Baeyer-Villiger Oxidation of Some C19 Steroids by Penicillium lanosocoeruleum

Abstract
:
1. Introduction
2. Results and Discussion


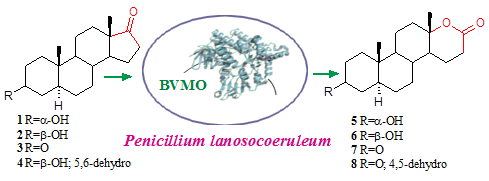{kind=link}
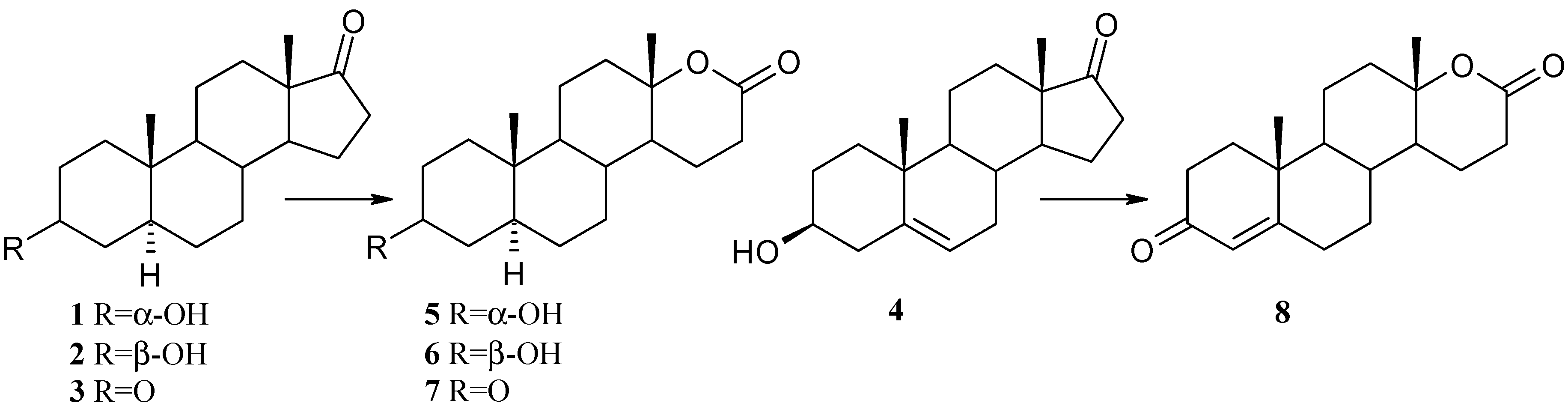{kind=link}
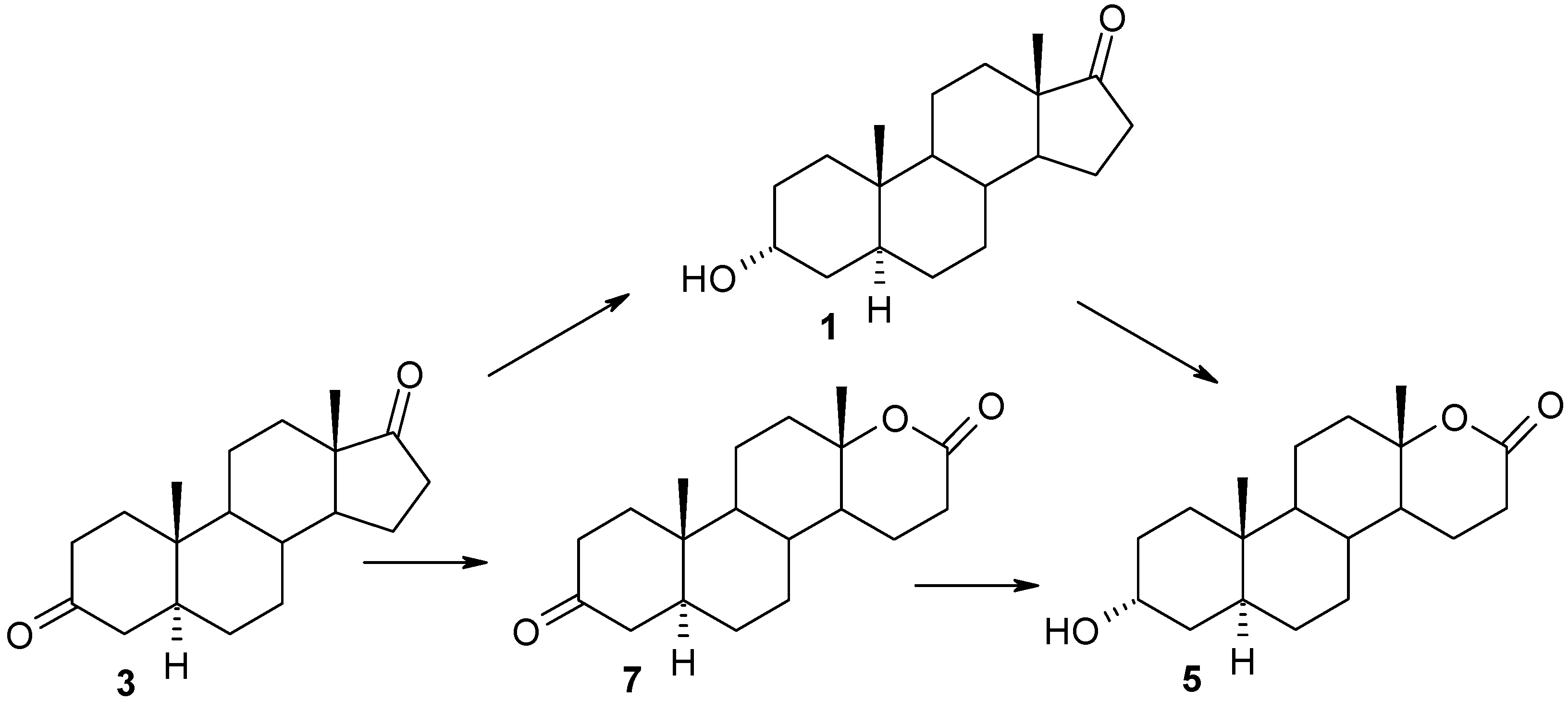{kind=link}
{kind=link}
| Starting material | Time (h) | Compounds | Yield (%) a,b |
|---|---|---|---|
| Androsterone (1) | 48 | Androsterone (1) | - |
| 3α-Hydroxy-17a-oxa-d-homo-5α-androstan-17-one (5) | 98 (96) | ||
| Epiandrosterone (2) | 48 | Epiandrosterone (2) | 19 (17) |
| 3β-Hydroxy-17a-oxa-d-homo-5α-androstan-17-one (6) | 75 (70) | ||
| Androstanedione (3) | 24 | Androstanedione (3) | - |
| 17a-Oxa-d-homo-5α-androstan-3,17-dione (7) | 40 (37) | ||
| 3α-Hydroxy-17a-oxa-d-homo-5α-androstan-17-one (5) | 59 (55) | ||
| DHEA (4) | 24 | DHEA (4) | - |
| Testololactone (8) | 96 (91) | ||
| 17a-Oxa-d-homo-5α-androstan-3,17-dione (7) | 48 | 17a-Oxa-d-homo-5α-androstan-3,17-dione (7) 3α-Hydroxy-17a-oxa-d-homo-5α-androstan-17-one (5) | 24 (23) 71 (67) |
| Substrate | Rt (min) | Compounds present in the mixture (%) a | Time of transformation (h) | ||||
|---|---|---|---|---|---|---|---|
| 4 | 8 | 12 | 24 | 48 | |||
| Androstanedione (3) | 3.75 | Androstanedione (3) | 100 | 95 | 26 | 5 | - |
| 3.15 | Androsterone (1) | - | 3 | 8 | - | - | |
| 7.19 | 3α-Hydroxy-17a-oxa-d-homo-5α-androstan-17-one (5) | - | - | 24 | 40 | 59 | |
| 7.60 | 17a-Oxa-d-homo-5α-androstan-3,17-dione (7) | - | 2 | 40 | 54 | 39 | |
| DHEA (4) | 3.39 | DHEA (4) | 85 | 26 | 1 | - | |
| 4.42 | Androstenedione (9) b | 15 | 63 | 58 | 4 | ||
| 8.42 | Testololactone (8) | - | 11 | 40 | 94 | ||
| 6.77 | 3β-hydroxy-17a-oxa-d-homo-androst-5-en-17-one c | - | - | - | - | ||


3. Experimental
3.1. General Conditions of Cultivation and Transformation
3.2. Chemicals and Reagents
3.3. Microorganism
3.4. Time Course Experiments
3.5. Isolation and Identification of the Products
3.6. Products Isolated in the Course of Transformations
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Świzdor, A.; Kołek, T.; Panek, A.; Milecka, N. Selective modifications of steroids performed by oxidative enzymes. Curr. Org. Chem. 2012, 16, 2551–2582. [Google Scholar] [CrossRef]
- Donova, M.V.; Egorova, O.V. Microbial steroid transformations: Current state and prospects. Appl. Microbiol. Biotechnol. 2012, 94, 1423–1447. [Google Scholar] [CrossRef]
- Beneventi, E.; Ottolina, G.; Carrea, G.; Panzeri, W.; Fronza, G.; Lau, P.C.K. Enzymatic Baeyer-Villiger oxidation of steroids with cyclopentadecanone monooxygenase. J. Mol. Catal. B: Enzym. 2009, 58, 164–168. [Google Scholar] [CrossRef] [Green Version]
- Van Beek, H.L.; de Gonzalo, G.; Fraaije, M.W. Blending Baeyer-Villiger monooxygenases: Using a robust BVMO as a scaffold for creating chimeric enzymes with novel catalytic properties. Chem. Commun. 2012, 48, 3288–3290. [Google Scholar] [CrossRef]
- Sebek, O.K.; Reineke, L.M.; Peterson, D.H. Intermediates in the metabolism of steroids by Penicillium lilacinum. J. Bacteriol. 1962, 83, 1327–1331. [Google Scholar]
- Kołek, T.; Szpineter, A.; Świzdor, A. Baeyer-Villiger oxidation of DHEA, pregnenolone, and androstenedione by Penicillium lilacinum AM111. Steroids 2008, 73, 1441–1445. [Google Scholar] [CrossRef]
- Liu, H.-M.; Li, H.; Shan, L.; Wu, J. Synthesis of steroidal lactones by Penicillium citreo-viride. Steroids 2006, 71, 931–934. [Google Scholar] [CrossRef]
- Huang, L.H.; Li, J.; Xu, G.; Zhang, X.-H.; Wang, Y.-G.; Yin, Y.-L.; Liu, H.-M. Biotransformation of dehydroepiandrosterone (DHEA) with Penicillium griseopurpureum Smith and Penicillium glabrum (Wehmer) Westling. Steroids 2010, 75, 1039–1046. [Google Scholar] [CrossRef]
- Kołek, T.; Szpineter, A.; Świzdor, A. Studies on Baeyer-Villiger oxidation of steroids: DHEA and pregnenolone d-lactonization pathways in Penicillium camemberti AM83. Steroids 2009, 74, 859–862. [Google Scholar]
- Bartmańska, A.; Dmochowska-Gładysz, J.; Huszcza, E. Steroids’ transformations in Penicillium notatum culture. Steroids 2005, 70, 193–198. [Google Scholar] [CrossRef]
- Brannon, R.; Parrish, F.W.; Wiley, B.J.; Long, L. Microbial transformation of series of androgens with Aspergillus tamarii. J. Org. Chem. 1967, 32, 1521–1527. [Google Scholar]
- Hunter, A.C.; Carragher, N.E. Flexibility of the endogenous progesterone lactonisation pathway in Aspergillus tamarii KITA: Transformation of a series of cortical steroid analogues. J. Steroid Biochem. Mol. Biol. 2003, 87, 301–308. [Google Scholar] [CrossRef]
- Hunter, A.C.; Kennedy, S.; Clabby, S.-J.; Elsom, J. Fate of novel Quasi reverse steroidal substrates by Aspergillus tamarii KITA: Bypass of lactonisation and an exclusive role for the minor hydroxylation pathway. Biochim. Biophys. Acta 2005, 1734, 190–197. [Google Scholar] [CrossRef]
- Hunter, A.C.; Elsom, J.; Ross, L.; Barrett, R. Ring-B functionalized androst-4-en-3-ones and ring-C substituted pregn-4-en-3-ones undergo differential transformation in Aspergillus tamarii KITA: Ring-A transformation with all C-6 substituted steroids and ring-D transformation with C-11 substituents. Biochim. Biophys. Acta 2006, 1761, 360–366. [Google Scholar] [CrossRef]
- Hunter, A.C.; Coyle, E.; Morse, F.; Dedi, C.; Dodd, H.T.; Koussoroplis, S.-J. Transformation of 5-ene steroids by the fungus Aspergillus tamarii KITA: Mixed molecular fate in lactonization and hydroxylation pathways with identification of a putative 3β-hydroxy-steroid dehydrogenase/Δ5-Δ4 isomerase pathway. Biochim. Biophys. Acta Mol. Cell. Biol. Lipids 2009, 1791, 110–117. [Google Scholar] [CrossRef]
- Hunter, A.C.; Khuenl-Brady, H.; Barrett, P.; Dodd, H.T.; Dedi, C. Transformation of some 3α-substituted steroids by Aspergillus tamarii KITA reveals stereochemical restriction of steroid binding orientation in the minor hydroxylation pathway. J. Steroid Biochem. Mol. Biol. 2010, 118, 171–176. [Google Scholar] [CrossRef]
- Hunter, A.C.; Collins, C.; Dodd, H.T.; Dedi, C.; Koussoroplis, S.-J. Transformation of a series of saturated isomeric steroidal diols by Aspergillus tamarii KITA reveals a precise stereochemical requirement for entrance into the lactonization pathway. J. Steroid Biochem. Mol. Biol. 2010, 122, 352–358. [Google Scholar] [CrossRef]
- Yildirim, K.; Uzuner, A.; Gulcuoglu, E. Biotransformation of some steroids by Aspergillus terreus MRC 200365. Collect. Czechoslov. Chem. Commun. 2010, 75, 665–673. [Google Scholar] [CrossRef]
- Yildirim, K.; Uzuner, A.; Gulcuoglu, E.Y. Baeyer-Villiger oxidation of some steroids by Aspergillus tamarii MRC 72400. Collect. Czechoslov. Chem. Commun. 2011, 76, 743–754. [Google Scholar] [CrossRef]
- Faramarzi, M.; Yazdi, M.T.; Amini, M.; Mohseni, F.A.; Zarrini, G.; Amani, A.; Shafiee, A. Microbial production of testosterone and testololactone in the culture of Aspergillus terreus. World J. Microbiol. Biotechnol. 2004, 20, 657–660. [Google Scholar] [CrossRef]
- Čapek, A.; Hanč, O. Microbiological transformation of steroids. XII. Transformation of steroids by different species and strains of Fusaria. Folia Microbiol. 1960, 5, 251–256. [Google Scholar] [CrossRef]
- Zhang, H.; Ren, J.; Wang, Y.; Sheng, C.; Wu, Q.; Diao, A.; Zhu, D. Effective multi-step functional biotransformations of steroids by a newly isolated Fusarium oxysporum SC1301. Tetrahedron 2013, 69, 184–189. [Google Scholar] [CrossRef]
- Al-Aboudi, A.; Mohammad, M.Y.; Musharraf, S.G.; Choudhary, M.I.; Atta-ur-Rahman. Microbial transformation of testosterone by Rhizopus stolonifer and Fusarium lini. Nat. Prod. Res. 2008, 22, 1498–1509. [Google Scholar] [CrossRef]
- Garrido, M.; Bratoeff, E.; Bonilla, D.; Soriano, J.; Heuze, Y.; Cabeza, M. New steroidal lactones as 5α-reductase inhibitors and antagonists for the androgen receptor. J. Steroid Biochem. Mol. Biol. 2011, 127, 367–373. [Google Scholar] [CrossRef]
- Balunas, M.J.; Su, B.; Brueggemeier, R.W.; Kinghorn, A.D. Natural products as aromatase inhibitors. Anticancer Agents Med. Chem. 2008, 8, 646–682. [Google Scholar]
- Eugster, E.A. Aromatase inhibitors in precocious puberty: Rationale and experience to date. Treat. Endocrinol. 2004, 3, 141–151. [Google Scholar] [CrossRef]
- Mendelovici, M.; Glotter, E. Epoxidation and Baeyer-Villiger oxidation of γ-hydroxy-α,β-unsaturated ketones on exposure to m-chloroperbenzoic acid. J. Chem. Soc. Perkin Trans. I 1992, 1735–1740. [Google Scholar] [CrossRef]
- Zinczuk, J.; Bacigaluppo, J.A.; Colombo, M.I.; Cravero, R.M.; González-Sierra, M.; Rúveda, E.A. An efficient and environmentally benign chemical synthesis of testolactone. J. Braz. Chem. Soc. 2003, 14, 970–974. [Google Scholar]
- Świzdor, A.; Kołek, T.; Panek, A.; Białońska, A. Microbial Baeyer-Villiger oxidation of steroidal ketones using Beauveria bassiana: Presence of an 11α-hydroxyl group essential to generation of d-homo lactones. Biochim. Biophys. Acta Mol. Cell. Biol. Lipids 2011, 1811, 253–262. [Google Scholar] [CrossRef]
- Hanson, J.R.; Hunter, A.C. The hydroxylation of steroidal ring D lactones by Cephalosporium aphidicola. Phytochemistry 1998, 49, 2349–2353. [Google Scholar] [CrossRef]
- Bridgeman, J.E.; Cherry, P.C.; Clegg, A.S.; Evans, J.M.; Jones, E.R.H.; Kasal, J.A.; Kumar, V.; Meakins, G.D.; Morisawa, Y.; Richards, E.E.; et al. Microbiological hydroxylation of steroids. Part I. Proton magnetic resonance spectra of ketones, alcohols, and acetates in the androstane, pregnane, and oestrane series. J. Chem. Soc. Perkin 1 1970, 250–257. [Google Scholar]
- Blunt, J.W.; Stothers, J.B. 13C N.m.r. spectra of steroids – A survey and commentary. Org. Magn. Reson. 1977, 9, 439–464. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the compounds 5–8 are available from the authors.
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Świzdor, A. Baeyer-Villiger Oxidation of Some C19 Steroids by Penicillium lanosocoeruleum. Molecules 2013, 18, 13812-13822. https://doi.org/10.3390/molecules181113812
Świzdor A. Baeyer-Villiger Oxidation of Some C19 Steroids by Penicillium lanosocoeruleum. Molecules. 2013; 18(11):13812-13822. https://doi.org/10.3390/molecules181113812
Chicago/Turabian StyleŚwizdor, Alina. 2013. "Baeyer-Villiger Oxidation of Some C19 Steroids by Penicillium lanosocoeruleum" Molecules 18, no. 11: 13812-13822. https://doi.org/10.3390/molecules181113812
APA StyleŚwizdor, A. (2013). Baeyer-Villiger Oxidation of Some C19 Steroids by Penicillium lanosocoeruleum. Molecules, 18(11), 13812-13822. https://doi.org/10.3390/molecules181113812