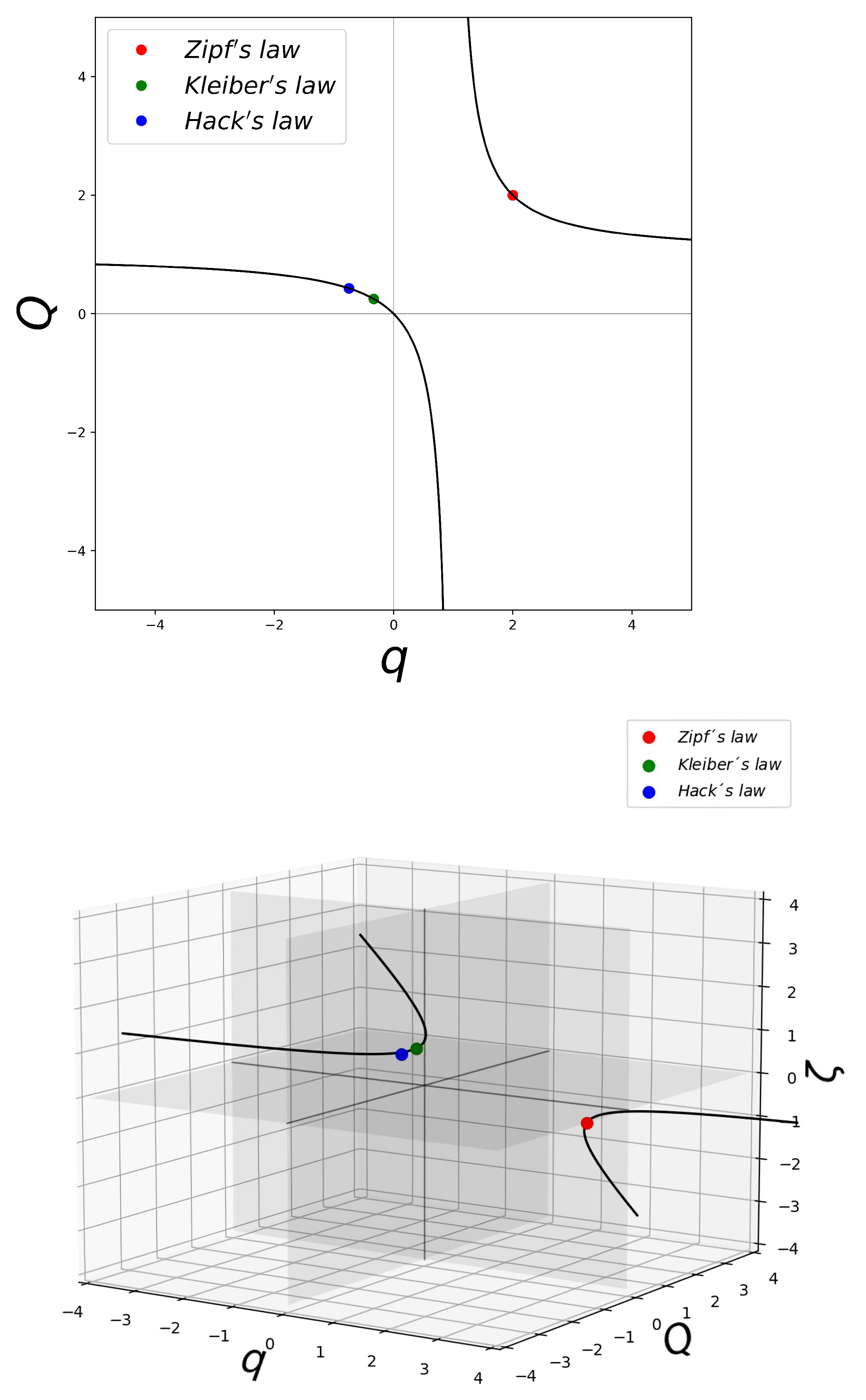
2. Rank Distributions and Their Universality Classes
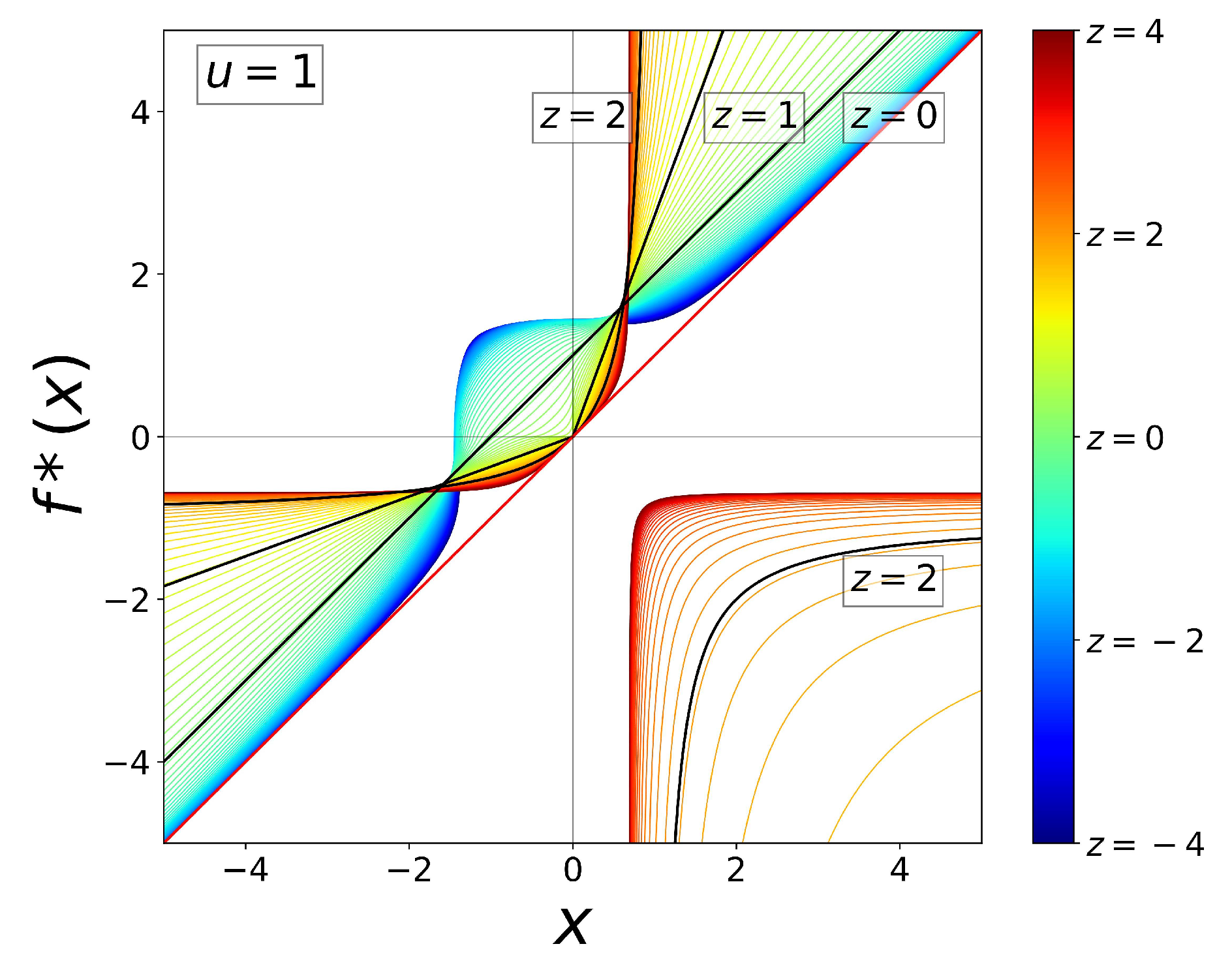
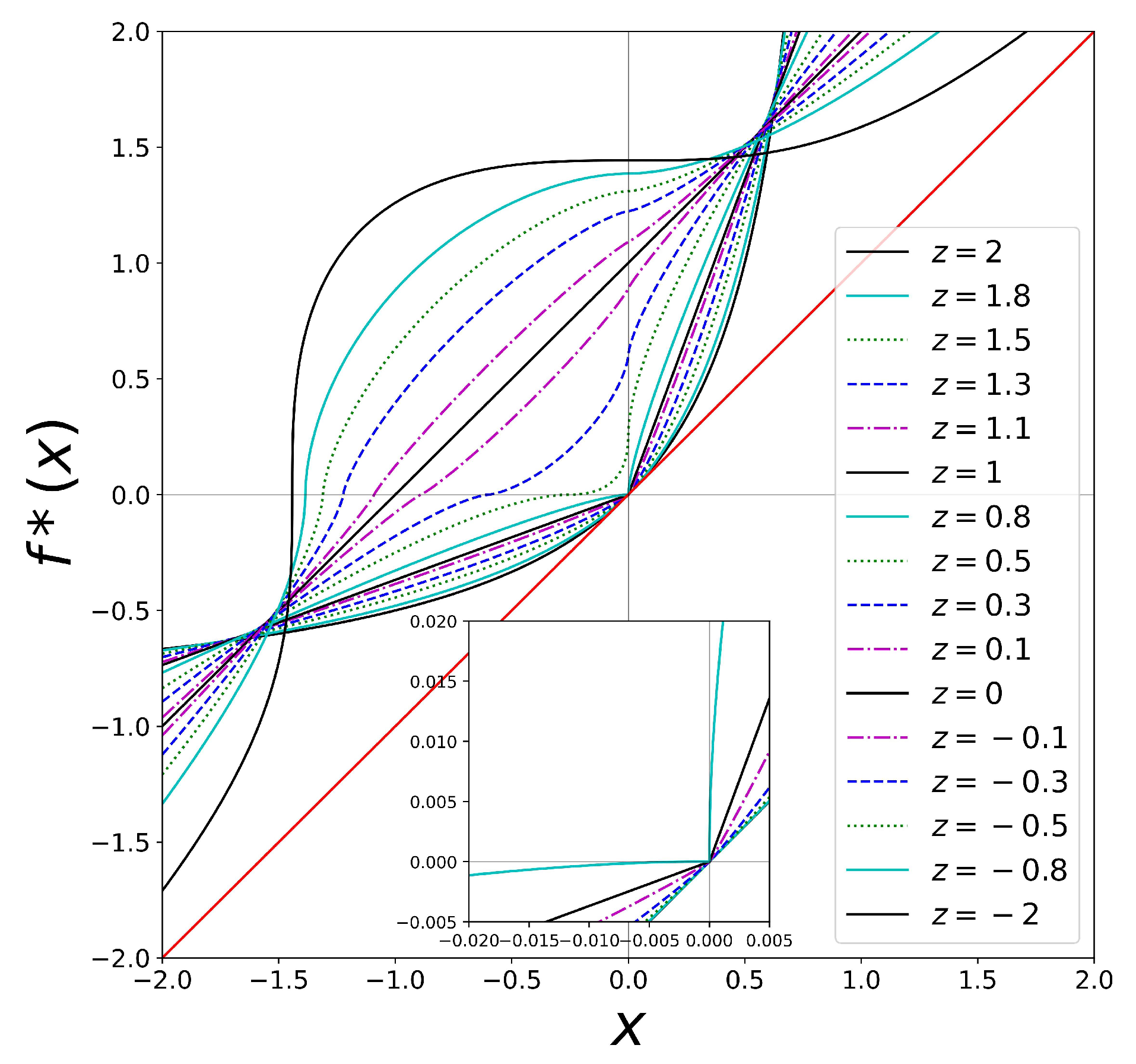
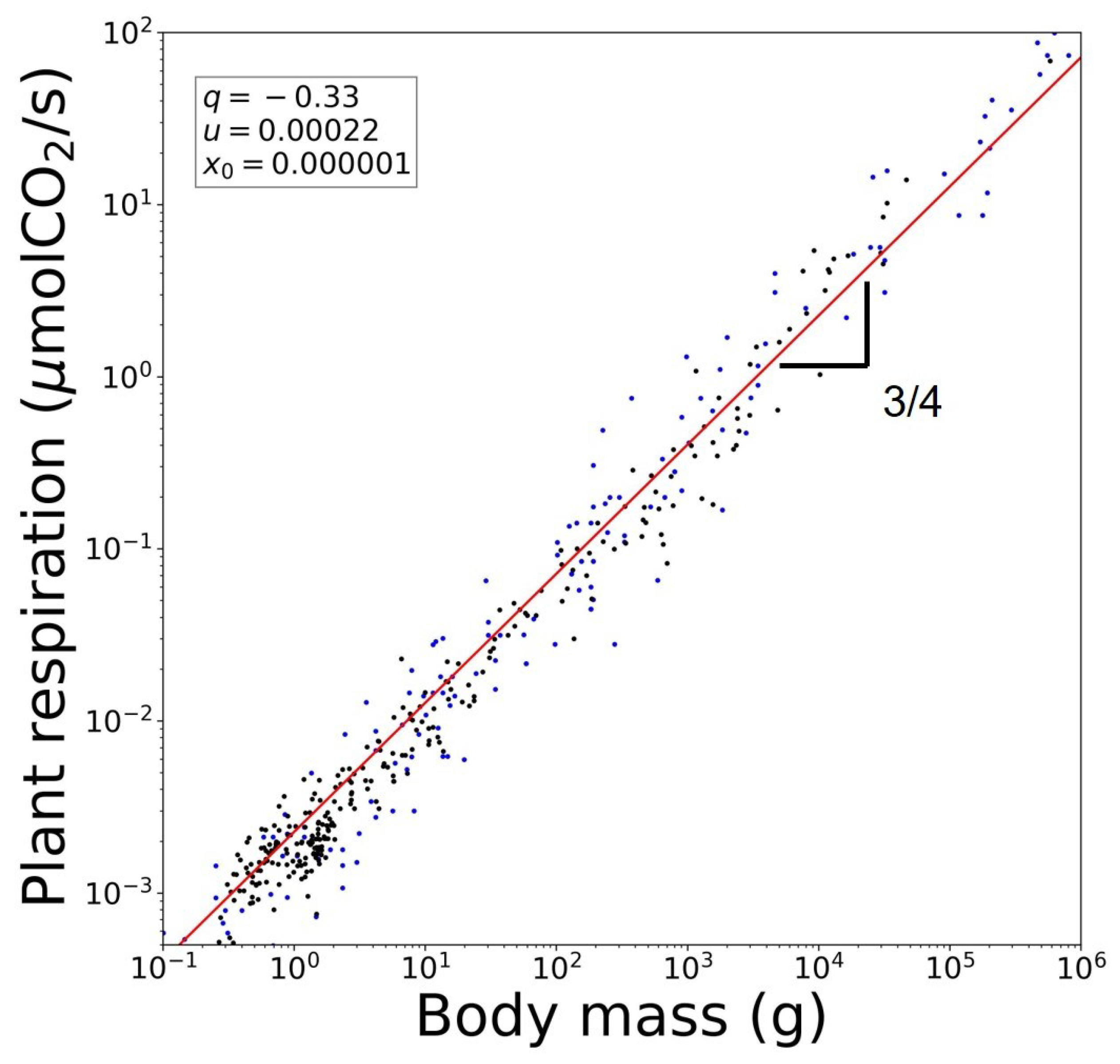
3. Scaling of Rates and Characteristic Times and Their Universality Classes
4. Rich Gets Richer and Effortless Motion

5. Summary and Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kleibers’s Law. Wikipedia. Available online: https://en.wikipedia.org/wiki/Kleiber%27s_law (accessed on 24 December 2023).
- Kleiber, M. Body size and metabolism. Hilgardia 1932, 6, 315–353. [Google Scholar] [CrossRef]
- Kleiber, M. Body size and metabolic rate. Physiol. Rev. 1947, 27, 511–541. [Google Scholar] [CrossRef] [PubMed]
- Kleiber, M. The Fire of Life. An Introduction to Animal Energetics; John Wiley & Sons, Inc.: New York, NY, USA; London, UK, 1961. [Google Scholar]
- Von Bertalanffy, L. Quantitative laws in metabolism and growth. Q. Rev. Biol. 1957, 32, 217–231. [Google Scholar] [CrossRef] [PubMed]
- West, G.; Brown, J.; Enquist, B. A general model for the origin of allometric scaling laws in biology. Science 1997, 276, 122–126. [Google Scholar] [CrossRef] [PubMed]
- West, G.; Brown, J.; Enquist, B. A general model for the structure and allometry of plant vascular systems. Nature 1999, 400, 664–667. [Google Scholar] [CrossRef]
- West, G.; Brown, J.; Enquist, B. The fourth dimension of life: Fractal geometry and allometric scaling of organisms. Science 1999, 284, 1677–1679. [Google Scholar] [CrossRef]
- West, G.; Woodruff, W.; Brown, J. Allometric scaling of metabolic rate from molecules and mitochondria to cells and mammals. Proc. Natl. Acad. Sci. USA 2002, 99, 2473–2478. [Google Scholar] [CrossRef]
- Banavar, J.; Maritan, A.; Rinaldo, A. Size and form in efficient transportation networks. Nature 1999, 399, 130–132. [Google Scholar] [CrossRef]
- Banavar, J.; Moses, M.; Brown, J.; Damuth, J.; Rinaldo, A.; Sibly, R.; Maritan, A. A general basis for quarter-power scaling in animals. Proc. Natl. Acad. Sci. USA 2010, 107, 15816–15820. [Google Scholar] [CrossRef]
- Banavar, J.; Cooke, T.; Rinaldo, A.; Maritan, A. Form, function, and evolution of living organisms. Proc. Natl. Acad. Sci. USA 2014, 111, 3332–3337. [Google Scholar] [CrossRef]
- Reich, P.; Tjoelker, M.; Machado, J.; Oleksyn, J. Universal scaling of respiratory metabolism, size and nitrogen in plants. Nature 2006, 439, 457–461. [Google Scholar] [CrossRef] [PubMed]
- Mori, S.; Yamaji, K.; Ishida, A.; Prokushkin, S.; Masyagina, O.; Hagihara, A.; Hoque, A.; Suwa, R.; Osawa, A.; Nishizono, T. Others Mixed-power scaling of whole-plant respiration from seedlings to giant trees. Proc. Natl. Acad. Sci. USA 2010, 107, 1447–1451. [Google Scholar] [CrossRef] [PubMed]
- Ballesteros, F.; Martinez, V.; Luque, B.; Lacasa, L.; Valor, E.; Moya, A. On the thermodynamic origin of metabolic scaling. Sci. Rep. 2018, 8, 1448. [Google Scholar] [CrossRef] [PubMed]
- Rameaux, S. Rapport sur un mémoire adressé a l’Académie royale de Médicine. Bull. Acad. Nat. Roy. 1838, 3, 1094–1100. [Google Scholar]
- Velarde, C.; Robledo, A. Rank distributions: Frequency vs. magnitude. PLoS ONE 2017, 12, e0186015. [Google Scholar] [CrossRef] [PubMed]
- Velarde, C.; Robledo, A. Dynamical analogues of rank distributions. PLoS ONE 2019, 14, e0211226. [Google Scholar] [CrossRef] [PubMed]
- Velarde, C.; Robledo, A. Number theory, borderline dimension and extensive entropy in distributions of ranked data. PLoS ONE 2022, 17, e0279448. [Google Scholar] [CrossRef]
- Saddle-Node Bifurcation (Redirected from Tangent Bifurcation). Wikipedia. Available online: https://en.wikipedia.org/wiki/Saddle-node_bifurcation (accessed on 24 December 2023).
- Schuster, H.; Just, W. Deterministic Chaos: An Introduction; John Wiley & Sons, Inc.: New York, NY, USA; London, UK, 2006. [Google Scholar]
- Hu, B.; Rudnick, J. Exact solutions to the Feigenbaum renormalization-group equations for intermittency. Phys. Rev. Lett. 1982, 48, 1645. [Google Scholar] [CrossRef]
- Baldovin, F.; Robledo, A. Sensitivity to initial conditions at bifurcations in one-dimensional nonlinear maps: Rigorous nonextensive solutions. Europhys. Lett. 2002, 60, 518. [Google Scholar] [CrossRef]
- Routes to Chaos. Encyclopedia of Mathematics. Wikipedia. Available online: https://encyclopediaofmath.org/wiki/Routes_to_chaos (accessed on 24 December 2023).
- Robledo, A.; Velarde, C. How, why and when Tsallis statistical mechanics provides precise descriptions of natural phenomena. Entropy 2022, 24, 1761. [Google Scholar] [CrossRef]
- Tsallis, C. Possible generalization of Boltzmann-Gibbs statistics. J. Stat. Phys. 1988, 52, 479–487. [Google Scholar] [CrossRef]
- Tsallis, C. Introduction to Nonextensive Statistical Mechanics: Approaching a Complex World; Springer: Berlin/Heidelberg, Germany, 2009. [Google Scholar]
- Pietronero, L.; Tosatti, E.; Tosatti, V.; Vespignani, A. Explaining the uneven distribution of numbers in nature: The laws of Benford and Zipf. Phys. Stat. Mech. Appl. 2001, 293, 297–304. [Google Scholar] [CrossRef]
- Quantile Function. Wikipedia. Available online: https://en.wikipedia.org/wiki/Quantile_function (accessed on 24 December 2023).
- Zipf’s Law. Wikipedia. Available online: https://en.wikipedia.org/wiki/Zipf%27s_law (accessed on 24 December 2023).
- Barabási, A.; Albert, R. Emergence of scaling in random networks. Science 1999, 286, 509–512. [Google Scholar] [CrossRef] [PubMed]
- Barabási, A.-L. The Barabási-Albert Model. In Network Science; Cambridge University Press: Cambridge, UK, 2016; Available online: http://barabasi.com/networksciencebook/ (accessed on 24 December 2023).
- Landau Theory. Wikipedia. Available online: https://en.wikipedia.org/wiki/Landau_theory (accessed on 24 December 2023).
- Robledo, A.; Camacho-Vidales, L.J. A zodiac of studies on complex systems. Supl. Rev. Mex. Física 2020, 1, 32–53. [Google Scholar] [CrossRef]
- Kurosawa, Y.; Mori, S.; Wang, M.; Pedro Ferrio, J.; Nishizono, T.; Yamaji, K.; Koyama, K.; Haruma, T.; Doyama, K. Ontogenetic changes in root and shoot respiration, fresh mass and surface area of Fagus crenata. Ann. Bot. 2023, 131, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Hack’s Law. Wikipedia. Available online: https://en.wikipedia.org/wiki/Hack%27s_law (accessed on 24 December 2023).
- Rinaldo, A.; Rigon, R.; Banavar, J.; Maritan, A.; Rodriguez-Iturbe, I. Evolution and selection of river networks: Statics, dynamics, and complexity. Proc. Natl. Acad. Sci. USA 2014, 111, 2417–2424. [Google Scholar] [CrossRef]
- McNab, B. An analysis of the factors that influence the level and scaling of mammalian BMR. Comp. Biochem. Physiol. Part Mol. Integr. Physiol. 2008, 151, 5–28. [Google Scholar] [CrossRef]
- McNab, B. Ecological factors affect the level and scaling of avian BMR. Comp. Biochem. Physiol. Part Mol. Integr. Physiol. 2009, 152, 22–45. [Google Scholar] [CrossRef]
- Lyapunov Function. Wikipedia. Available online: https://en.wikipedia.org/wiki/Lyapunov_function (accessed on 24 December 2023).
- Ezekiel, M. The cobweb theorem. Q. J. Econ. 1938, 52, 255–280. [Google Scholar] [CrossRef]
- Waugh, F. Cobweb models. Am. J. Agric. Econ. 1964, 46, 732–750. [Google Scholar] [CrossRef]
- Winfree, A. Biological rhythms and the behavior of populations of coupled oscillators. J. Theor. Biol. 1967, 16, 15–42. [Google Scholar] [CrossRef] [PubMed]
- Feigenbaum, M. Quantitative universality for a class of nonlinear transformations. J. Stat. Phys. 1978, 19, 25–52. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Camacho-Vidales, L.J.; Robledo, A. A Nonlinear Dynamical View of Kleiber’s Law on the Metabolism of Plants and Animals. Entropy 2024, 26, 32. https://doi.org/10.3390/e26010032
Camacho-Vidales LJ, Robledo A. A Nonlinear Dynamical View of Kleiber’s Law on the Metabolism of Plants and Animals. Entropy. 2024; 26(1):32. https://doi.org/10.3390/e26010032
Chicago/Turabian StyleCamacho-Vidales, Luis Jovanny, and Alberto Robledo. 2024. "A Nonlinear Dynamical View of Kleiber’s Law on the Metabolism of Plants and Animals" Entropy 26, no. 1: 32. https://doi.org/10.3390/e26010032
APA StyleCamacho-Vidales, L. J., & Robledo, A. (2024). A Nonlinear Dynamical View of Kleiber’s Law on the Metabolism of Plants and Animals. Entropy, 26(1), 32. https://doi.org/10.3390/e26010032