Effects of Anonaine on Dopamine Biosynthesis and L-DOPA-Induced Cytotoxicity in PC12 Cells

Abstract
:Introduction

Results and Discussion
Intracellular dopamine content

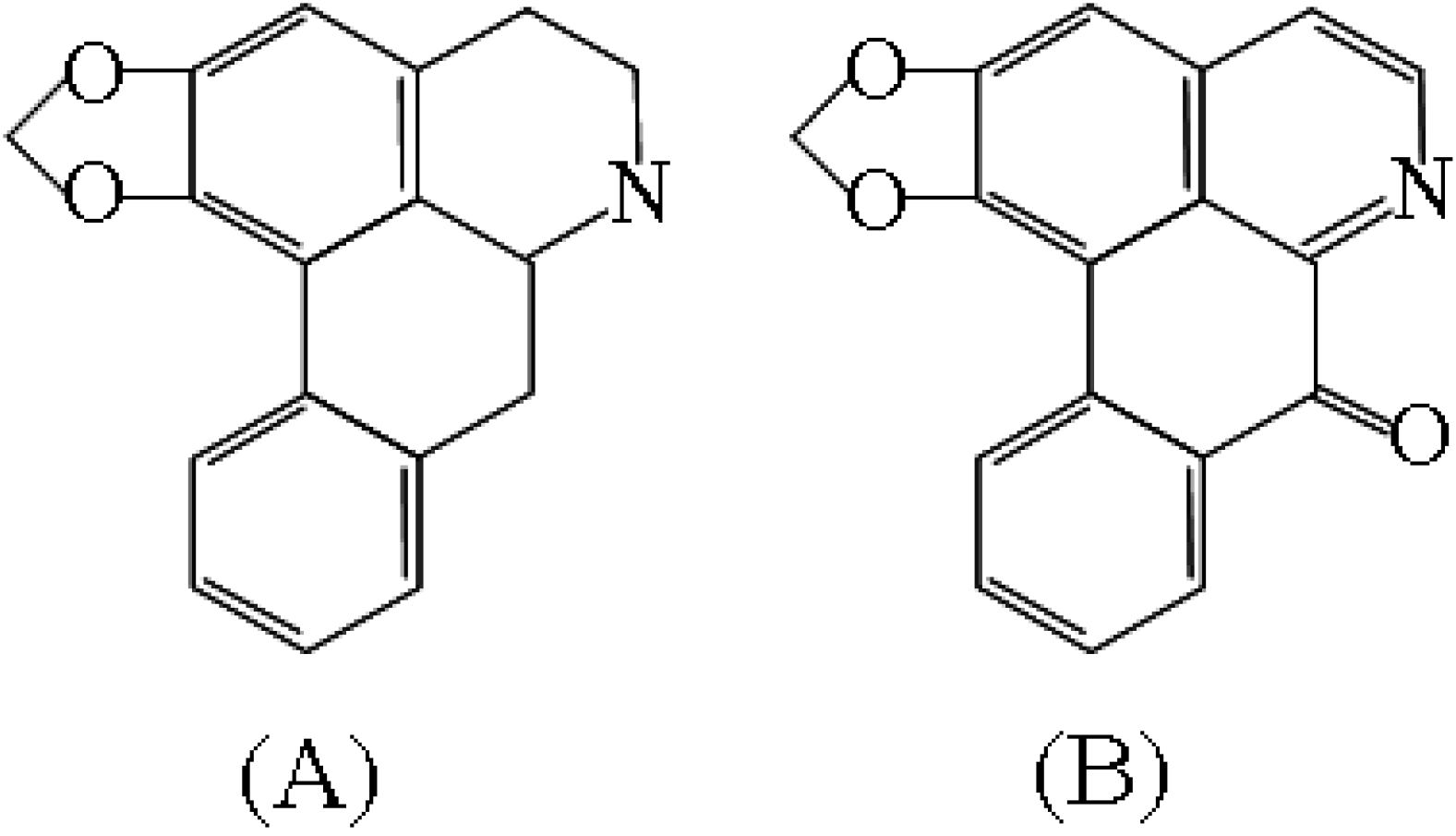{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dopamine content (nmol/mg protein) | (% of control) IC50 value | |
|---|---|---|
| Control | 3.70 ± 0.23 | (100) |
| Anonaine | 0.05 μM | |
| 0.01 μM | 3.05 ± 0.26 | (82.4) |
| 0.03 μM | 2.55± 0.17 | (68.9)* |
| 0.06 μM | 1.56 ± 0.19 | (42.2)** |
| 0.08 μM | 0.94 ± 0.15 | (25.4)*** |

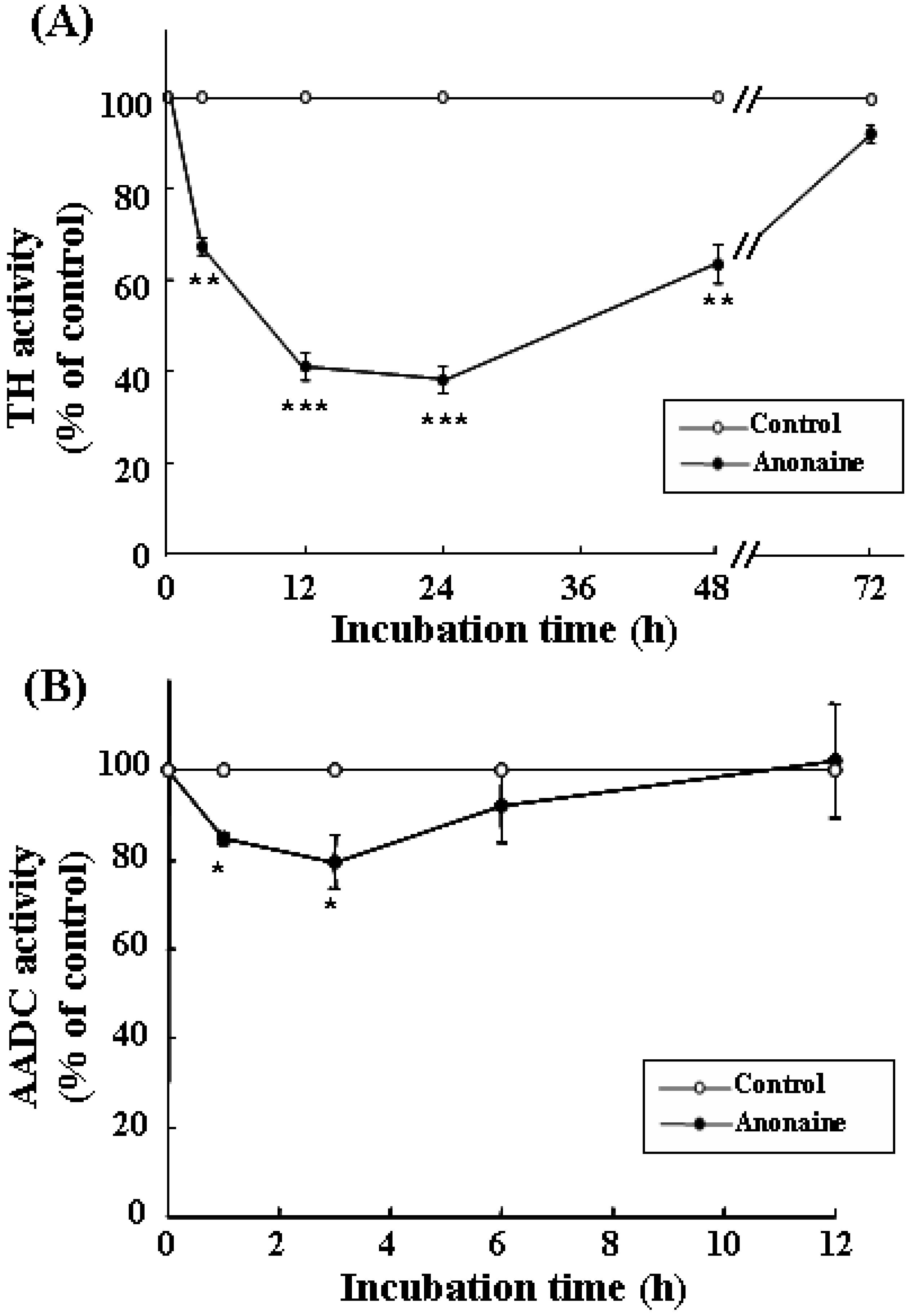
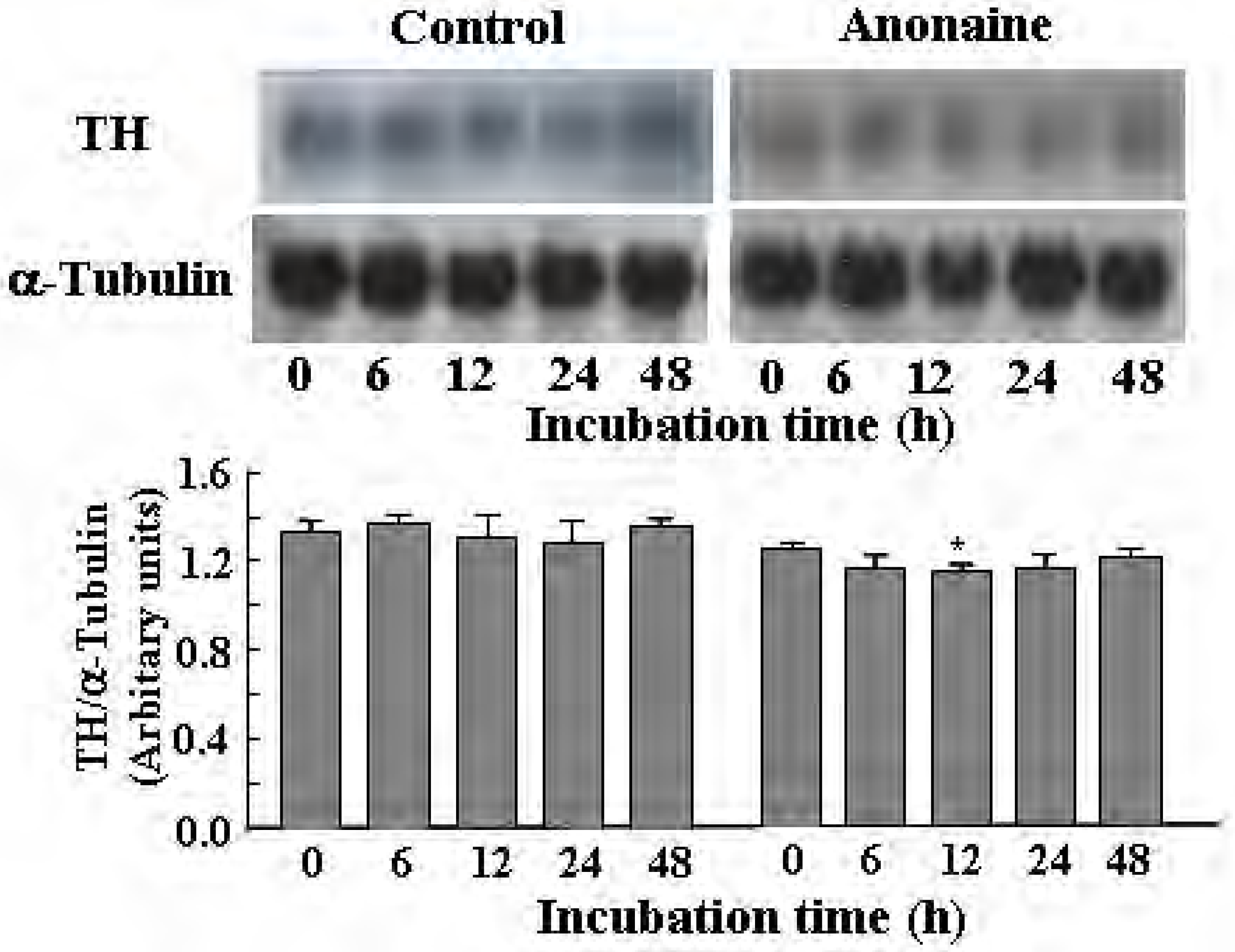
TH and AADC activities

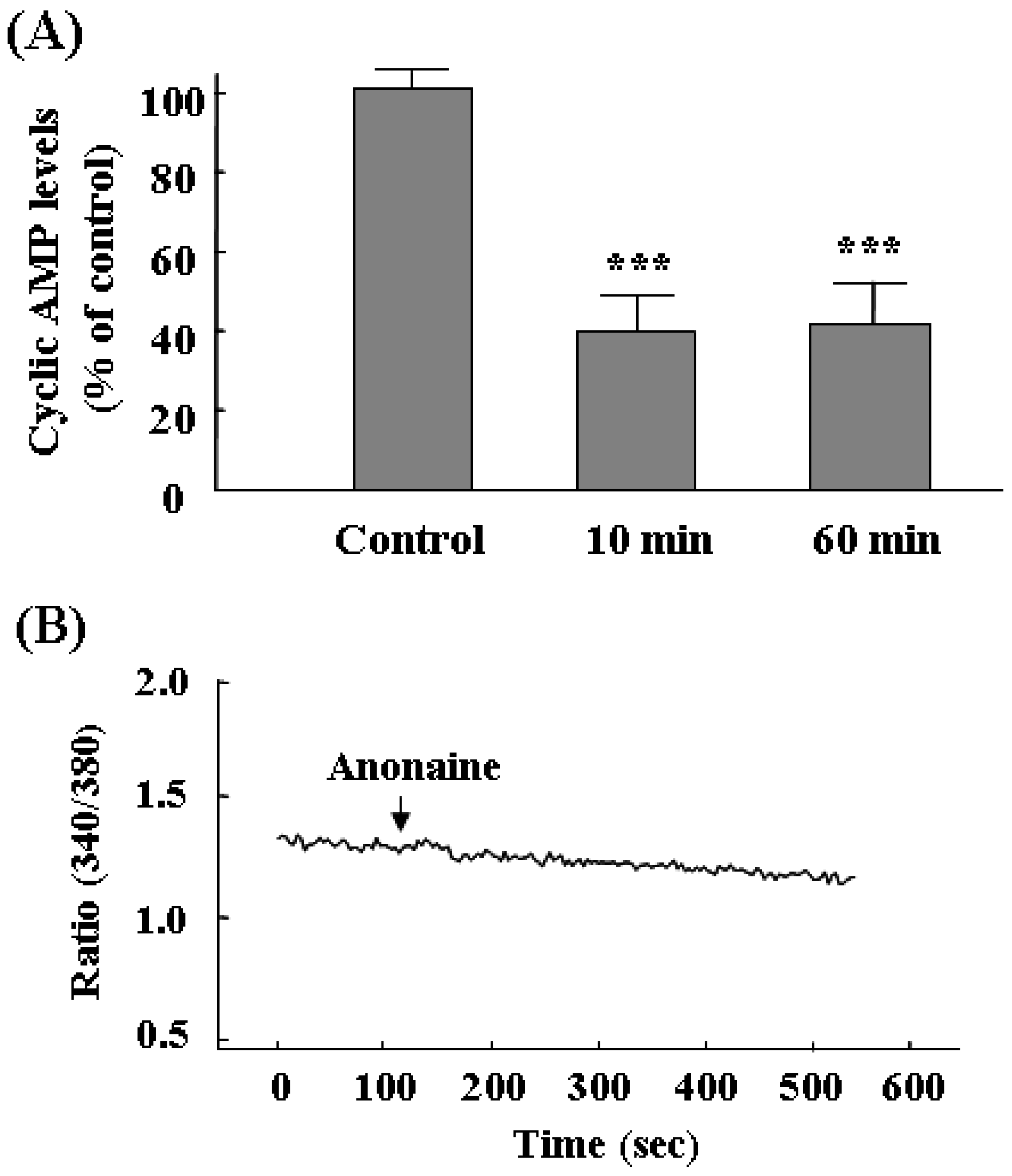
TH mRNA and cyclic AMP levels, and Ca2+ concentration

l-DOPA-induced dopamine biosynthesis and cytotoxicity


Conclusions
Experimental
Materials
Cell culture
Determination of dopamine content
Assay for TH and AADC activities
RNA extraction and Northern blotting
Measurement of cyclic AMP levels
Measurement of intracellular Ca2+ concentration
Measurement of cell viability: MTT assay
Statistical analysis
Acknowledgements
References
- Martinez, L.A.; Rios, J.L.; Paya, M.; Alcaraz, M.J. Inhibition of nonenzymic lipid peroxidation by benzylisoquinoline alkaloids. Free Rad. Biol. Med. 1992, 12, 287–292. [Google Scholar] [CrossRef]
- Paulo, M. de Q.; Barbosa-Fiho, J.M.; Lima, E.O.; Maia, R.F.; Barbosa, R. de C.; Kaplan, M.A. Antimicrobial activity of benzylisoquinoline alkaloids from annona salzmanii D.C. J. Ethnopharmacol. 1992, 36, 39–41. [Google Scholar] [CrossRef]
- Protais, P.; Arbaoui, J.; Bakkali, E.H.; Bermejo, A.; Cortes, D. Effects of various isoquinoline alkaloids on in vitro 3H-dopamine uptake by rat striatal synaptosomes. J. Nat. Prod. 1995, 58, 1475–1484. [Google Scholar] [CrossRef]
- Kim, K.S.; Park, D.H.; Wessel, T.C.; Song, B.; Wagner, J.A.; Joh, T.H. A dual role for the cAMP-dependent protein kinase in tyrosine hydroxylase gene expression. Proc. Natl. Acad. Sci. USA 1993, 90, 3471–3475. [Google Scholar]
- Dunkley, P.R.; Cote, A.; Harrison, S.M.; Herd, L.; Hall, A.; Powis, D.A. Tyrosine hydroxylase phosphorylation in bovine adrenal chromaffin cells. Biochem. Pharmacol. 1996, 51, 239–245. [Google Scholar] [CrossRef]
- Young, E.A.; Duchemin, A.-M.; Neff, N.H.; Hadjiconstantinou, M. Parallel modulation of striatal dopamine synthetic enzymes by second messenger pathways. Eur. J. Pharmacol. 1998, 357, 15–23. [Google Scholar] [CrossRef]
- Marsden, C.D. Parkinson’s disease. J. Neurol. Neurosurg. Ps. 1994, 57, 672–681. [Google Scholar] [CrossRef]
- Ziv, I.; Zilkha-Falb, R.; Offen, D.; Shirvan, A.; Barzilai, A.; Melamed, E. Levodopa induces apoptosis in cultured neuronal cells: a possible accelerator of nigrostriatal degeneration in Parkinson's disease? Mov. Disord. 1997, 12, 17–23. [Google Scholar] [CrossRef]
- Basma, A.N.; Morris, E.J.; Nicklas, W.J.; Geller, H.M. L-DOPA cytotoxicity to PC12 cells in culture is via its antoxidation. J. Neurochem. 1995, 64, 825–832. [Google Scholar]
- Shin, J.S.; Kim, K.T.; Lee, M.K. Inhibitory effects of bulbocapnine on dopamine biosynthesis in PC12 cells. Neurosci. Lett. 1998, 244, 161–164. [Google Scholar] [CrossRef]
- Jin, C.H.; Lee, J.J.; Yang, Y.J.; Kim, Y.M.; Kim, Y.K.; Ryu, S.Y.; Lee, M.K. Liriodenine inhibits dopamine biosynthesis and L-DOPA-induced dopamine content in PC12 cells. Arch. Pharm. Res. 2007, 30, 984–990. [Google Scholar] [CrossRef]
- Cassels, B.K.; Asencio, M.; Conget, P.; Speisky, H.; Videla, L.A.; Lissi, E.A. Structure-antioxidative activity relationships in benzylisoquinoline alkaloids. Pharmacol. Res. 1995, 31, 103–107. [Google Scholar]
- Asencio, M.; Delaquerriere, B.; Cassels, B.K.; Speisky, H.; Comoy, E.; Protais, P. Biochemical and behavioral effects of boldine and glaucine on dopamine systems. Pharmacol. Biochem. Behav. 1999, 62, 7–13. [Google Scholar] [CrossRef]
- Loghin, F.; Chagraoui, A.; Asencio, M.; Comoy, E.; Speisky, H.; Cassels, B.; Protais, P. Effects of some antioxidative aporphine derivatives on striatal dopaminergic transmission and on MPTP-induced striatal dopamine depletion in B6CBA mice. Pharmacol. Sci. 2003, 18, 133–140. [Google Scholar]
- Tischler, A.S.; Perlman, R.L.; Morse, G.M.; Sheard, B.E. Glucocorticoids increase catecholamine synthesis and storage in PC12 cells pheochromocytoma cell culture. J. Neurochem. 1983, 40, 364–370. [Google Scholar] [CrossRef]
- Byrd, J.C.; Hadjiconstantinou, M.; Cavalla, D. Epinephrine synthesis in the PC12 pheochromo-cytoma cell line. Eur. J. Pharmacol. 1986, 127, 139–142. [Google Scholar] [CrossRef]
- Lee, J.J.; Kim, Y.M.; Yin, S.Y.; Park, H.D.; Kang, M.H.; Hong, J.T.; Lee, M.K. Aggravation of l-DOPA-induced neurotoxicity by tetrahydropapaveroline in PC12 cells. Biochem. Pharmacol. 2003, 66, 1787–1795. [Google Scholar] [CrossRef]
- Shin, J.S.; Yun-Choi, H.S.; Kim, E.I.; Lee, M.K. Inhibitory effects of higenamine on dopamine content in PC12 cells. Planta Med. 1999, 65, 452–455. [Google Scholar] [CrossRef]
- Shin, J.S.; Kim, E.I.; Kai, M.; Lee, M.K. Inhibition of dopamine biosynthesis by protoberberine alkaloids in PC12 cells. Neurochem. Res. 2000, 25, 363–368. [Google Scholar] [CrossRef]
- Shin, J.S.; Lee, J.J.; Kim, Y.; Lee, C.K.; Yun, Y.P.; Lee, M.K. Inhibitory effects of ethaverine, a homologue of papaverine, on dopamine content in PC12 cells. Biol. Pharm. Bull. 2001, 24, 103–105. [Google Scholar] [CrossRef]
- Yin, S.Y.; Kim, Y.M.; Lee, J.J.; Jin, C.M.; Yang, Y.J.; Ma, J.J.; Kang, M.H.; Kai, M.; Lee, M.K. Enantio-selective inhibition of (1R,9S)- and (1S,9R)-β-hydrastines on dopamine biosynthesis in PC12 cells. Neuropharmacol. 2004, 47, 1045–1052. [Google Scholar] [CrossRef]
- Kim, Y.M.; Kim, M.N.; Lee, J.J.; Lee, M.K. Inhibition of dopamine biosynthesis by tetrahydropapaveroline. Neurosci. Lett. 2005, 386, 1–4. [Google Scholar] [CrossRef]
- Lee, M.K.; Zhang, Y.H. Inhibition of tyrosine hydroxylase by berberine. Med. Sci. Res. 1996, 24, 561–562. [Google Scholar]
- Lee, M.K.; Zhang, Y.H.; Shin, J.S.; Lee, S.S. Inhibition of tyrosine hydroxylase by hydrastine. Med. Sci. Res. 1997, 25, 619–620. [Google Scholar]
- Zhang, Y.H.; Shin, J.S.; Lee, S.S.; Kim, S.H.; Lee, M.K. Inhibition of tyrosine hydroxylase by bulbocapnine. Planta Med. 1997, 63, 362–363. [Google Scholar] [CrossRef]
- Cho, S.; Neff, N.H.; Hadjiconstantinou, M. Regulation of tyrosine hydroxylase and aromatic L-amino acid decarboxylase by dopaminergic drugs. Eur. J. Pharmacol. 1997, 323, 149–157. [Google Scholar] [CrossRef]
- Yin, S.Y.; Lee, J.J.; Kim, Y.M.; Jin, C.M.; Yang, Y.J.; Kang, M.H.; Kai, M.; Lee, M.K. Effects of (1R,9S)-β-hydrastine on L-DOPA-induced cytotoxicity in PC12 cells. Eur. J. Pharmacol. 2004, 488, 71–77. [Google Scholar] [CrossRef]
- Bannach, R.; Valenzuela, A.; Cassels, B.K.; Nunez-Vergara, L.J.; Speisky, H. Cytoprotective and antioxidant effects of boldine on tert-butyl hydroperoxide-induced damage to isolated hepatocytes. Cell. Biol. Toxicol. 1996, 12, 89–100. [Google Scholar] [CrossRef]
- Youn, Y.C.; Kwon, O.S.; Han, E.S.; Song, J.H.; Shin, Y.K.; Lee, C.S. Protective effect of boldine on dopamine-induced membrane permeability transition in brain mitochondria and viability loss in PC12 cells. Biochem. Pharmacol. 2002, 63, 495–505. [Google Scholar] [CrossRef]
- Woo, S.H.; Sun, N.J.; Cassady, J.M.; Snapka, R.M. Topoisomerase II inhibition by aporphine alkaloids. Biochem. Pharmacol. 1999, 57, 1141–1145. [Google Scholar]
- Kim, Y.K.; Ryu, S.Y. Cytotoxic components from stem bark of Magnolia ovobata. Planta Med. 1999, 65, 291–292. [Google Scholar] [CrossRef]
- Nagatsu, T.; Oka, K.; Kato, K. Highly sensitive assay for tyrosine hydroxylase activity by high-performance liquid chromatography. J. Chromatogr. 1979, 163, 247–252. [Google Scholar] [CrossRef]
- Lee, M.K.; Nohta, H.; Ohkura, Y. Occurrence of aromatic L-amino acid decarboxylase in human plasma and its assay by high-performance liquid chromatography with fluorescence detection. J. Chromatogr. 1986, 378, 329–336. [Google Scholar] [CrossRef]
- Chomczynski, P.; Sacchi, N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-cholroform extraction. Anal. Biochem. 1987, 162, 156–159. [Google Scholar]
- Mosman, T. Rapid colorimetric assay for cellular growth and survival: application to proliferation and cytotoxic assays. J. Immunol. Meth. 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosenbrough, N.L.; Farr, A.L.; Randall, R.L. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar]
- Sample availibility: Contact the authors.
© 2008 by MDPI (http://www.mdpi.org). Reproduction is permitted for noncommercial purposes.
Share and Cite
Lee, J.J.; Jin, C.M.; Kim, Y.K.; Ryu, S.Y.; Lim, S.C.; Lee, M.K. Effects of Anonaine on Dopamine Biosynthesis and L-DOPA-Induced Cytotoxicity in PC12 Cells. Molecules 2008, 13, 475-487. https://doi.org/10.3390/molecules13020475
Lee JJ, Jin CM, Kim YK, Ryu SY, Lim SC, Lee MK. Effects of Anonaine on Dopamine Biosynthesis and L-DOPA-Induced Cytotoxicity in PC12 Cells. Molecules. 2008; 13(2):475-487. https://doi.org/10.3390/molecules13020475
Chicago/Turabian StyleLee, Jae Joon, Chun Mei Jin, Young Kyoon Kim, Shi Yong Ryu, Sung Cil Lim, and Myung Koo Lee. 2008. "Effects of Anonaine on Dopamine Biosynthesis and L-DOPA-Induced Cytotoxicity in PC12 Cells" Molecules 13, no. 2: 475-487. https://doi.org/10.3390/molecules13020475
APA StyleLee, J. J., Jin, C. M., Kim, Y. K., Ryu, S. Y., Lim, S. C., & Lee, M. K. (2008). Effects of Anonaine on Dopamine Biosynthesis and L-DOPA-Induced Cytotoxicity in PC12 Cells. Molecules, 13(2), 475-487. https://doi.org/10.3390/molecules13020475