Anti-Proliferative and Apoptosis-Inducing Effects of Camptothecin-20(s)-O-(2-pyrazolyl-1)acetic Ester in Human Breast Tumor MCF-7 Cells


Abstract
:1. Introduction

2. Results and Discussion
2.1. Antiproliferative Activity of CPT6


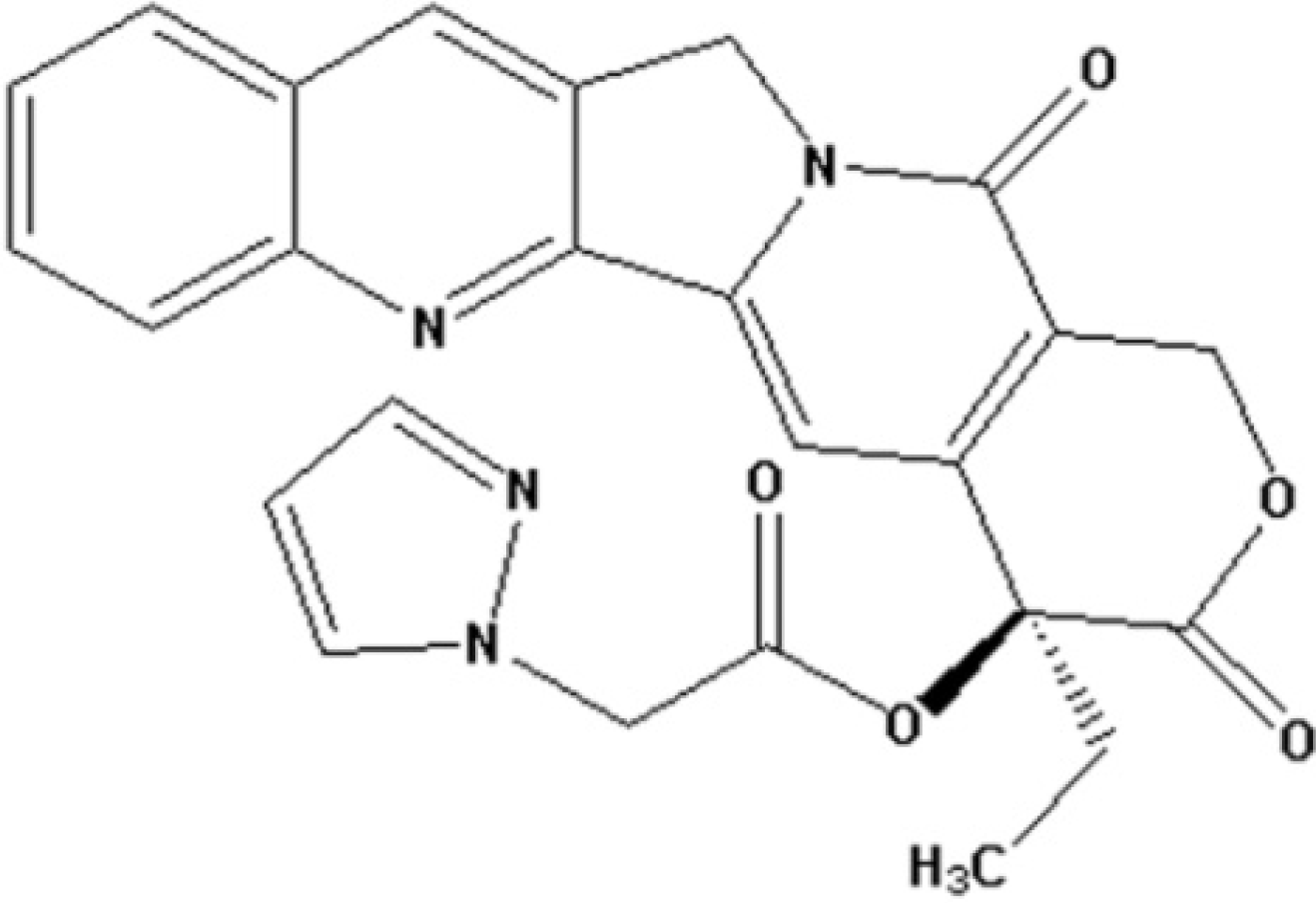{kind=link}
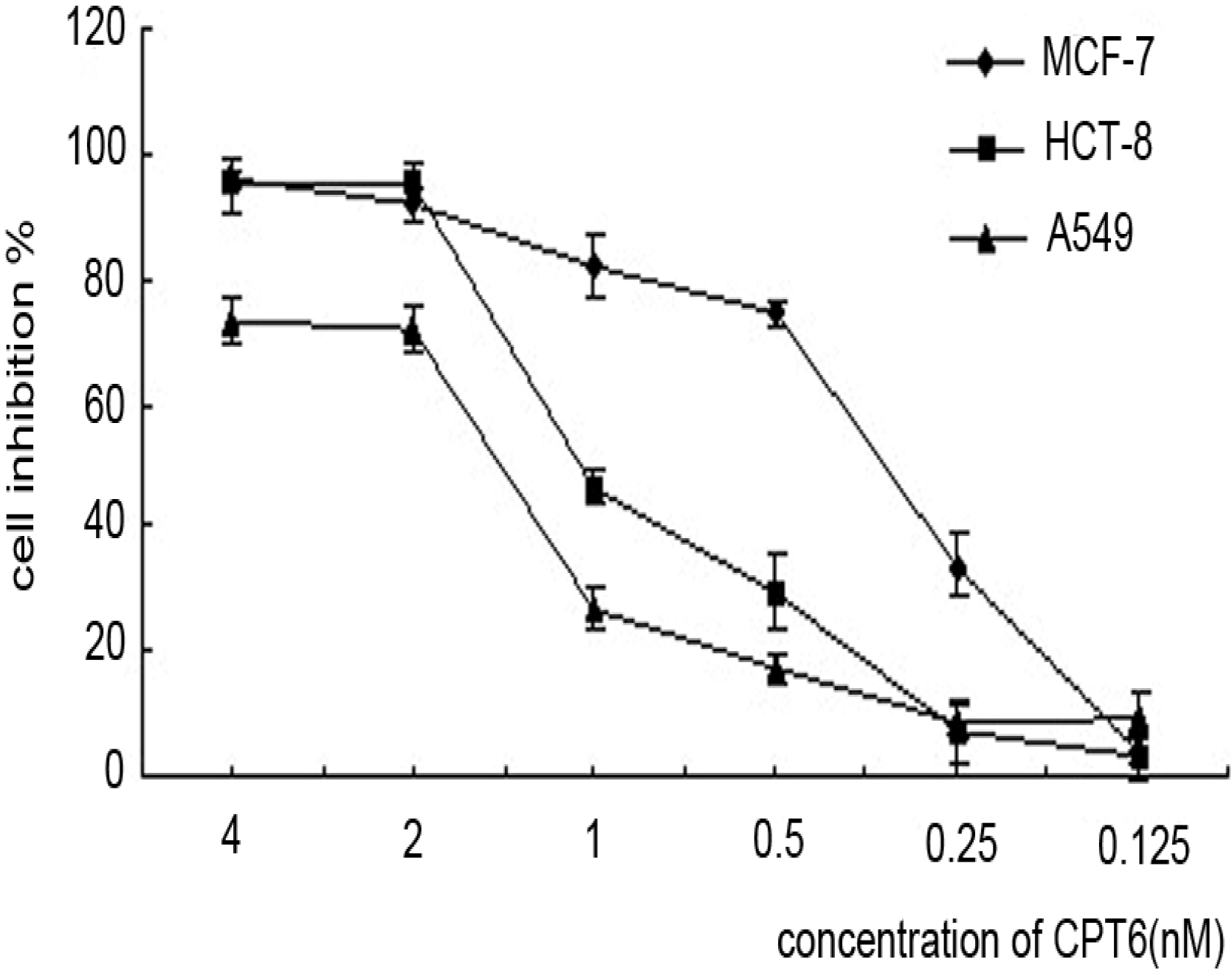{kind=link}
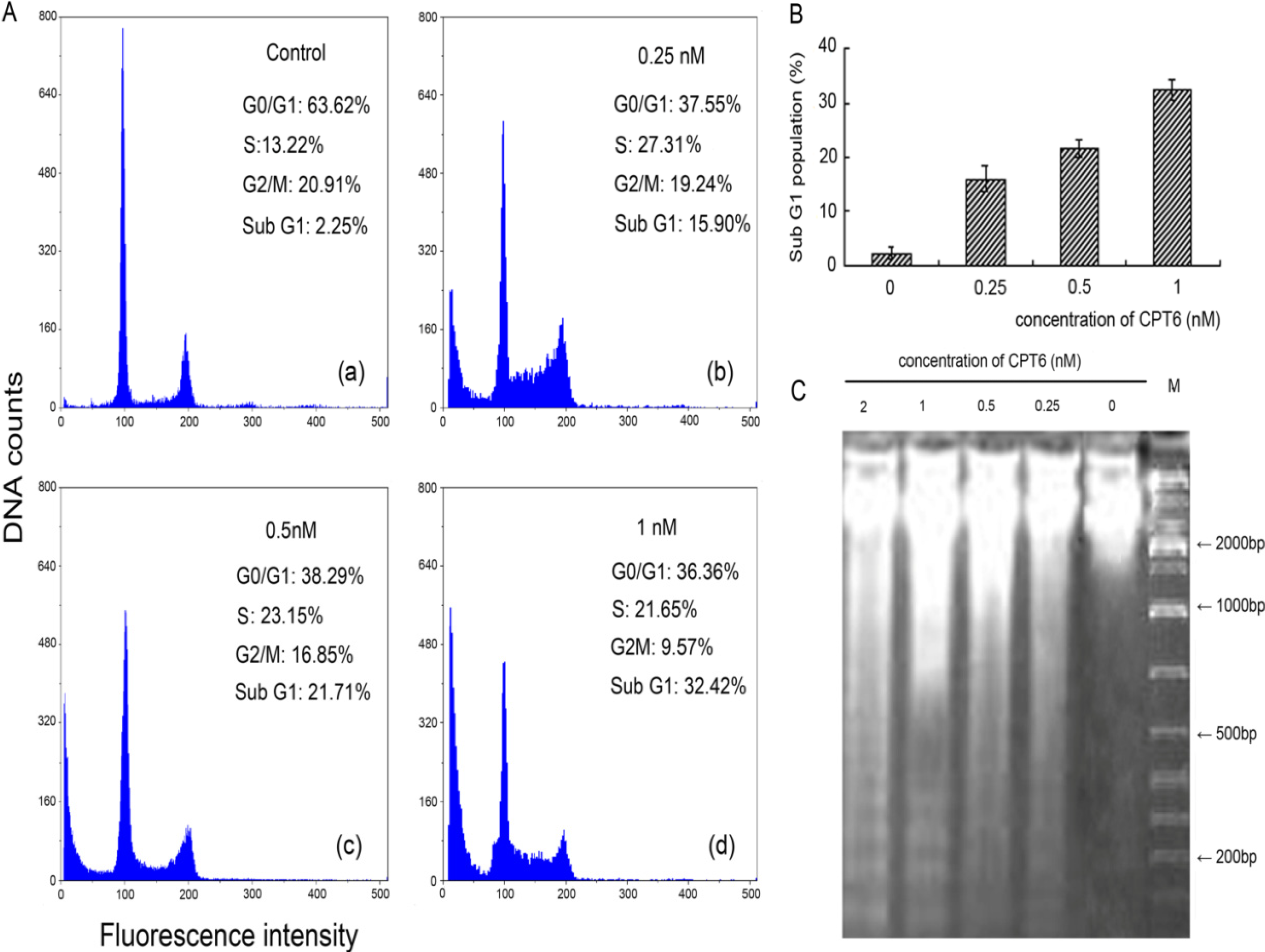{kind=link}
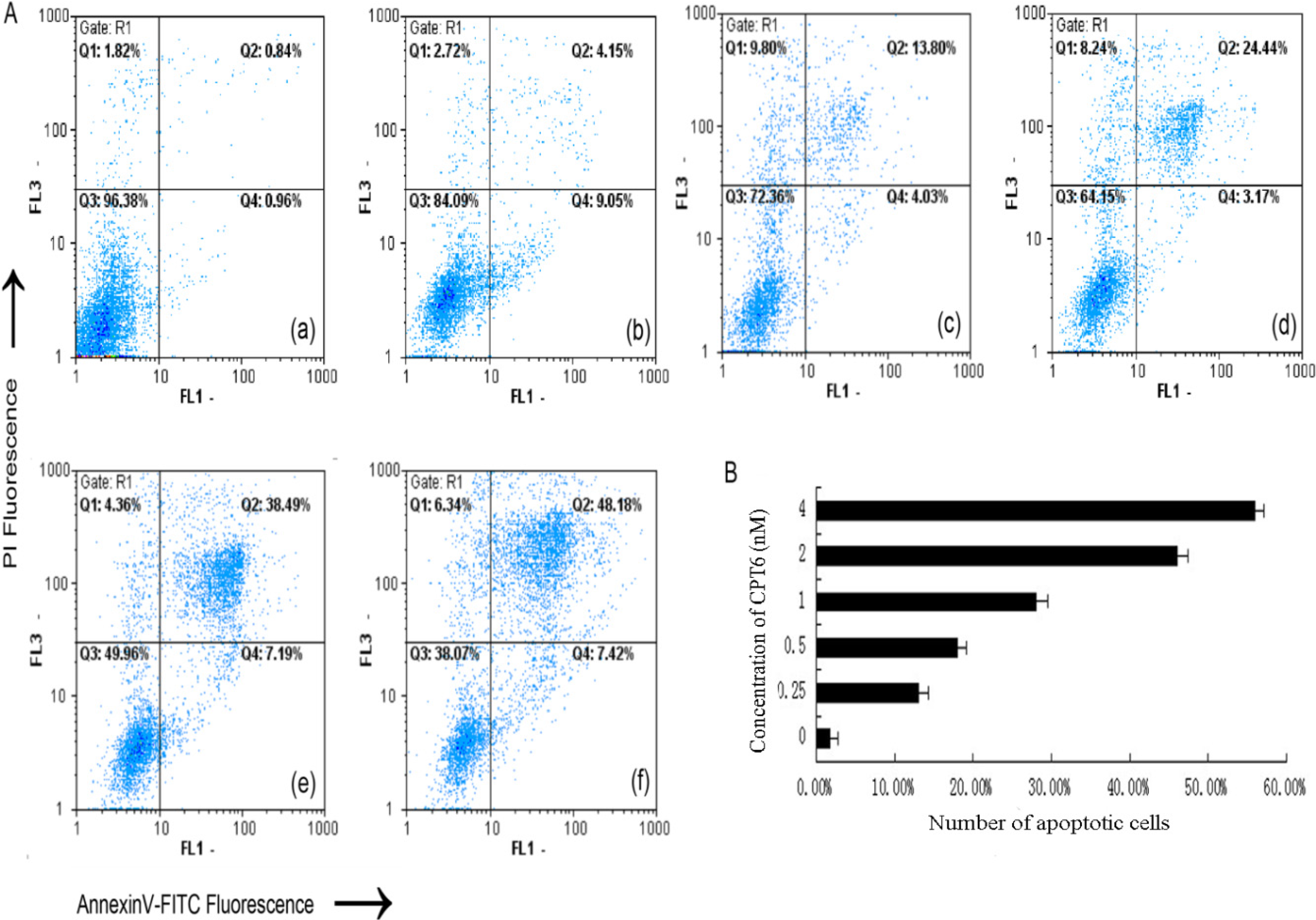{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CPT6 (nM/mL) | Inhibition (%) ± SD | ||
|---|---|---|---|
| 24 h | 48 h | 72 h | |
| 0.125 | 8.55 ± 2.68 * | 6.96 ± 2.44 * | 10.77 ± 1.48 ** |
| 0.25 | 14.09 ± 3.05 | 18.02 ± 3.11 | 32.88 ± 3.07 |
| 0.5 | 27.10 ± 2.69 * | 30.82 ± 3.59 | 52.83 ± 4.01 |
| 1 | 36.69 ± 3.29 | 43.56 ± 2.59 * | 64.31 ± 3.13 |
| 2 | 48.53 ± 3.62 | 53.73 ± 3.15 | 73.94 ± 3.82 |
| 4 | 56.80 ± 2.42 * | 67.36 ± 3.22 | 80.63 ± 4.16 |
2.2. Analysis of Cell Cycle Distribution

2.3. DNA Damage
2.4. Apoptosis/Necrosis Analysis

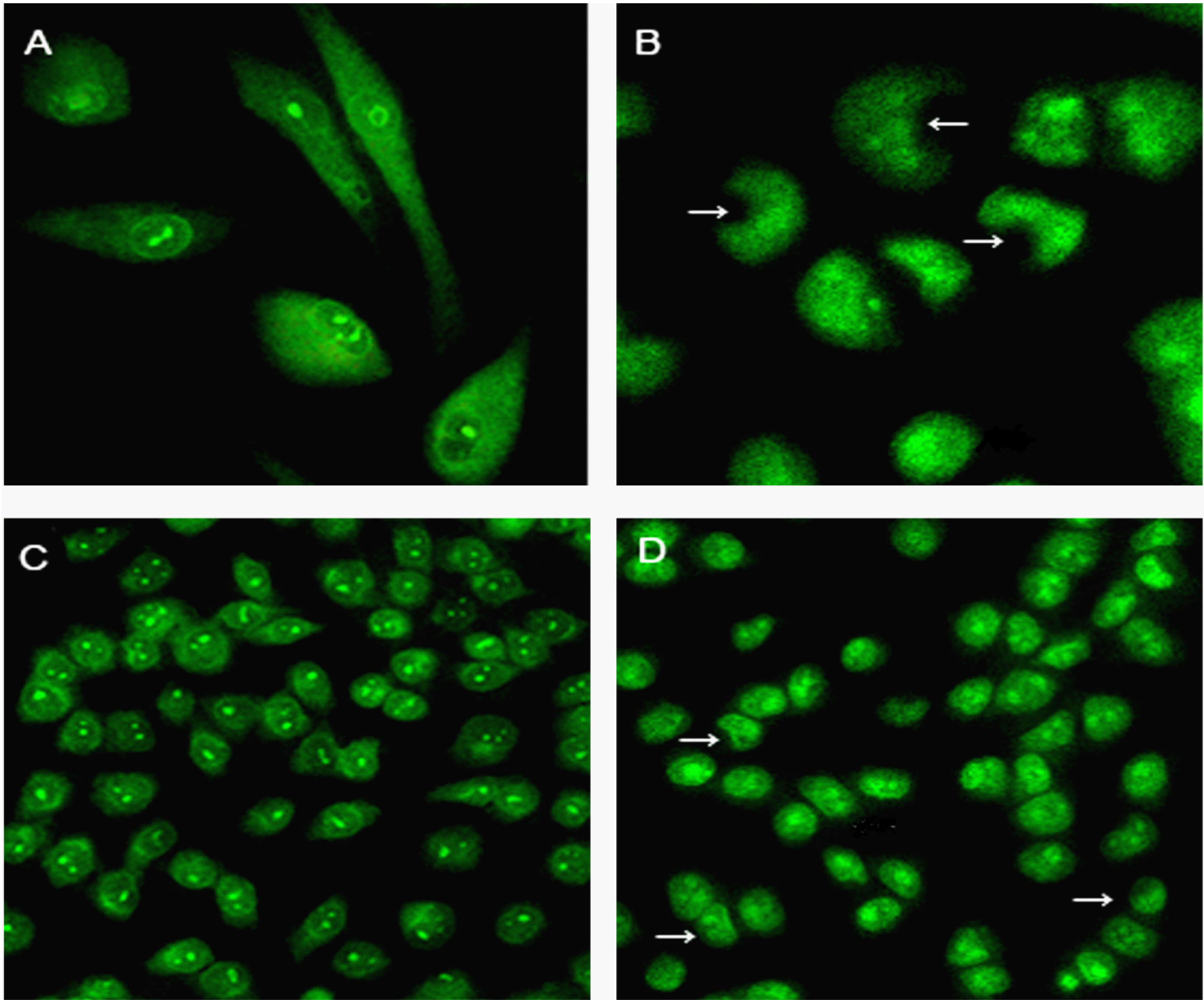
2.5. Changes in Nuclear Morphology and Chromatin Condensation

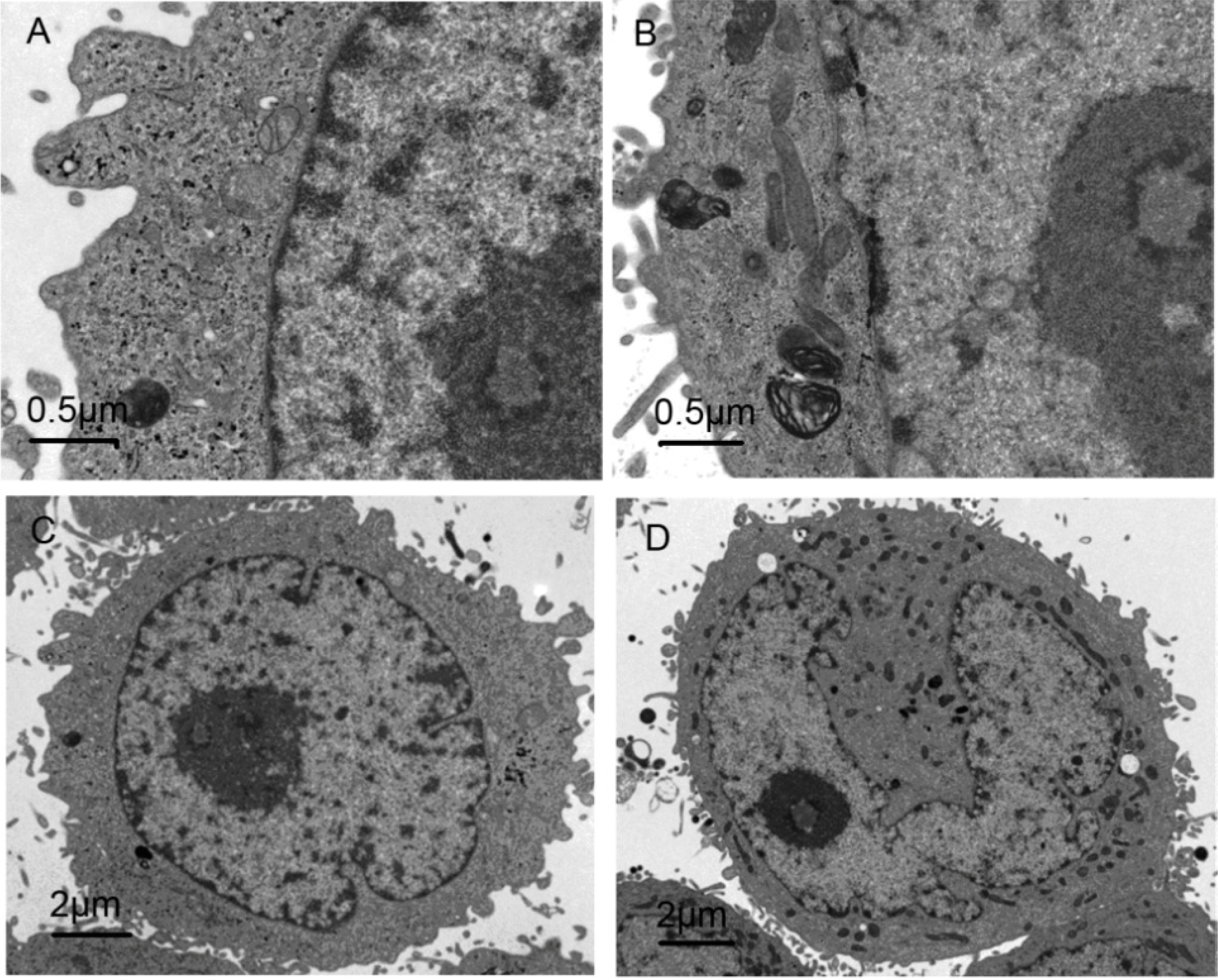
2.6. Morphological Observation of Cell Ultrastructure by TEM

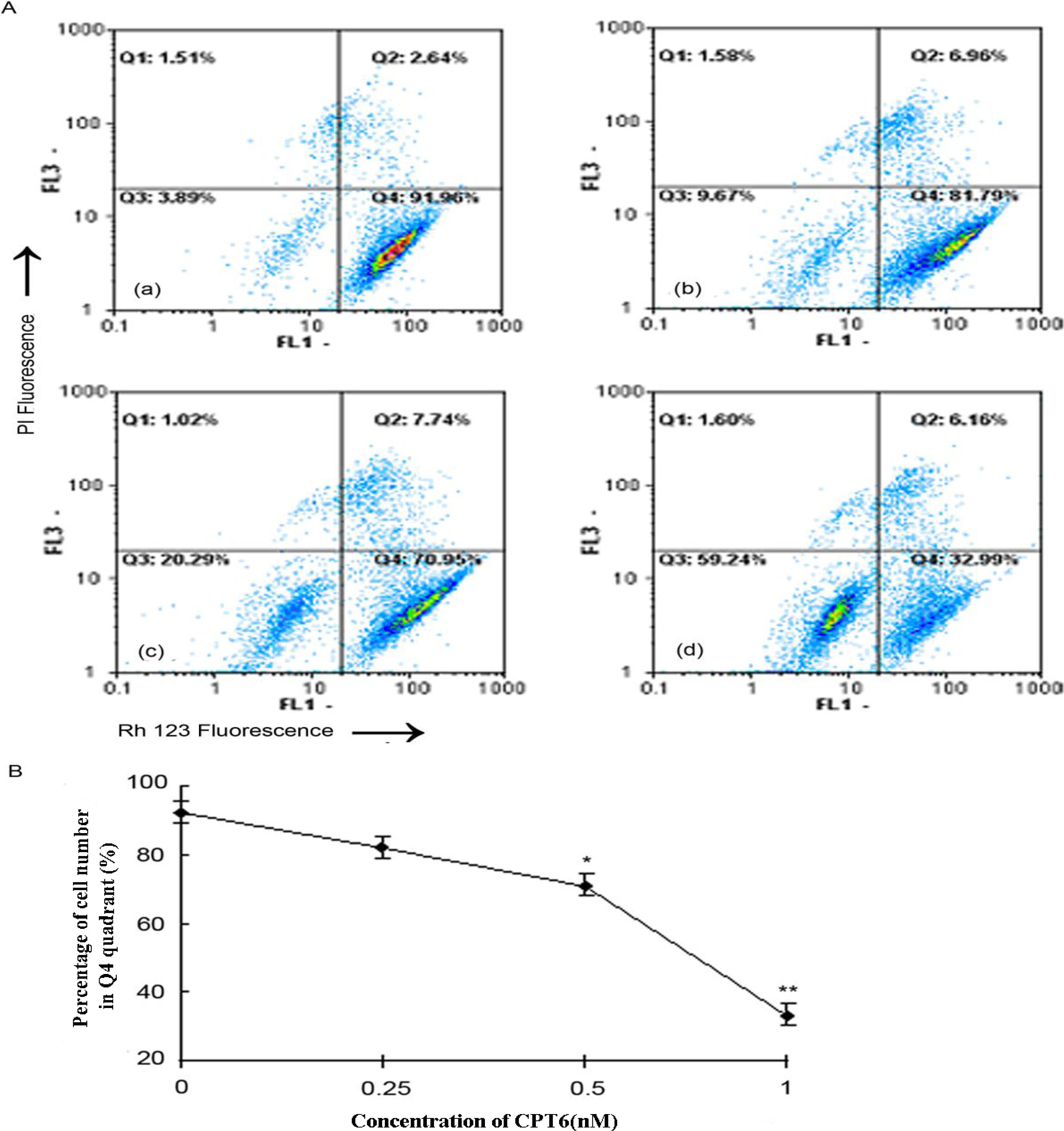
2.7. CPT6-Induced Loss of ΔΨ m in MCF-7 Cells
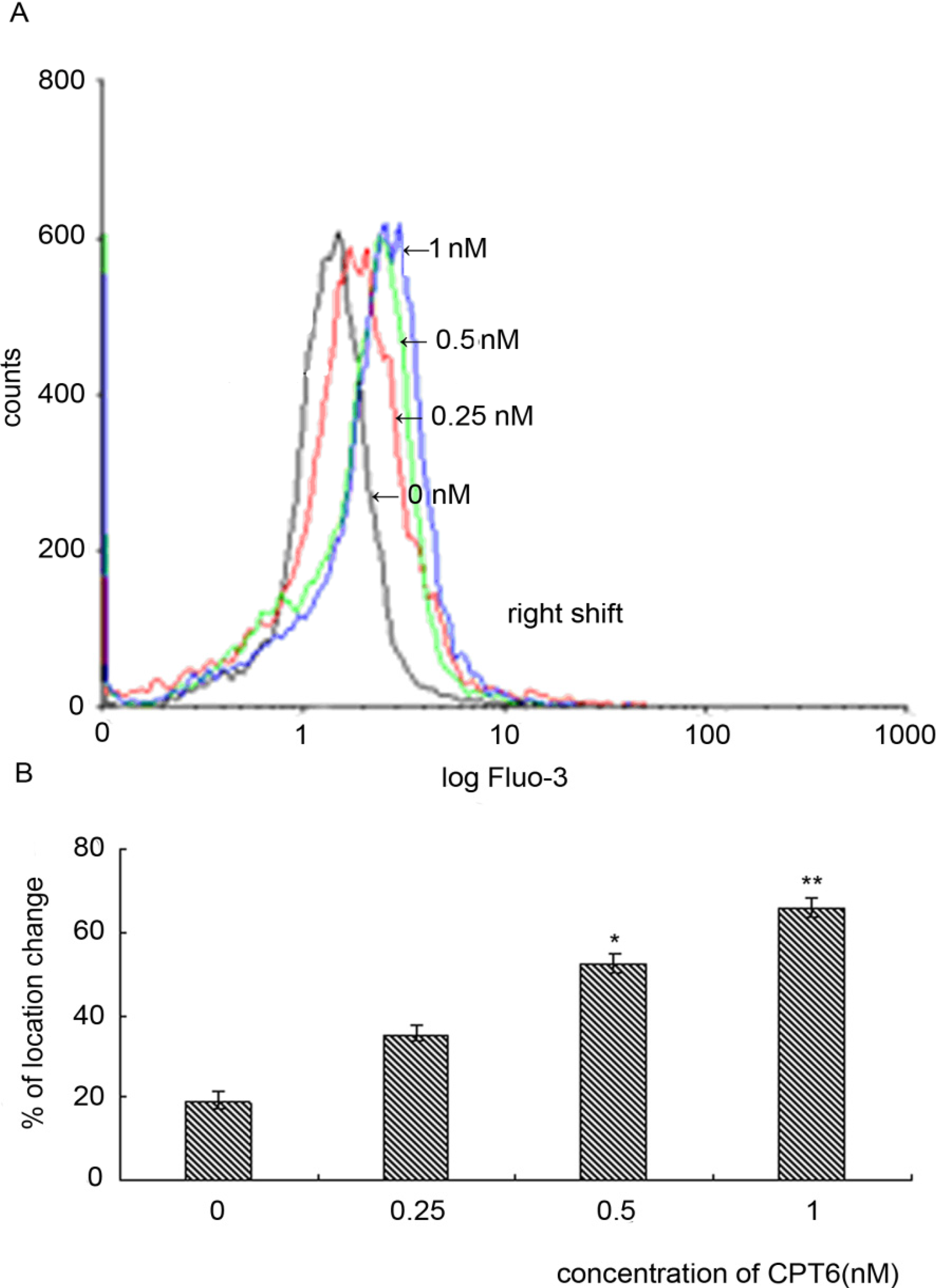
2.8. Effect of CPT6 on Intracellular Calcium in MCF-7 Cells


3. Experimental
3.1. Materials
3.2. Cell Line and Culture
3.3. Cytotoxicity and Antiproliferative Activities
3.4. Cell Cycle Distribution
3.5. Detection of DNA Fragmentation
3.6. Apoptosis Assays
3.7. Morphological Observations
3.8. Assessment of Mitochondrial Membrane Potential (ΔΨ m)
3.9. Determination of the Concentration of Calcium in Cells
3.10. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Verschraegen, C.F.; Jaeckle, K. Alternative administration of camptothecin analogues. Ann. N.Y. Acad. Sci. 2000, 922, 237–246. [Google Scholar]
- Giovanell, B.C.; Stehlin, J.S. DNA-topoisomerase I-targeted chemotherapy of human colon cancer in xenografts. Science 1989, 246, 1046–1048. [Google Scholar]
- Hsiang, Y.H.; Hertzberg, R. Camptothecin induces protein-linked DNA breaks via mammalian DNA topoisomerase I. J. Biol. Chem. 1985, 260, 14873–14878. [Google Scholar]
- Bom, D.; Curran, D.P. The novel silatecan 7-tert-butyldimethylsilyl-10-hydroxycamptothecin displays high lipophilicity, improved human blood stability, and potent anticancer activity. J. Med. Chem. 2000, 43, 3970–3980. [Google Scholar] [CrossRef]
- Van Hattum, A.H.; Pinedo, H.M. New highly lipophilic camptothecin BNP1350 is an effective drug in experimental human cancer. Int. J. Cancer. 2000, 88, 260–266. [Google Scholar] [CrossRef]
- Li, Q.Y.; Zu, Y.G.; Shi, R.Z. Synthesis and antitumor activity of novel 10-substituted camptothecin analogues. Bioorg. Med. Chem. 2006, 14, 7175–7182. [Google Scholar] [CrossRef]
- Li, Q.Y.; Lv, H.Y. Synthesis and antitumor activity of novel 20s-camptothecin analogues. Bioorg. Med. Chem. Lett. 2009, 19, 513–515. [Google Scholar] [CrossRef]
- Bhutia, S.K.; Mallick, S.K. Abrus abrin derived peptides induce apoptosis by targeting mitochondria in HeLa cells. Cell Biol. International 2009, 33, 720–727. [Google Scholar]
- Blagosklonny, M.V.; Pardee, A.B. Exploiting cancer cell cycling for selective protection of normal cells. Cancer Res. 2002, 61, 4301–4305. [Google Scholar]
- Giri, B.; Gomes, A. Antiproliferative, cytotoxic and apoptogenic activity of Indian toad (Bufo melanostictus, Schneider) skin extract on U937 and K562 cells. Toxicon 2006, 48, 388–400. [Google Scholar] [CrossRef]
- Liu, W.; Zhu, Y.S. Therapeutic efficacy of NSC606985, a novel camptothecin analog, in a mouse model of acute promyelocytic leukemia. Leukemia Res. 2007, 31, 1565–1574. [Google Scholar] [CrossRef]
- Croce, A.C.; Bottiroli, G. Subcellular localization of the camptothecin analogues, topotecan and gimatecan. Biochem. Pharm. 2004, 67, 1035–1045. [Google Scholar] [CrossRef]
- Stevenson, D.; Wokosin, D. Measurement of the intracellular distribution of reduced glutathione in cultured rat hepatocytes using monochlorobimane and confocallaser scanning microscopy. Toxicol. In Vitro. 2002, 16, 609–619. [Google Scholar] [CrossRef]
- Jiang, S.G.; Zu, Y.G. Activation of the mitochondria-driven pathway of apoptosis in human PC-3 prostate cancer cells by a novel hydrophilic paclitaxel derivative, 7-xylosyl-10-deacetylpaclitaxel. Int. J. Oncol. 2008, 33, 103–111. [Google Scholar] [CrossRef]
- Xiao, X.J.; Huang, G.Q. Morphological study on apoptosis Hela cells induced by soyasaponins. Toxicol. in Vitro 2007, 21, 820–826. [Google Scholar] [CrossRef]
- Xiao, J.X.; Huang, G.Q. Soyasaponins inhibit the proliferation of Hela cells by inducing apoptosis. Exp. Toxicol. Pathol. 2007, 59, 35–42. [Google Scholar] [CrossRef]
- Huang, M.; Gao, H.Y. Chimmitecan, a novel 9-substituted Camptothecin, with improved anticancer pharmacologic profiles in vitro and in vivo. Clin. Cancer Res. 2007, 13, 1298–1307. [Google Scholar] [CrossRef]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef]
- Vermeulen, K.; Van Bockstaele, D.R. Apoptosis: Mechanisms and relevance in cancer. Ann. Hematol. 2005, 84, 627–639. [Google Scholar] [CrossRef]
- Frankfurt, O.S.; Krishan, A. Apoptosis-based drug screening and detection of selective toxicity to cancer cells. Anticancer Drugs 2003, 14, 555–561. [Google Scholar] [CrossRef]
- Liu, L.F. DNA topoisomerase poisons as antitumor drugs. Annu. Rev. Biochem. 1989, 58, 351–375. [Google Scholar] [CrossRef]
- D’Arpa, P.; Beardmore, C. Involvement of nucleic acid synthesis in cell killing mechanisms of topoisomerase poisons. Cancer Res. 1990, 50, 6919–24. [Google Scholar]
- Green, D.R. Apoptotic pathways: The roads to run. Cell 1998, 94, 695–698. [Google Scholar] [CrossRef]
- Kastan, M.B.; Bartek, J. Cell-cycle checkpoints and cancer. Nature 2004, 432, 316–323. [Google Scholar] [CrossRef]
- Kang, G.Y.; Lee, E.R. Downregulation of PLK-1 expression in kaempferol-induced apoptosis of MCF-7 cells. Eur. J. Pharmacol. 2009, 611, 17–21. [Google Scholar] [CrossRef]
- Vermes, I.; Haanen, C. A novel assay for apoptosis, Flow cytometric detection of phosphatidylserin expression on early apoptotic cells using fluorescein labeled annexin-V. J. Immunol. Meth. 1995, 184, 39–51. [Google Scholar] [CrossRef]
- Fu, Y.R.; Yi, Z.J. Hydroxycamptothecin-induced apoptosis in hepatoma SMMC-7721 cells and the role of mitochondrial pathway. Mitochondrion 2006, 6, 211–217. [Google Scholar] [CrossRef]
- Han, J.; Goldstein, L.A. Interrelated roles for Mcl-1 and BIM in regulation of TRAIL-mediated mitochondrial apoptosis. J. Biol. Chem. 2006, 281, 10153–10163. [Google Scholar]
- Yoneyama, K. Validation of confocal laser scanning microscopy for detecting intracellular calcium heterogeneity in live slices. J. Pharmacol. Toxicol. Methods 2001, 45, 187–193. [Google Scholar]
- Sample Availability: Samples of the compounds are available from the authors.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Chu, C.; Xu, J.; Cheng, D.; Li, X.; Tong, S.; Yan, J.; Li, Q. Anti-Proliferative and Apoptosis-Inducing Effects of Camptothecin-20(s)-O-(2-pyrazolyl-1)acetic Ester in Human Breast Tumor MCF-7 Cells. Molecules 2014, 19, 4941-4955. https://doi.org/10.3390/molecules19044941
Chu C, Xu J, Cheng D, Li X, Tong S, Yan J, Li Q. Anti-Proliferative and Apoptosis-Inducing Effects of Camptothecin-20(s)-O-(2-pyrazolyl-1)acetic Ester in Human Breast Tumor MCF-7 Cells. Molecules. 2014; 19(4):4941-4955. https://doi.org/10.3390/molecules19044941
Chicago/Turabian StyleChu, Chu, Jialin Xu, Dongping Cheng, Xingnuo Li, Shengqiang Tong, Jizong Yan, and Qingyong Li. 2014. "Anti-Proliferative and Apoptosis-Inducing Effects of Camptothecin-20(s)-O-(2-pyrazolyl-1)acetic Ester in Human Breast Tumor MCF-7 Cells" Molecules 19, no. 4: 4941-4955. https://doi.org/10.3390/molecules19044941
APA StyleChu, C., Xu, J., Cheng, D., Li, X., Tong, S., Yan, J., & Li, Q. (2014). Anti-Proliferative and Apoptosis-Inducing Effects of Camptothecin-20(s)-O-(2-pyrazolyl-1)acetic Ester in Human Breast Tumor MCF-7 Cells. Molecules, 19(4), 4941-4955. https://doi.org/10.3390/molecules19044941