Molecular Docking Studies and Anti-enzymatic Activities of Thai Mango Seed Kernel Extract Against Snake Venoms

Abstract
:1. Introduction
2. Results and Discussion
2.1. In vitro tests for the inhibition of enzymatic activities
2.1.1. PLA2

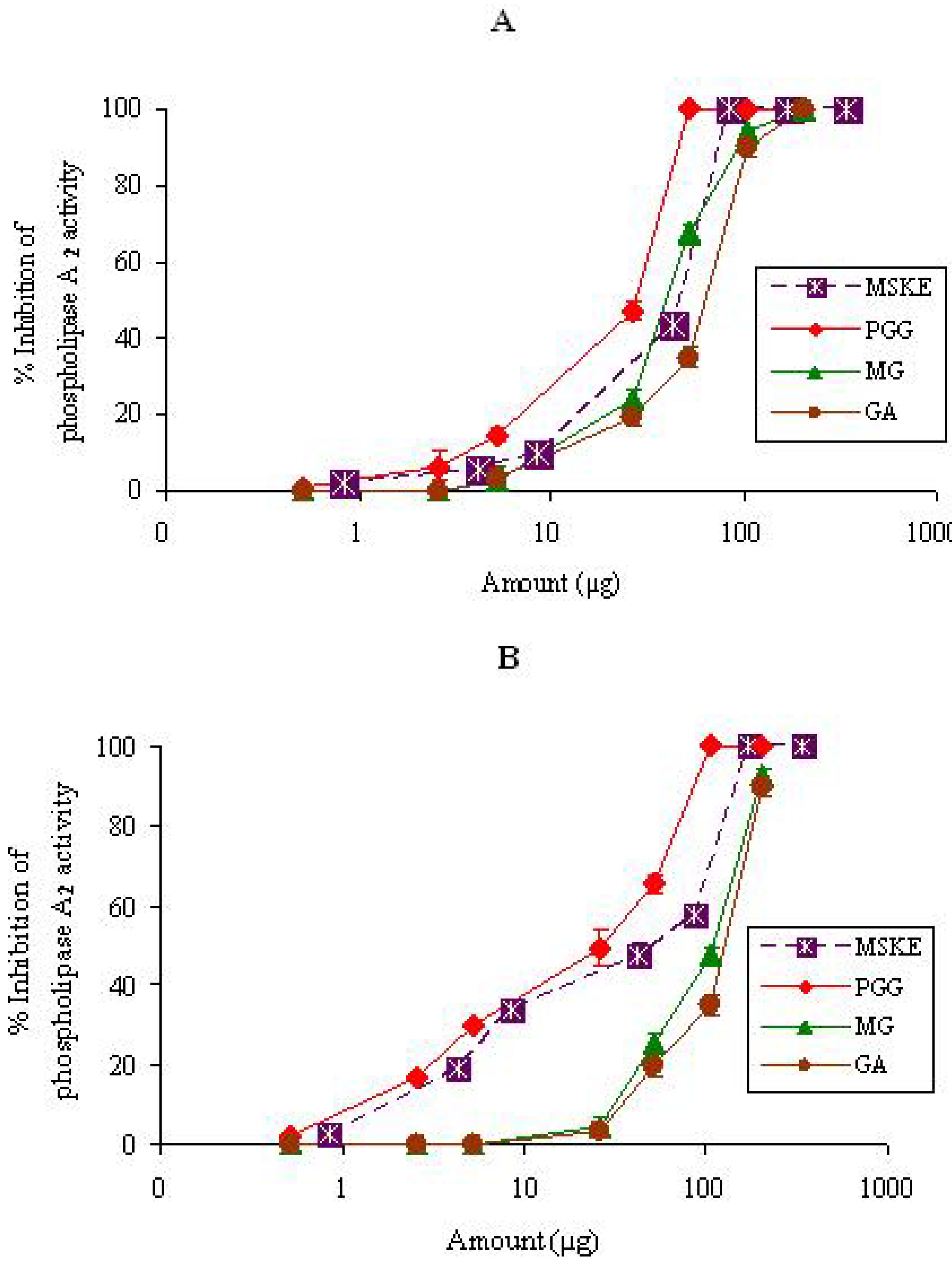{kind=link}
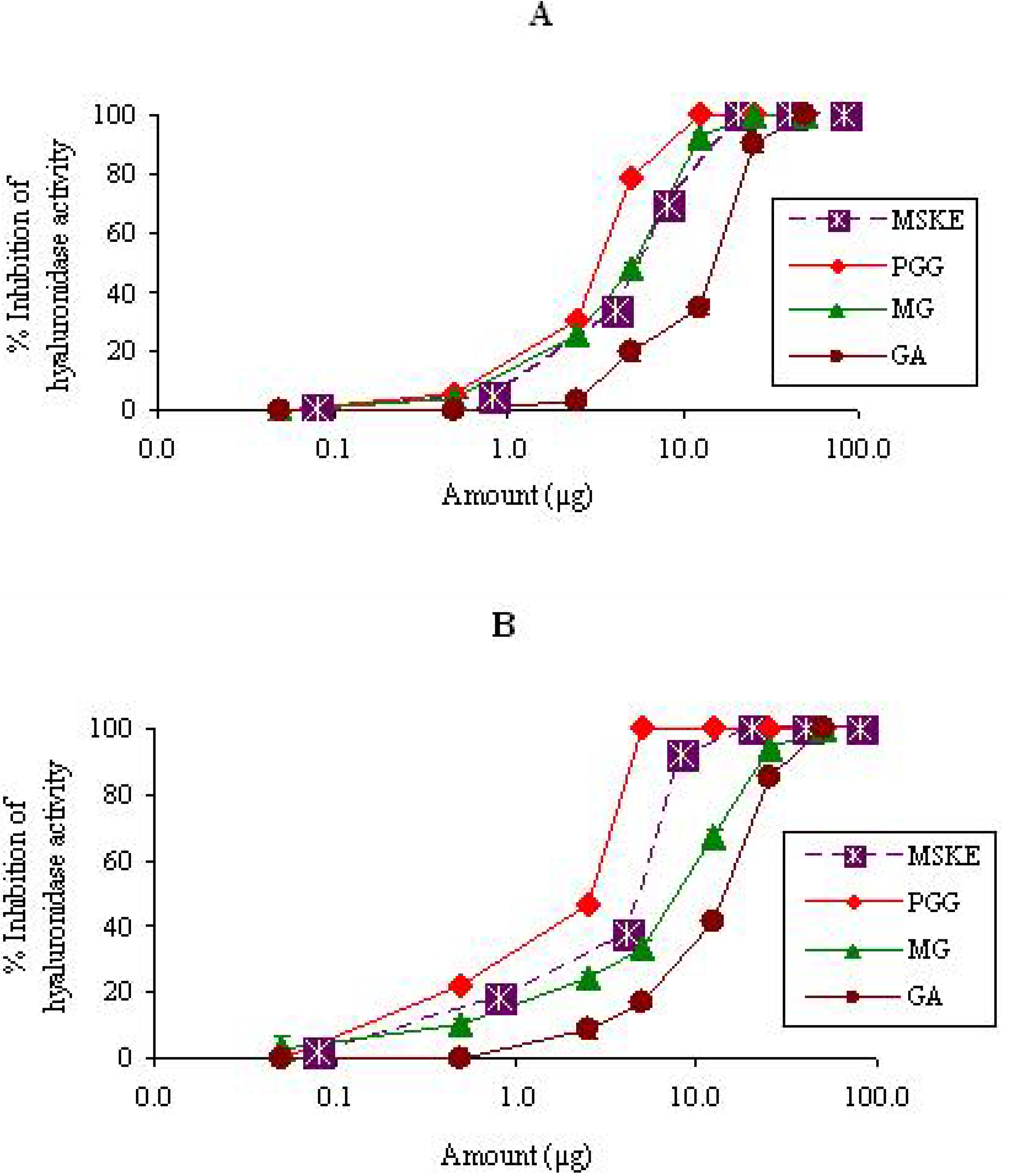{kind=link}
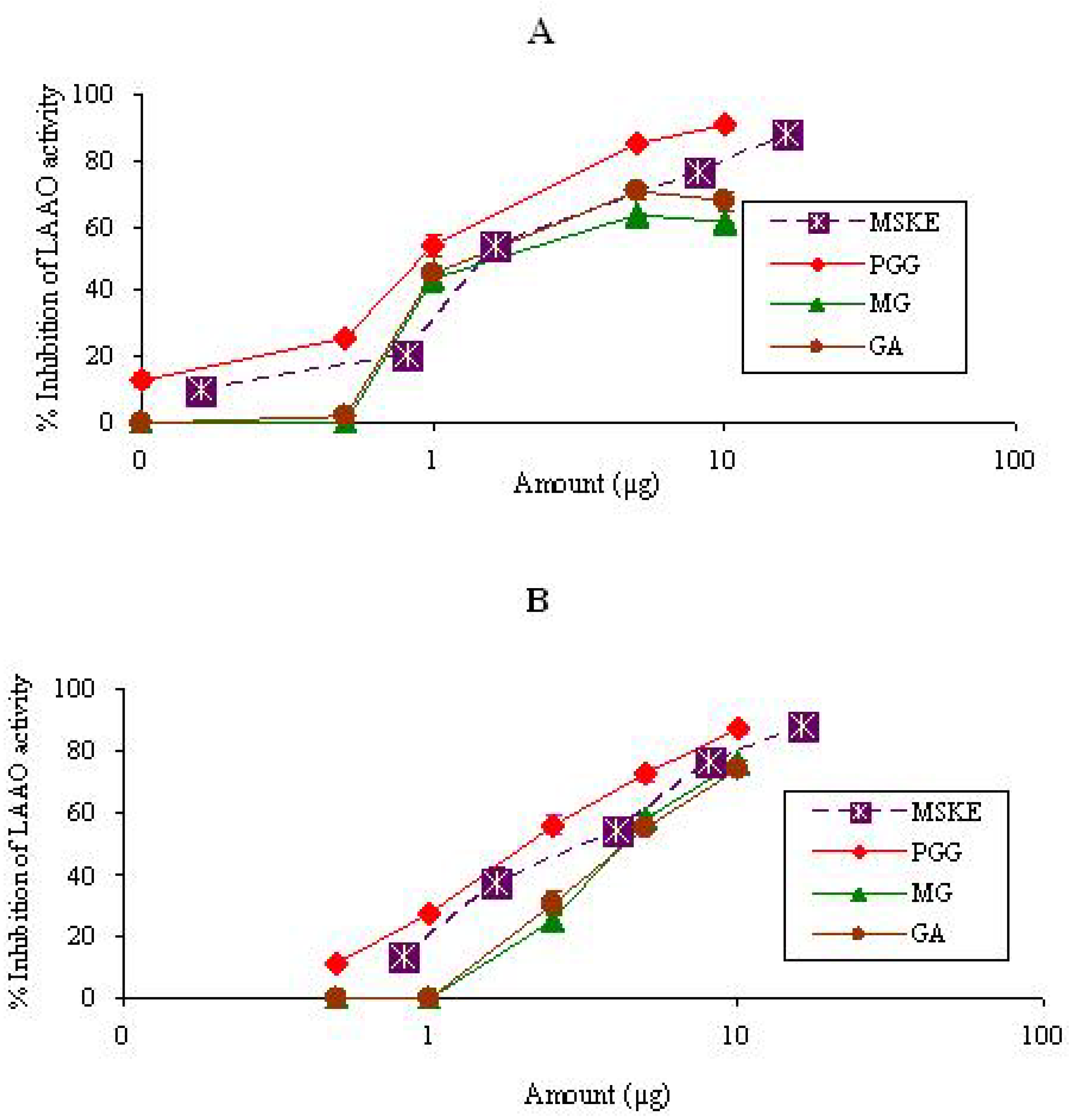{kind=link}
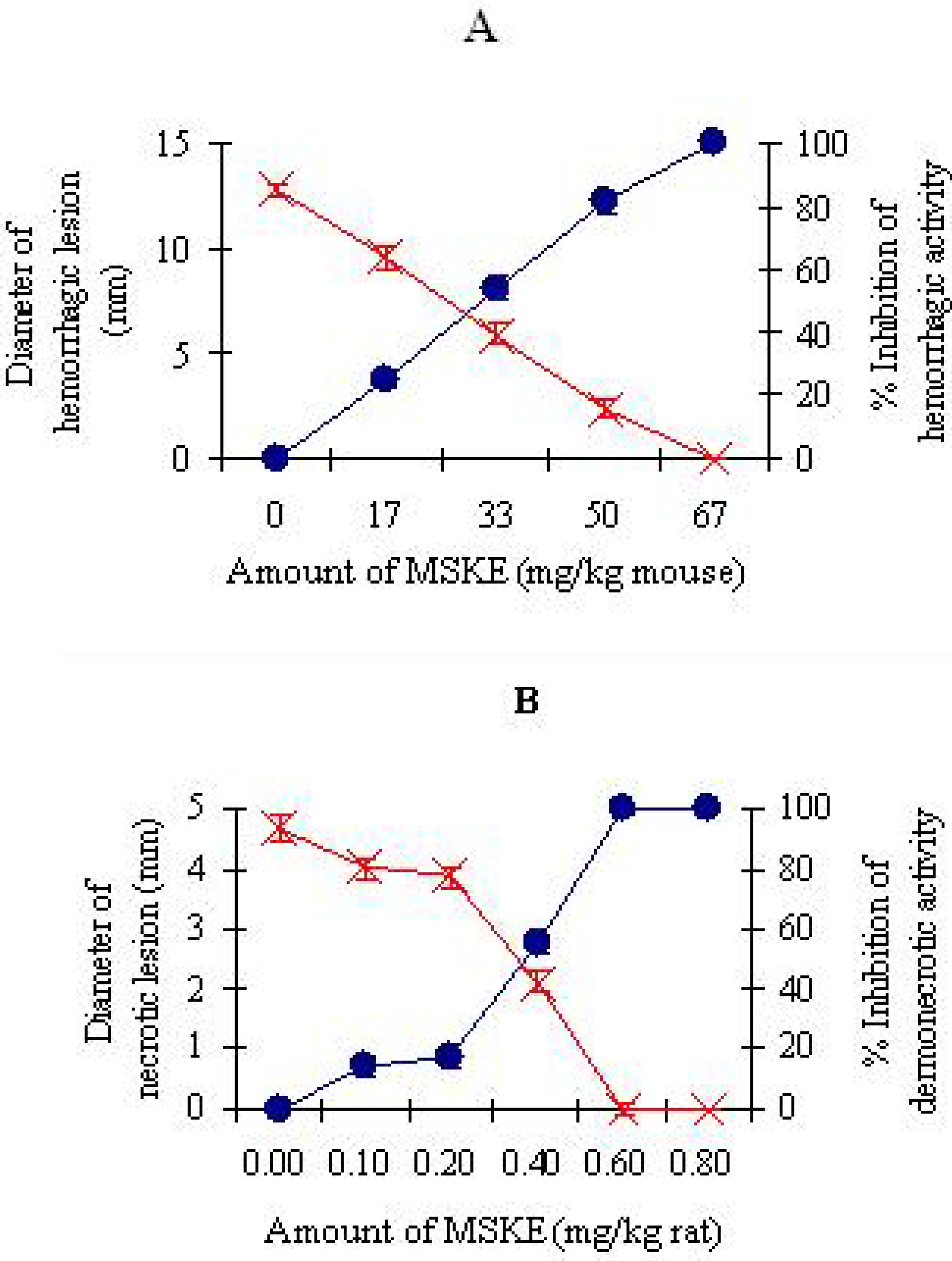{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
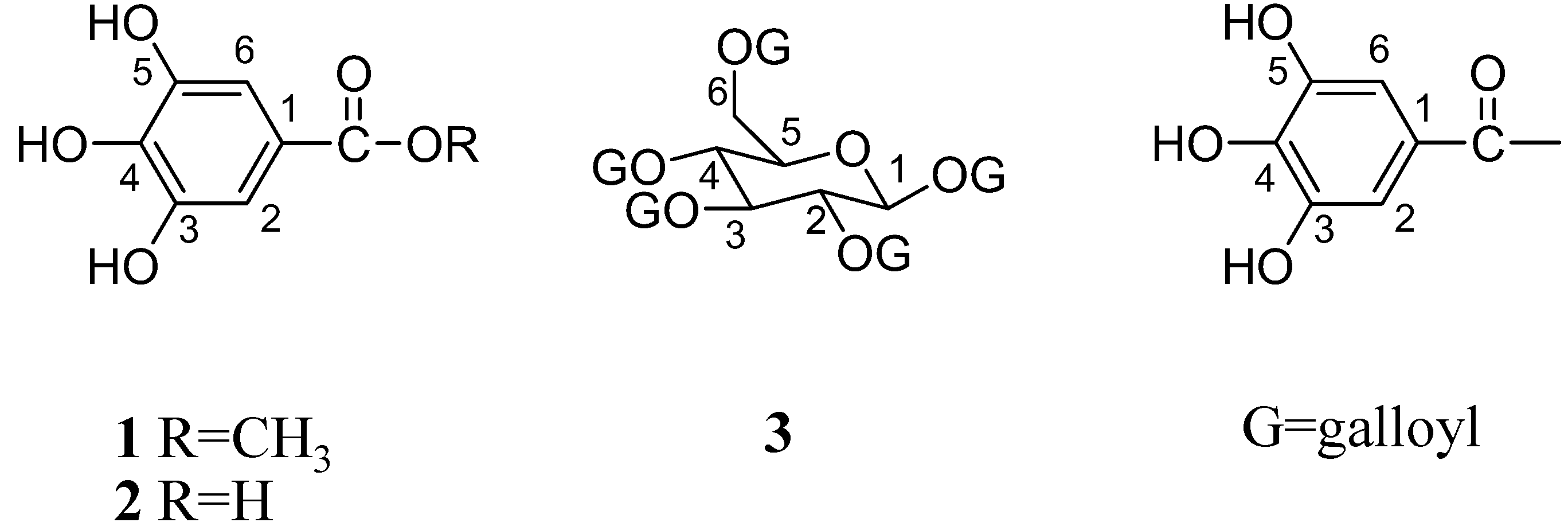
| MSKE and its isolated principles | Contents | PLA2 (ED50, µg) | Hyaluronidase (ED50, µg) | LAAO (ED50, µg) | |||
|---|---|---|---|---|---|---|---|
| (mg/g dry weight) | CR venom | NK venom | CR venom | NK venom | CR venom | NK venom | |
| MSKE | - | 44.4 ± 0.4 | 57.9 ± 0.5 | 6.0 ± 1.9 | 5.0 ± 2.6 | 1.5 ± 3.2 | 4.0 ± 1.7 |
| GA | 4.40 ± 0.05 | 66.0 ± 2.5 | 123.5 ± 2.5 | 16.0 ± 2.5 | 14.8 ± 1.4 | 1.7 ± 3.4 | 4.5 ± 2.8 |
| MG | 6.80 ± 0.02 | 42.9 ± 2.3 | 108.7 ± 2.0 | 5.9 ± 2.4 | 8.6 ± 2.1 | 2.3 ± 2.4 | 4.4 ± 2.0 |
| PGG | 612.80 ± 34.80 | 25.6 ± 1.3 | 30.2 ± 2.4 | 3.4 ± 2.4 | 2.6 ± 1.7 | 0.9 ± 2.4 | 2.2 ± 2.0 |

2.1.2. Hyaluronidase

2.1.3. LAAO

2.2. In vivo tests
2.2.1. Inhibition of hemorrhagic and dermonecrotic activities

2.3. Molecular modeling
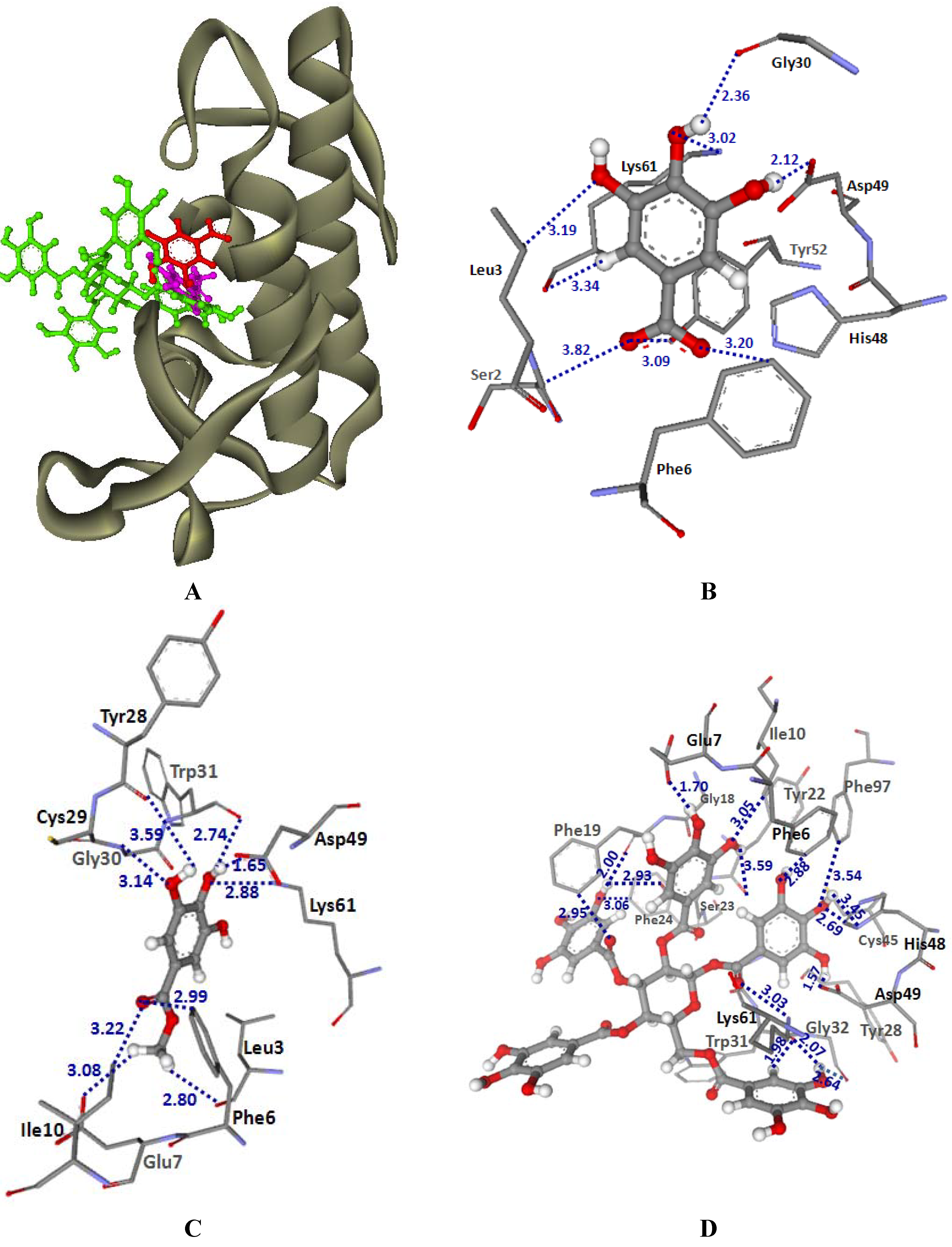
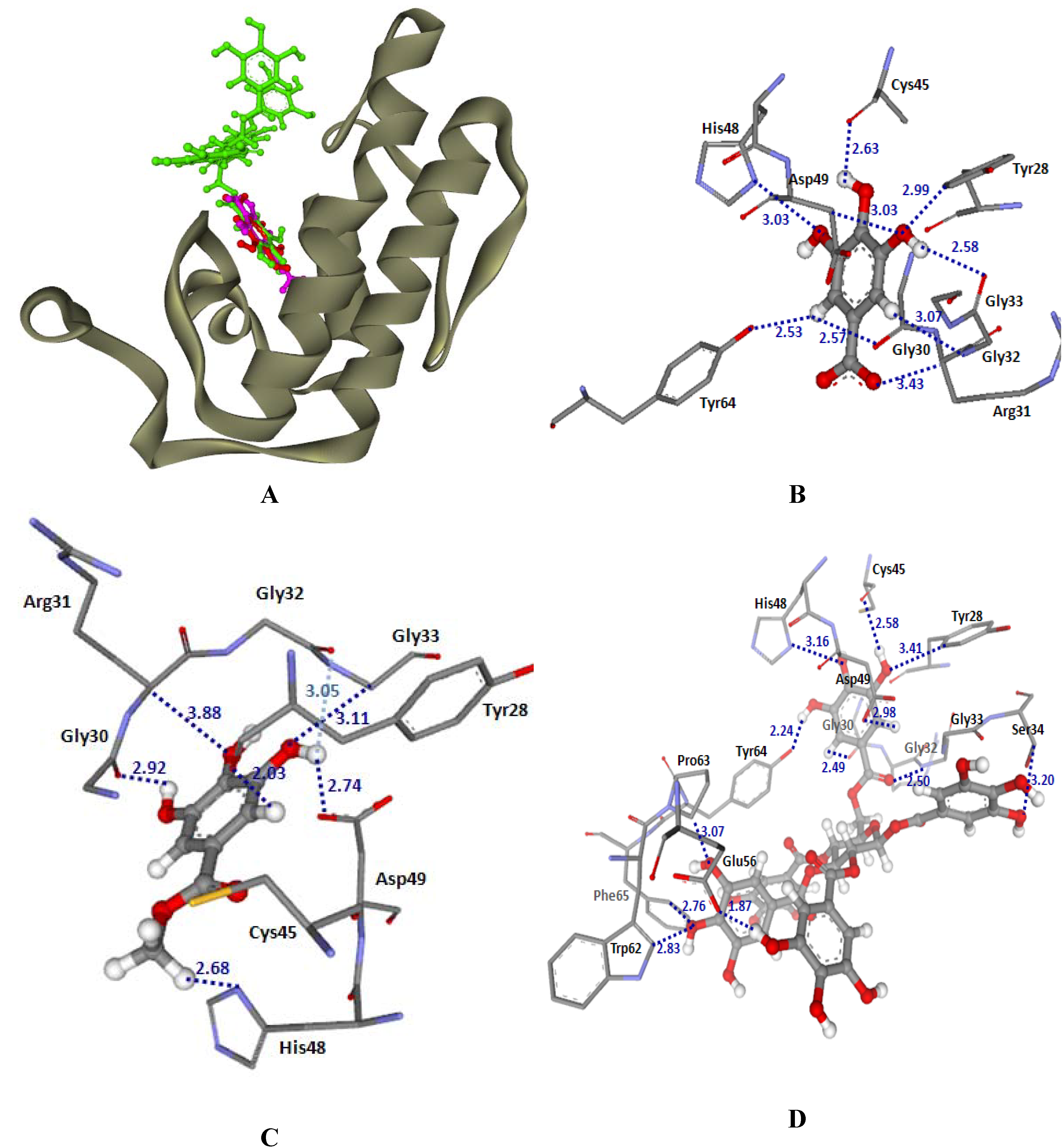
2.3.1. Docking into PLA2-CR and PLA2-NK structures
| GoldScore | |||
|---|---|---|---|
| MSKE constituents | LAAO | PLA2 | |
| CR venom | CR venom | NK venom | |
| GA | 38.95 | 37.55 | 35.52 |
| MG | 39.71 | 37.49 | 34.75 |
| PGG | 69.58 | 77.26 | 64.92 |


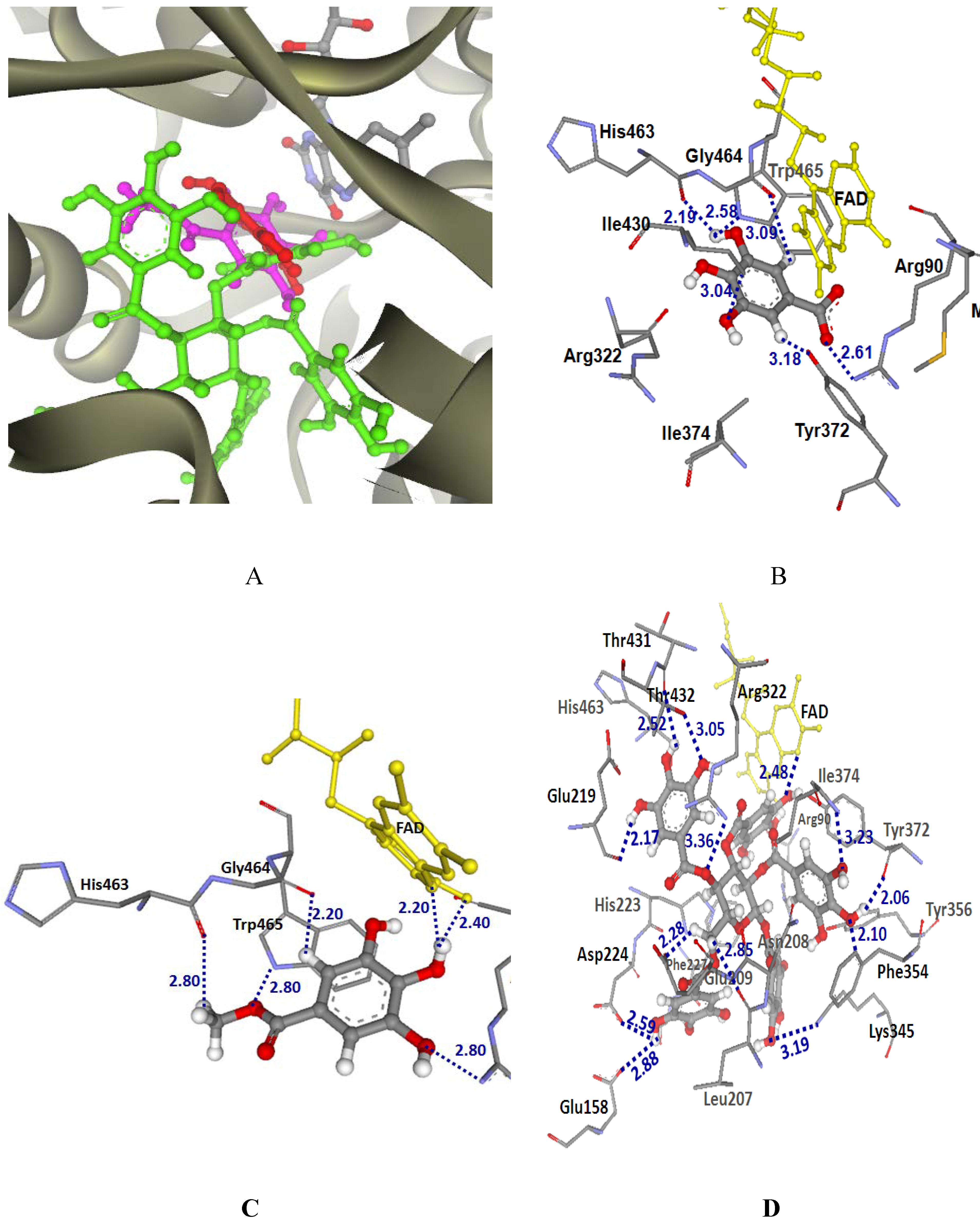
2.3.2. Docking into LAAO-CR structure

3. Conclusions
4. Experimental
4.1. Animals and venoms
4.2. Chemicals
4.3. Plant material and extraction
4.4. Standardization
4.5. In vitro tests
4.5.1. Inhibition of PLA2 activity
4.5.2. Inhibition of hyaluronidase activity
4.5.3. Inhibition of LAAO activity
4.6. In vivo tests
4.6.1. Inhibition of hemorrhagic activity
4.6.2. Inhibition of dermonecrotic activity
4.7. Molecular modeling

4.8. Statistical analysis
Acknowledgements
References
- Viravan, C.; Veeravat, U.; Warrell, M.J.; Theakston, R.D.; Warrell, D.A. ELISA confirmation of acute and past envenoming by the monocellate Thai cobra (Naja kaouthia). Am. J. Trop. Med. Hyg. 1986, 35, 173–181. [Google Scholar]
- Songsumard, S. Snakebite in Thailand. Annual Epidemiological Surveillance Report; Ministry of Public Health: Bangkok, 1995; pp. 324–333. [Google Scholar]
- Markland, F.S. Snake venoms. Drugs 1997, 54, 1–10. [Google Scholar] [CrossRef]
- Aird, S.D. Ophidian envenomation strategies and the role of purines. Toxicon 2002, 40, 335–393. [Google Scholar] [CrossRef]
- Mukherjee, A.K.; Ghosal, S.K.; Maity, C.R. Effect of oral supplementation of vitamin E on the hemolysis and erythrocyte phospholipid-splitting action of cobra and viper venoms. Toxicon 1998, 36, 657–664. [Google Scholar] [CrossRef]
- Kini, R.M.; Evans, H.J. Structure-function relationships of phospholipases. The anticoagulant region of phospholipases A2. J. Biol. Chem. 1987, 262, 14402–14407. [Google Scholar]
- Berg, O.G.; Gelb, M.H.; Tsai, M.D.; Jain, M.K. Interfacial Enzymology: The Secreted Phospholipase A2-Paradigm. Chem. Rev. 2001, 101, 2613–2653. [Google Scholar] [CrossRef]
- Girish, K.S.; Shashidharamurthy, R.; Nagaraju, S.; Gowda, T.V.; Kemparaju, K. Isolation and characterization of hyaluronidase a “Spreading factor” from Indian cobra (Naja naja) venom. Biochimie 2004, 86, 193–202. [Google Scholar] [CrossRef]
- Du, X.Y.; Clemetson, K.J. Snake venom L-amino acid oxidases. Toxicon 2002, 40, 659–665. [Google Scholar] [CrossRef]
- Suhr, S.M.; Kim, D.S. Identification of the snake venom substance that induces apoptosis. Biochem. Biophys. Res. Commun. 1996, 224, 134–139. [Google Scholar] [CrossRef]
- Sakurai, Y.; Takatsuka, H.; Yoshioka, A.; Matsui, T.; Suzuki, M.; Titani, K.; Fujimura, Y. Inhibition of human platelet aggregation by L-amino acid oxidase purified from Naja naja kaouthia venom. Toxicon 2001, 39, 1827–1833. [Google Scholar] [CrossRef]
- Pawelek, P.D.; Cheah, J.; Coulombe, R.; Macheroux, P.; Ghisla, S.; Vrielink, A. The structure of L-amino acid oxidase reveals the substrate trajectory into an enantiomerically conserved active site. EMBO J. 2000, 19, 4204–4215. [Google Scholar] [CrossRef]
- Okonogi, T.; Hattori, Z.; Ogiso, A.; Mitsui, S. Detoxification by persimmon tannin of snake venoms and bacterial toxins. Toxicon 1979, 17, 524–527. [Google Scholar] [CrossRef]
- Abubakar, M.S.; Sule, M.I.; Pateh, U.U.; Abdurahman, E.M.; Haruna, A.K.; Jahun, B.M. In vitro snake venom detoxifying action of the leaf extract of Guiera senegalensis. J. Ethnopharmacol. 2000, 69, 253–257. [Google Scholar] [CrossRef]
- Borges, M.H.; Alves, D.L.F.; Raslan, D.S.; Piló-Veloso, D.; Rodrigues, V.M.; Homsi-Brandeburgo, M.I.; De Lima, M.E. Neutralizing properties of Musa paradisiaca L. (Musaceae) juice on phospholipase A2, myotoxic, hemorrhagic and lethal activities of crotalidae venoms. J. Ethnopharmacol. 98, 21–29.
- Pithayanukul, P.; Ruenraroengsak, P.; Bavovada, R.; Pakmanee, N.; Suttisri, R.; Saen-oon, S. Inhibition of Naja kaouthia venom activities by plant polyphenols. J. Ethnopharmacol. 2005, 97, 527–533. [Google Scholar] [CrossRef]
- Nithitanakool, S.; Pithayanukul, P.; Bavovada, R. Antioxidant and hepatoprotective activities of Thai mango seed kernel extract. Planta Med. (online March 26, 2009).
- Nithitanakool, S.; Pithayanukul, P.; Bavovada, R.; Saparpakorn, P. Molecular docking studies and anti-tyrosinase activity of Thai mango seed kernel extract. Molecules 2009, 14, 257–265. [Google Scholar] [CrossRef]
- Ketelhut, D.F.J.; Mello, M.H.; Veronese, E.L.G.; Esmeraldino, L.E.; Murakami, M.T.; Arni, R.K.; Giglio, J.R.; Cintra, A.C.O.; Sampaio, S.V. Isolation, characterization and biological activity of acidic phospholipase A2 isoforms from Bothrops jararacussu snake venom. Biochimie 2003, 85, 983–991. [Google Scholar] [CrossRef]
- Soares, A.M.; Ticli, F.K.; Marcussi, S.; Lourenco, M.V.; Januario, A.H.; Sampaio, S.V.; Giglio, J.R.; Lomonte, B.; Pereira, P.S. Medicinal plants with inhibitory properties against snake venoms. Curr. Med. Chem. 2005, 12, 2625–2641. [Google Scholar] [CrossRef]
- Ushanandini, S.; Nagaraju, S.; Kumar, K.H.; Vedavathi, M.; Machiah, D.K.; Kemparaju, K.; Vishwanath, B.S.; Gowda, T.V.; Girish, K.S. The anti-snake venom properties of Tamarindus indica (leguminosae) seed extract. Phytother. Res. 2006, 20, 851–858. [Google Scholar] [CrossRef]
- Leanpolchareanchai, J.; Pithayanukul, P.; Bavovada, R. Anti-necrosis potential of polyphenols against snake venoms. Immunopharmacol. Immunotoxicol. 2009, (in press). [Google Scholar]
- Arni, R.K.; Ward, R.J. Phospholipase A2—a structural review. Toxicon 1996, 34, 827–841. [Google Scholar] [CrossRef]
- Haslam, E. Natural polyphenols (Vegetable tannins) as drugs: Possible modes of action. J. Nat. Prod. 1996, 59, 205–215. [Google Scholar] [CrossRef]
- Bjarnason, J.B.; Fox, J.W. Hemorrhagic toxins from snake venoms. J. Toxicol. Toxin Rev. 1988, 7, 121–209. [Google Scholar] [CrossRef]
- Gutiérrez, J.M.; Avila, C.; Rojas, E.; Cerdas, L. An alternative in vitro method for testing the potency of the polyvalent antivenom produced in Costa Rica. Toxicon 1988, 26, 411–413. [Google Scholar] [CrossRef]
- Ferrante, N.D. Turbidimetric measurement of acid mucopolysaccharides and hyaluronidase activity. J. Biol. Chem. 1956, 220, 303–306. [Google Scholar]
- Yingprasertchai, S.; Bunyasrisawat, S.; Ratanabanangkoon, K. Hyaluronidase inhibitors (sodium cromoglycate and sodium auro-thiomalate) reduce the local tissue damage and prolong the survival time of mice injected with Naja kaouthia and Calloselasma rhodostoma venoms. Toxicon 2003, 42, 635–646. [Google Scholar] [CrossRef]
- Tan, N.H.; Tan, C.S. A comparative study of cobra (Naja) venom enzymes. Comp. Biochem. Physiol. B 1988, 90, 745–750. [Google Scholar]
- Kondo, H.; Kondo, S.; Ikezawa, H.; Murata, R.; Ohsaka, A. Studies on the quantitative method for determination of hemorrhagic activity of Habu snake venom. Jpn. J. Med. Sci. Biol. 1960, 13, 43–51. [Google Scholar]
- Gutiérrez, J.M.; Gené, J.A.; Rojas, G.; Cerdas, L. Neutralization of proteolytic and hemorrhagic activities of Costa Rican snake venoms by a polyvalent antivenom. Toxicon 1985, 23, 887–893. [Google Scholar] [CrossRef]
- Theakston, R.D.G.; Reid, H.A. Development of simple standard assay procedures for the characterization of snake venoms. Bull. World Health Organ. 1983, 61, 949–956. [Google Scholar]
- Frisch, M.J.; Trucks, G.W.; Schlegel, H.R.; Scuseria, G.E.; Robb, M.A.; Cheeseman, J.R.; Montgomery, Jr. J.A.; Vreven, T.; Kudin, K.N.; Burant, J.C.; Millam, J.M.; Iyengar, S.S.; Tomasi, J.; Barone, V.; Mennucci, B.; Cossi, M.; Scalmani, G.; Rega, N.; Petersson, G.A.; Nakatsuji, H.; Hada, M.; Ehara, M.; Toyota, K.; Fukuda, R.; Hasegawa, J.; Ishida, M.; Nakajima, T.; Honda, Y.; Kitao, O.; Nakai, H.; Klene, M.; Li, X.; Knox, J.E.; Hratchian, H.P.; Cross, J.B.; Bakken, V.; Adamo, C.; Jaramillo, J.; Gomperts, R.; Stratmann, R.E.; Yazyev, O.; Austin, A.J.; Cammi, R.; Pomelli, C.; Ochterski, J.W.; Ayala, P.Y.; Morokuma, K.; Voth, G.A.; Salvador, P.; Dannenberg, J.J.; Zakrzewski, V.G.; Dapprich, S.; Daniels, A.D.; Strain, M.C.; Farkas, O.; Malick, D.K.; Rabuck, A.D.; Raghavachari, K.; Foresman, J.B.; Ortiz, J.V.; Cui, Q.; Baboul, A.G.; Clifford, S.; Cioslowski, J.; Stefanov, B.B.; Liu, G.; Liashenko, A.; Piskorz, P.; Komaromi, I.; Martin, R.L.; Fox, D.J.; Keith, T.; Al-Laham, M.A.; Peng, C.Y.; Nanayakkara, A.; Challacombe, M.; Gill, P.M.W.; Johnson, B.; Chen, W.; Wong, M.W.; Gonzalez, C.; Pople, J.A. Gaussian03; Gaussian, Inc.: Pittsburgh, PA, USA, 2003. [Google Scholar]
- Combet, C.; Jambon, M.; Deléage, G.; Geourjon, C. Geno3D: Automatic comparative molecular modelling of protein. Bioinformatics 2002, 18, 213–214. [Google Scholar] [CrossRef]
- Laskowski, R.A.; MacArthur, M.W.; Moss, D.S.; Thornton, J.M. PROCHECK: A program to check the stereochemical quality of protein structures. J. Appl. Crystallogr. 1993, 26, 283–291. [Google Scholar] [CrossRef]
- Tsai, I.H.; Wang, Y.M.; Au, L.C.; Ko, T.P.; Chen, Y.H.; Chu, Y.F. Phospholipases A2 from Callosellasma rhodostoma venom gland cloning and sequencing of 10 of the cDNAs, three-dimensional modelling and chemical modification of the major isozyme. Eur. J. Biochem. 2000, 267, 6684–6691. [Google Scholar] [CrossRef]
- Joubert, F.J.; Taljaard, N. Purification, some properties and amino-acid sequences of two phospholipases A (CM-II and CM-III) from Naja naja kaouthia venom. Eur. J. Biochem. 1980, 112, 493–499. [Google Scholar] [CrossRef]
- Jones, G.; Willett, P.; Glen, R.C.; Leach, A.R.; Taylor, R. Development and validation of a genetic algorithm for flexible docking. J. Mol. Biol. 1997, 267, 727–748. [Google Scholar]
- Brady, G.P.; Stouten, P.F.W. Fast prediction and visualization of protein binding pockets with PASS. J. Comput. Aided Mol. Des. 2000, 14, 383–401. [Google Scholar] [CrossRef]
- Gu, L.; Wang, Z.; Song, S.; Shu, Y.; Lin, Z. Crystal structures of an acidic phospholipase A2 from the venom of Naja kaouthia. Toxicon 2002, 40, 917–922. [Google Scholar] [CrossRef]
- Sample Availability: Samples are available from authors.
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Leanpolchareanchai, J.; Pithayanukul, P.; Bavovada, R.; Saparpakorn, P. Molecular Docking Studies and Anti-enzymatic Activities of Thai Mango Seed Kernel Extract Against Snake Venoms. Molecules 2009, 14, 1404-1422. https://doi.org/10.3390/molecules14041404
Leanpolchareanchai J, Pithayanukul P, Bavovada R, Saparpakorn P. Molecular Docking Studies and Anti-enzymatic Activities of Thai Mango Seed Kernel Extract Against Snake Venoms. Molecules. 2009; 14(4):1404-1422. https://doi.org/10.3390/molecules14041404
Chicago/Turabian StyleLeanpolchareanchai, Jiraporn, Pimolpan Pithayanukul, Rapepol Bavovada, and Patchreenart Saparpakorn. 2009. "Molecular Docking Studies and Anti-enzymatic Activities of Thai Mango Seed Kernel Extract Against Snake Venoms" Molecules 14, no. 4: 1404-1422. https://doi.org/10.3390/molecules14041404
APA StyleLeanpolchareanchai, J., Pithayanukul, P., Bavovada, R., & Saparpakorn, P. (2009). Molecular Docking Studies and Anti-enzymatic Activities of Thai Mango Seed Kernel Extract Against Snake Venoms. Molecules, 14(4), 1404-1422. https://doi.org/10.3390/molecules14041404