SOCS3-Mediated Blockade Reveals Major Contribution of JAK2/STAT5 Signaling Pathway to Lactation and Proliferation of Dairy Cow Mammary Epithelial Cells in Vitro

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
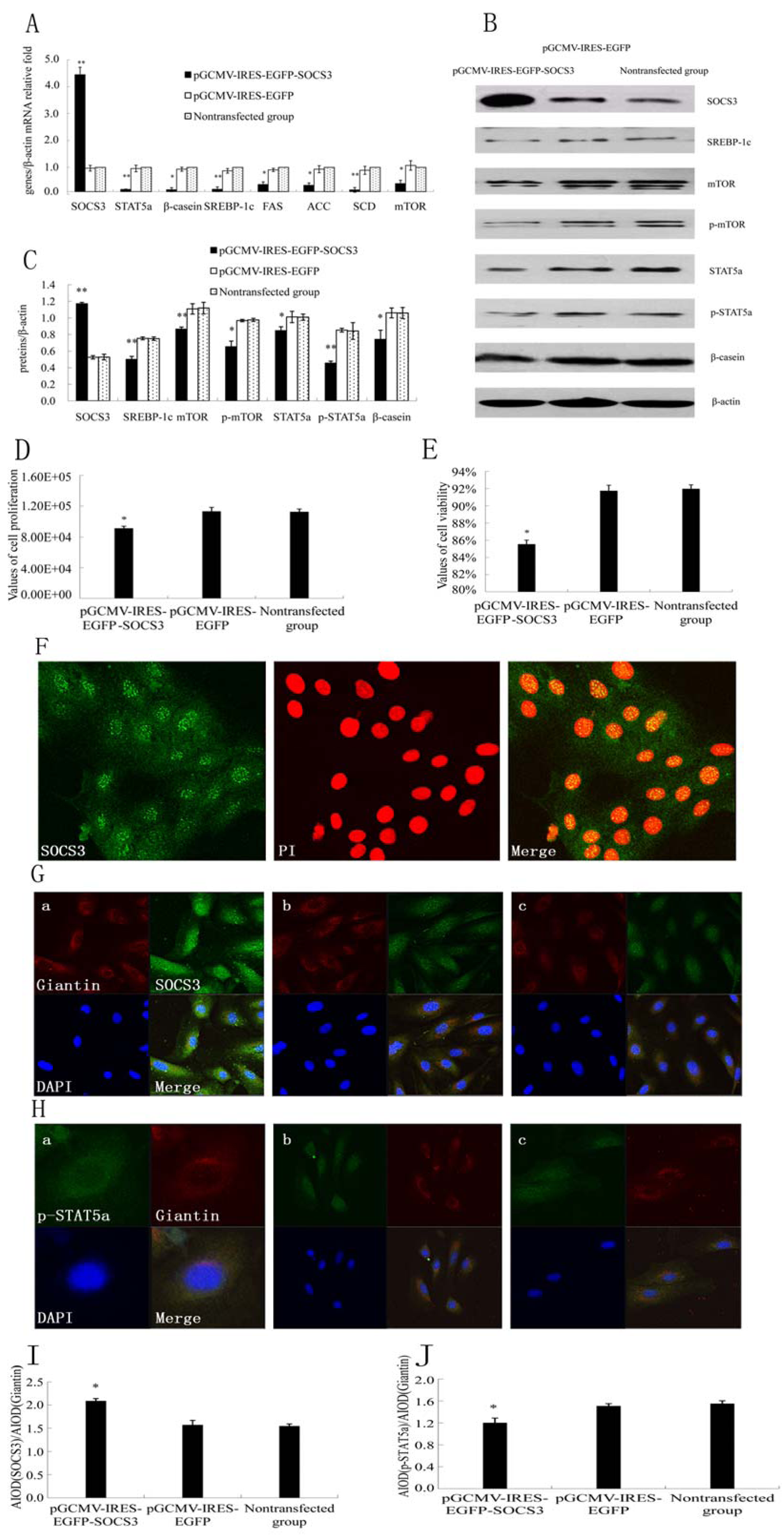
2.1. SOCS3 Overexpression Decreased Cell Proliferation and Negatively Regulated JAK2/STAT5a Signaling Pathway as well as Fatty Acid Synthesis in DCMECs
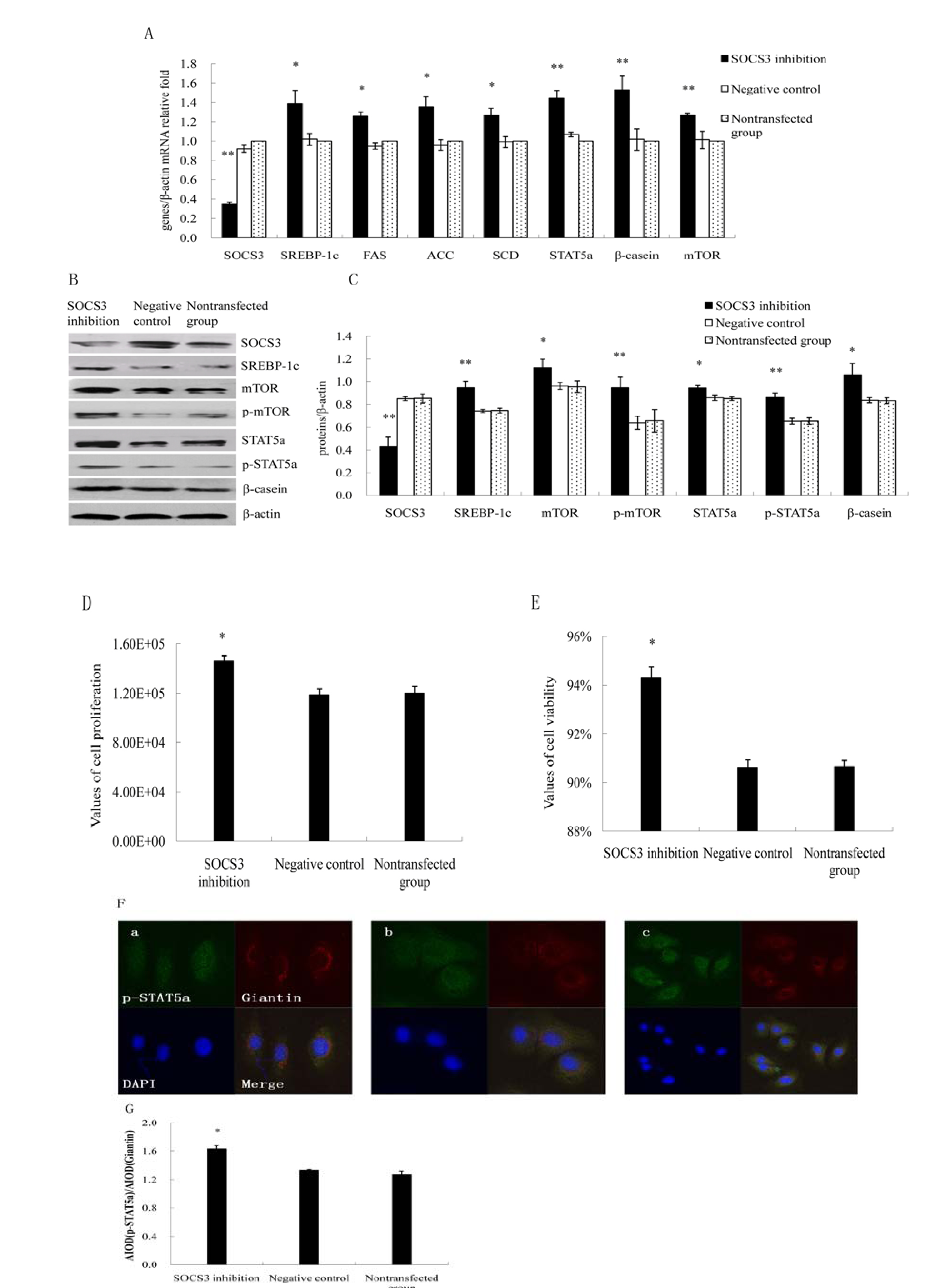
2.2. Inhibition of SOCS3 Increased the Expressions of Key Genes on Lactation and Proliferation of DCMECs


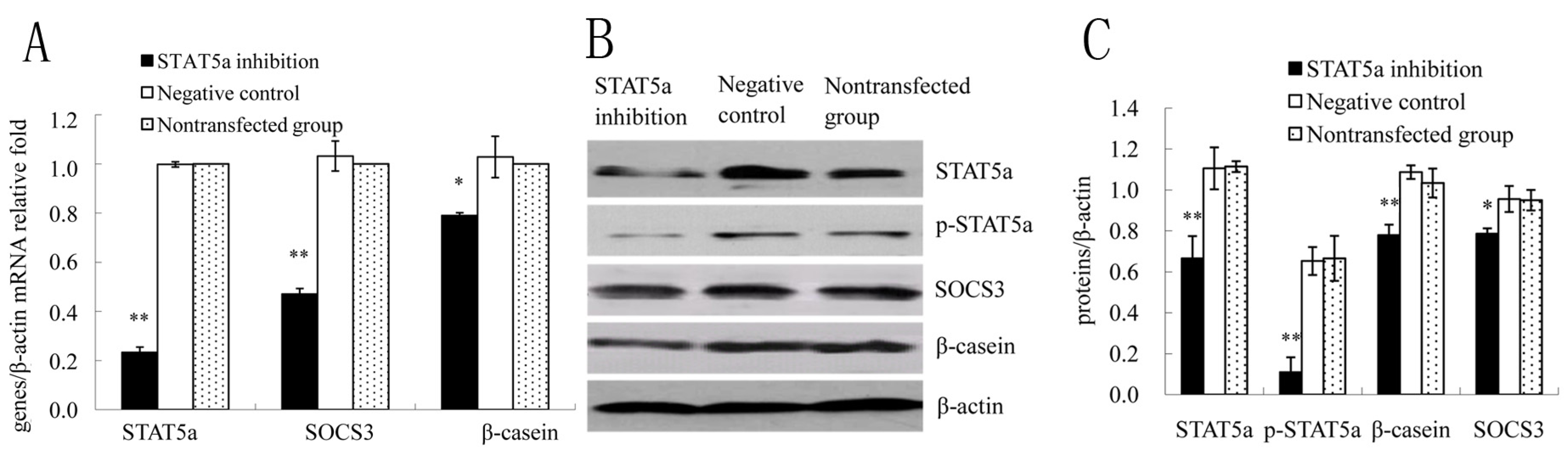
2.3. STAT5a was Required for SOCS3 Activation in DCMECs

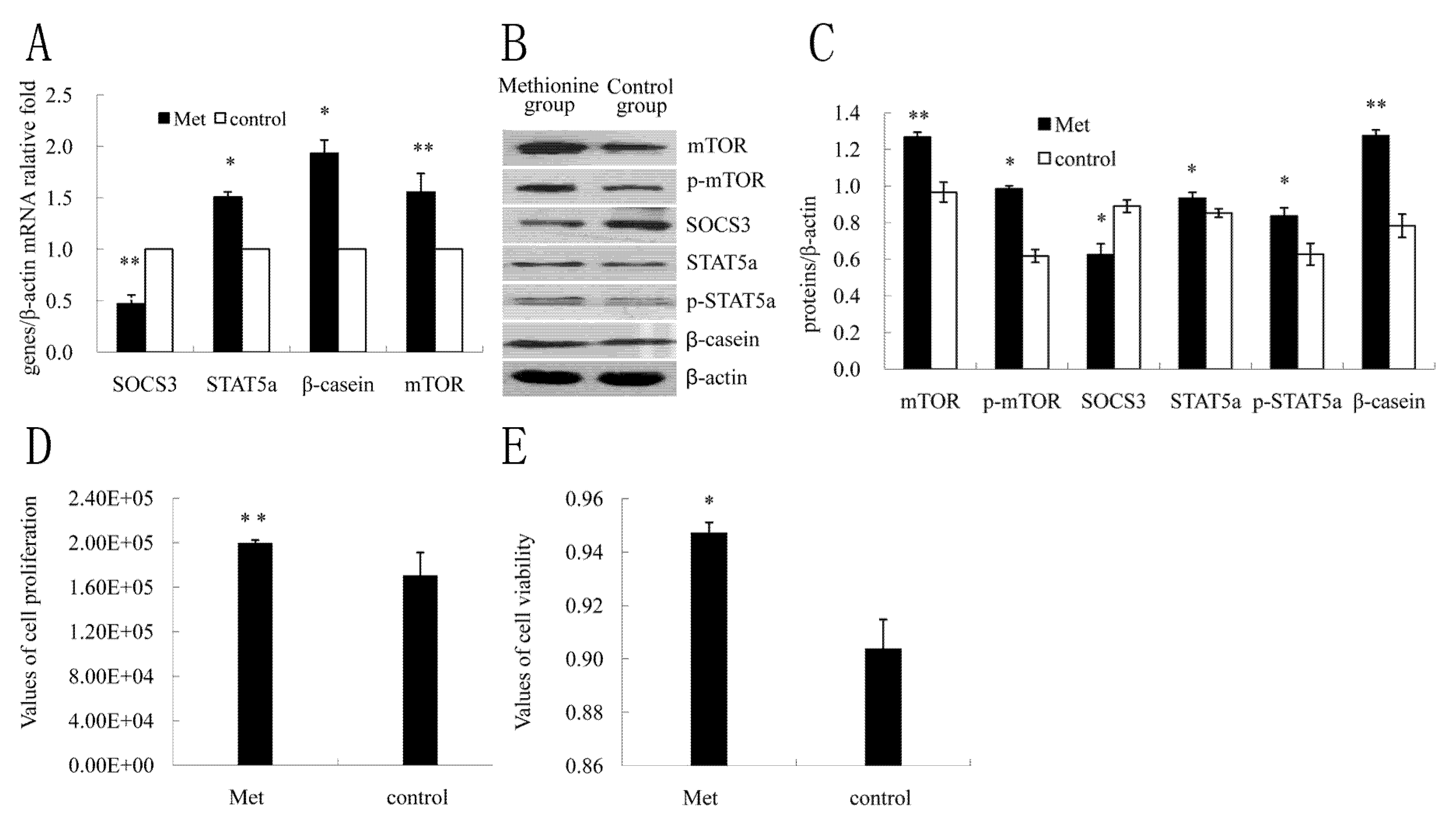
2.4. The Effect of Met on SOCS3 to Regulate β-Casein Expression and Cell Proliferation

2.5. Discussion
3. Experimental
3.1. Cell Preparation and Treatments
3.2. Cell Viability Assay
3.3. RNA Extraction and Quantitative Real-Time PCR
3.4. Immunofluorescence
3.5. Western Blot Analysis
3.6. Construction of SOCS3 Eukaryotic Expression Plasmid and Transfection
3.7. Small Interfering RNA Transfection
3.8. Statistical Analysis
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Starr, R.; Willson, T.A.; Viney, E.M.; Murray, L.; Rayner, J.R.; Jenkins, B.J.; Gonda, T.J.; Alexander, W.S.; Metcalf, D.; Nicola, N.A. A family of cytokine-inducible inhibitors of signalling. Nature 1997, 387, 917–921. [Google Scholar] [CrossRef]
- Naka, T.; Narazaki, M.; Hirata, M.; Matsumoto, T.; Minamoto, S.; Aono, A.; Nishimoto, N.; Kajita, T.; Taga, T.; Yoshizaki, K. STAT-induced STAT inhibitor. Nature 1997, 387, 924–929. [Google Scholar] [CrossRef]
- Endo, T.A.; Masuhara, M.; Yokouchi, M.; Suzuki, R.; Sakamoto, H.; Mitsui, K.; Matsumoto, A.; Tanimura, S.; Ohtsubo, M.; Misawa, H. A new protein containing an SH2 domain that inhibits JAK kinases. Nature 1997, 387, 921–924. [Google Scholar] [CrossRef]
- Sachithanandan, N.; Fam, B.C.; Fynch, S.; Dzamko, N.; Watt, M.J.; Wormald, S.; Honeyman, J.; Galic, S.; Proietto, J.; Andrikopoulos, S. Liver-pecific suppressor of cytokine signaling-3 deletion in mice enhances hepatic insulin sensitivity and lipogenesis resulting in fatty liver and obesity1. Hepatology 2010, 52, 1632–1642. [Google Scholar] [CrossRef]
- Trengove, M.C.; Ward, A.C. SOCS proteins in development and disease. Am. J. Clin. Exp. Immunol. 2013, 2, 1–29. [Google Scholar]
- Yoshimura, A.; Naka, T.; Kubo, M. SOCS proteins, cytokine signalling and immune regulation. Nat. Rev. Immunol. 2007, 7, 454–465. [Google Scholar]
- Tonko-Geymayer, S.; Goupille, O.; Tonko, M.; Soratroi, C.; Yoshimura, A.; Streuli, C.; Ziemiecki, A.; Kofler, R.; Doppler, W. Regulation and function of the cytokine-inducible SH-2 domain proteins, CIS and SOCS3, in mammary epithelial cells. Mol. Endocrinol. 2002, 16, 1680–1695. [Google Scholar] [CrossRef]
- Hadsell, D.L.; Parlow, A.F.; Torres, D.; George, J.; Olea, W. Enhancement of maternal lactation performance during prolonged lactation in the mouse by mouse GH and long-R3-IGF-I is linked to changes in mammary signaling and gene expression. J. Endocrinol. 2008, 198, 61–70. [Google Scholar] [CrossRef]
- Barclay, J.L.; Anderson, S.T.; Waters, M.J.; Curlewis, J.D. SOCS3 as a tumor suppressor in breast cancer cells, and its regulation by PRL. Int. J. Cancer 2009, 124, 1756–1766. [Google Scholar] [CrossRef]
- Anderson, S.T.; Isa, N.N.; Barclay, J.L.; Waters, M.J.; Curlewis, J.D. Maximal expression of suppressors of cytokine signaling in the rat ovary occurs in late pregnancy. Reproduction 2009, 138, 537–544. [Google Scholar] [CrossRef]
- Robinson, G.W.; Pacher-Zavisin, M.; Zhu, B.M.; Yoshimura, A.; Hennighausen, L. Socs3 modulates the activity of the transcription factor Stat3 in mammary tissue and controls alveolar homeostasis. Dev. Dyn. 2007, 236, 654–661. [Google Scholar] [CrossRef]
- Dif, F.; Saunier, E.; Demeneix, B.; Kelly, P.A.; Edery, M. Cytokine-inducible SH2-containing protein suppresses PRL signaling by binding the PRL receptor. Endocrinology 2001, 142, 5286–5293. [Google Scholar] [CrossRef]
- Paukku, K.; Silvennoinen, O. STATs as critical mediators of signal transduction and transcription: Lessons learned from STAT5. Cytokine Growth Factor Rev. 2004, 15, 435–455. [Google Scholar] [CrossRef]
- Barash, I. Stat5 in the mammary gland: controlling normal development and cancer. J. Cell. Physiol. 2006, 209, 305–313. [Google Scholar] [CrossRef]
- Dong, B.; Zhao, F.Q. Involvement of the ubiquitous Oct-1 transcription factor in hormonal induction of beta-casein gene expression. Biochem. J. 2007, 401, 57–64. [Google Scholar] [CrossRef]
- Khatib, H.; Monson, R.; Schutzkus, V.; Kohl, D.; Rosa, G.; Rutledge, J. Mutations in the STAT5A gene are associated with embryonic survival and milk composition in cattle. J. Dairy Sci. 2008, 91, 784–793. [Google Scholar]
- He, F.; Sun, D. X; Yu, Y. SNPs detection of STAT5A gene and association with milk production traits in Holstein cattle. Chin. J. Anim. Vet. Sci. 2007, 38, 326–331. [Google Scholar]
- Bionaz, M.; Loor, J.J. Gene networks driving bovine mammary protein synthesis during the lactation cycle. Bioinforma. Biol. Insights 2011, 5, 83–98. [Google Scholar]
- Bakan, I.; Laplante, M. Connecting mTORC1 signaling to SREBP-1 activation. Curr. Opin. Lipidol. 2012, 23, 226–234. [Google Scholar] [CrossRef]
- Yamamoto, T.; Watanabe, K.; Inoue, N.; Nakagawa, Y.; Ishigaki, N.; Matsuzaka, T.; Takeuchi, Y.; Kobayashi, K.; Yatoh, S.; Takahashi, A. Protein kinase Cbeta mediates hepatic induction of sterol-regulatory element binding protein-1c by insulin. J. Lipid Res. 2010, 51, 1859–1870. [Google Scholar] [CrossRef]
- Ueki, K.; Kadowaki, T.; Kahn, C.R. Role of suppressors of cytokine signaling SOCS-1 and SOCS-3 in hepatic steatosis and the metabolic syndrome. Hepatol. Res. 2005, 33, 185–192. [Google Scholar] [CrossRef]
- Orellana, R.A.; Jeyapalan, A.; Escobar, J.; Frank, J.W.; Nguyen, H.V.; Suryawan, A.; Davis, T.A. Amino acids augment muscle protein synthesis in neonatal pigs during acute endotoxemia by stimulating mTOR-dependent translation initiation. Am. J. Physiol.-Endocrinol. Metab. 2007, 293, E1416–E1425. [Google Scholar] [CrossRef]
- Correa-Fiz, F.; Reyes-Palomares, A.; Fajardo, I.; Melgarejo, E.; Gutierrez, A.; GarcIa-Ranea, J.A.; Medina, M.A.; Sanchez-Jimenez, F. Regulatory cross-talk of mouse liver polyamine and methionine metabolic pathways: A systemic approach to its physiopathological consequences. Amino Acids 2011, 42, 577–595. [Google Scholar]
- Honda, M.; Takehana, K.; Sakai, A.; Tagata, Y.; Shirasaki, T.; Nishitani, S.; Muramatsu, T.; Yamashita, T.; Nakamoto, Y.; Mizukoshi, E. Malnutrition impairs interferon signaling through mTOR and FoxO pathways in patients with chronic hepatitis C. Gastroenterology 2011, 141, 128–140. [Google Scholar] [CrossRef]
- Lu, L.M.; Gao, X.J.; Li, Q.Z.; Huang, J.G.; Liu, R.; Li, H.M. Comparative phosphoproteomics analysis of the effects of L-methionine on dairy cow mammary epithelial cells. Can. J. Anim. Sci. 2012, 92, 433–442. [Google Scholar] [CrossRef]
- Peltola, K.J.; Paukku, K.; Aho, T.L.; Ruuska, M.; Silvennoinen, O.; Koskinen, P.J. Pim-1 kinase inhibits STAT5-dependent transcription via its interactions with SOCS1 and SOCS3. Blood 2004, 103, 3744–3750. [Google Scholar] [CrossRef]
- Helman, D.; Sandowski, Y.; Cohen, Y.; Matsumoto, A.; Yoshimura, A.; Merchav, S.; Gertler, A. Cytokine-inducible SH2 protein (CIS3) and JAK2 binding protein (JAB) abolish prolactin receptor-mediated STAT5 signaling. FEBS Lett. 1998, 441, 287–291. [Google Scholar] [CrossRef]
- Tam, S.P.; Lau, P.; Djiane, J.; Hilton, D.J.; Waters, M.J. Tissue-specific induction of SOCS gene expression by PRL. Endocrinology 2001, 142, 5015–5026. [Google Scholar] [CrossRef]
- White, G.E.; Cotterill, A.; Addley, M.R.; Soilleux, E.J.; Greaves, D.R. Suppressor of cytokine signalling protein SOCS3 expression is increased at sites of acute and chronic inflammation. J. Mol. Histol. 2011, 42, 137–151. [Google Scholar] [CrossRef]
- Jiang, S.Z.; Ren, Z.R.; Xie, F.; Yan, J.B.; Huang, S.Z.; Zeng, Y.T. Bovine prolactin elevates hTF expression directed by a tissue-specific goat β-casein promoter through prolactin receptor-mediated STAT5a activation. Biotechnol. Lett. 2012, 34, 1991–1999. [Google Scholar] [CrossRef]
- He, X.; Chu, M.X.; Qiao, L.; He, J.N.; Wang, P.Q.; Feng, T.; Di, R.; Cao, G.L.; Fang, L.; An, Y.F. Polymorphisms of STAT5A gene and their association with milk production traits in Holstein cows. Mol. Biol. Rep. 2012, 39, 2901–2907. [Google Scholar]
- Chen, C.-C.; Boxer, R.B.; Stairs, D.B.; Portocarrero, C.P.; Horton, R.H.; Alvarez, J.V.; Birnbaum, M.J.; Chodosh, L.A. Akt is required for Stat5 activation and mammary differentiation. Breast Cancer Res. 2010, 12, R72. [Google Scholar] [CrossRef]
- Cui, Y.; Riedlinger, G.; Miyoshi, K.; Tang, W.; Li, C.; Deng, C.-X.; Robinson, G.W.; Hennighausen, L. Inactivation of Stat5 in mouse mammary epithelium during pregnancy reveals distinct functions in cell proliferation, survival, and differentiation. Mol. Cell. Biol. 2004, 24, 8037–8047. [Google Scholar] [CrossRef]
- Lee, K.-H.; Moon, K.-J.; Kim, H.S.; Yoo, B.C.; Park, S.; Lee, H.; Kwon, S.; Lee, E.S.; Yoon, S. Increased cytoplasmic levels of CIS, SOCS1, SOCS2, or SOCS3 are required for nuclear translocation. FEBS Lett. 2008, 582, 2319–2324. [Google Scholar] [CrossRef]
- Ehrentraut, S.; Nagel, S.; Scherr, M.E.; Schneider, B.; Quentmeier, H.; Geffers, R.; Kaufmann, M.; Meyer, C.; Prochorec-Sobieszek, M.; Ketterling, R.P. t (8; 9)(p22; p24)/PCM1-JAK2 activates SOCS2 and SOCS3 via STAT5. PLoS One 2013, 8, e53767. [Google Scholar]
- Le Provost, F.; Miyoshi, K.; Vilotte, J.-L.; Bierie, B.; Robinson, G.W.; Hennighausen, L. SOCS3 promotes apoptosis of mammary differentiated cells. Biochem. Biophys. Res. Commun. 2005, 338, 1696–1701. [Google Scholar] [CrossRef]
- Chueh, F.-Y.; Leong, K.-F.; Cronk, R.J.; Venkitachalam, S.; Pabich, S.; Yu, C.-L. Nuclear localization of pyruvate dehydrogenase complex-E2 (PDC-E2), a mitochondrial enzyme, and its role in signal transducer and activator of transcription 5 (STAT5)-dependent gene transcription. Cell Signal. 2011, 23, 1170–1178. [Google Scholar] [CrossRef]
- Toerien, C.A.; Trout, D.R.; Cant, J.P. Nutritional stimulation of milk protein yield of cows is associated with changes in phosphorylation of mammary eukaryotic initiation factor 2 and ribosomal s6 kinase 1. J. Nutr. 2010, 140, 285–292. [Google Scholar] [CrossRef]
- Galbaugh, T.; Cerrito, M.G.; Jose, C.C.; Cutler, M.L. EGF-induced activation of Akt results in mTOR-dependent p70S6 kinase phosphorylation and inhibition of HC11 cell lactogenic differentiation. BMC Cell Biol. 2006, 7, 34. [Google Scholar] [CrossRef]
- Stahl, A.; Joyal, J.-S.; Chen, J.; Sapieha, P.; Juan, A.M.; Hatton, C.J.; Pei, D.T.; Hurst, C.G.; Seaward, M.R.; Krah, N.M. SOCS3 is an endogenous inhibitor of pathologic angiogenesis. Blood 2012, 120, 2925–2929. [Google Scholar] [CrossRef]
- Boutinaud, M.; Guinard-Flament, J.; Jammes, H. The number and activity of mammary epithelial cells, determining factors for milk production. Reprod. Nutr. Dev. 2004, 44, 499–508. [Google Scholar] [CrossRef]
- He, B.; You, L.; Uematsu, K.; Zang, K.; Xu, Z.; Lee, A.Y.; Costello, J.F.; McCormick, F.; Jablons, D.M. SOCS-3 is frequently silenced by hypermethylation and suppresses cell growth in human lung cancer. Proc. Natl. Acad. Sci. USA 2003, 100, 14133–14138. [Google Scholar] [CrossRef]
- Elam, M.B.; Yellaturu, C.; Howell, G.E.; Deng, X.; Cowan, G.S.; Kumar, P.; Park, E.A.; Hiler, M.L.; Wilcox, H.G.; Hughes, T.A. Dysregulation of sterol regulatory element binding protein-1c in livers of morbidly obese women is associated with altered suppressor of cytokine signaling-3 and signal transducer and activator of transcription-1 signaling. Metabolism 2010, 59, 587–598. [Google Scholar] [CrossRef]
- Ueki, K.; Kondo, T.; Tseng, Y.-H.; Kahn, C.R. Central role of suppressors of cytokine signaling proteins in hepatic steatosis, insulin resistance, and the metabolic syndrome in the mouse. Proc. Natl. Acad. Sci. USA 2004, 101, 10422–10427. [Google Scholar] [CrossRef]
- Ma, L.; Corl, B.A. Transcriptional regulation of lipid synthesis in bovine mammary epithelial cells by sterol regulatory element binding protein-1. J. Dairy Sci. 2012, 95, 3743–3755. [Google Scholar] [CrossRef]
- Han, J.M.; Jeong, S.J.; Park, M.C.; Kim, G.; Kwon, N.H.; Kim, H.K.; Ha, S.H.; Ryu, S.H.; Kim, S. Leucyl-tRNA synthetase is an intracellular leucine sensor for the mTORC1-signaling pathway. Cell 2012, 149, 410–424. [Google Scholar] [CrossRef]
- Prizant, R.L.; Barash, I. Negative effects of the amino acids Lys, His, and Thr on S6K1 phosphorylation in mammary epithelial cells. J. Cell. Biochem. 2008, 105, 1038–1047. [Google Scholar] [CrossRef]
- Lu, L.M.; Li, Q.Z.; Huang, J.G.; Gao, X.J. Proteomic and functional analyses reveal MAPK1 regulates milk protein synthesis. Molecules 2013, 18, 263–275. [Google Scholar]
- Huang, J.G.; Gao, X.J.; Li, Q.Z.; Lu, L.M.; Liu, R.; Luo, C.C.; Wang, J.L.; Qiao, B.; Jin, X. Proteomic analysis of the nuclear phosphorylated proteins in dairy cow mammary epithelial cells treated with estrogen. Vitro Cell. Dev. Biol.-Anim 2012, 48, 449–457. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the dairy cow mammary epithelial cells (DCMECs) are available from the authors.
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Huang, Y.-L.; Zhao, F.; Luo, C.-C.; Zhang, X.; Si, Y.; Sun, Z.; Zhang, L.; Li, Q.-Z.; Gao, X.-J. SOCS3-Mediated Blockade Reveals Major Contribution of JAK2/STAT5 Signaling Pathway to Lactation and Proliferation of Dairy Cow Mammary Epithelial Cells in Vitro. Molecules 2013, 18, 12987-13002. https://doi.org/10.3390/molecules181012987
Huang Y-L, Zhao F, Luo C-C, Zhang X, Si Y, Sun Z, Zhang L, Li Q-Z, Gao X-J. SOCS3-Mediated Blockade Reveals Major Contribution of JAK2/STAT5 Signaling Pathway to Lactation and Proliferation of Dairy Cow Mammary Epithelial Cells in Vitro. Molecules. 2013; 18(10):12987-13002. https://doi.org/10.3390/molecules181012987
Chicago/Turabian StyleHuang, Yu-Ling, Feng Zhao, Chao-Chao Luo, Xia Zhang, Yu Si, Zhe Sun, Li Zhang, Qing-Zhang Li, and Xue-Jun Gao. 2013. "SOCS3-Mediated Blockade Reveals Major Contribution of JAK2/STAT5 Signaling Pathway to Lactation and Proliferation of Dairy Cow Mammary Epithelial Cells in Vitro" Molecules 18, no. 10: 12987-13002. https://doi.org/10.3390/molecules181012987
APA StyleHuang, Y. -L., Zhao, F., Luo, C. -C., Zhang, X., Si, Y., Sun, Z., Zhang, L., Li, Q. -Z., & Gao, X. -J. (2013). SOCS3-Mediated Blockade Reveals Major Contribution of JAK2/STAT5 Signaling Pathway to Lactation and Proliferation of Dairy Cow Mammary Epithelial Cells in Vitro. Molecules, 18(10), 12987-13002. https://doi.org/10.3390/molecules181012987