Immunostimulatory Effects of Polysaccharides Isolated from Makgeolli (Traditional Korean Rice Wine)

Abstract
:1. Introduction
2. Results and Discussion
2.1. The Chemical Properties of PSM

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PSM | ||
|---|---|---|
| Yield (g/L) | 0.33 | |
| Chemical composition (%) | Neutral sugar | 87.3 ± 0.0 |
| Uronic acid | 2.5 ± 0.0 | |
| Protein | 10.2 ± 0.0 | |
| Composition of neutral sugar a (mole%) b | Glucose | 66.4 ± 0.2 |
| Mannose | 24.5 ± 0.1 | |
| Galactose | 6.6 ± 0.0 | |
| Xylose | 1.5 ± 0.0 | |
| Arabinose | 1.0 ± 0.0 | |
2.2. Effects of PSM on Immune Functions in Normal Mice
2.2.1. Effects on Body and Organ Weights
| Group | Number of animals | Initial body weight (g) | Final body weight (g) | Weight growth rate (%) | Spleen Index (SI) a | Thymus Index (TI) b |
|---|---|---|---|---|---|---|
| Control | 10 | 22.00 ± 1.79 | 35.50 ± 4.38 | 61.36 | 0.65 ± 0.08 | 0.35 ± 0.13 |
| CVT c 200 mg/kg | 10 | 21.45 ± 1.90 | 34.66 ± 2.65 | 61.59 | 0.77 ± 0.09 * | 0.42 ± 0.11 |
| PSM 100 mg/kg | 10 | 21.60 ± 1.90 | 34.88 ± 5.33 | 61.48 | 0.68 ± 0.10 | 0.36 ± 0.11 |
| PSM 200 mg/kg | 10 | 21.60 ± 1.84 | 34.86 ± 3.44 | 61.39 | 0.76 ± 0.13 * | 0.40 ± 0.12 |
2.2.2. Effects on Lymphocytes Proliferation
2.3. Effects of PSM on Immune Functions in CY-induced Immunosuppressed Mice
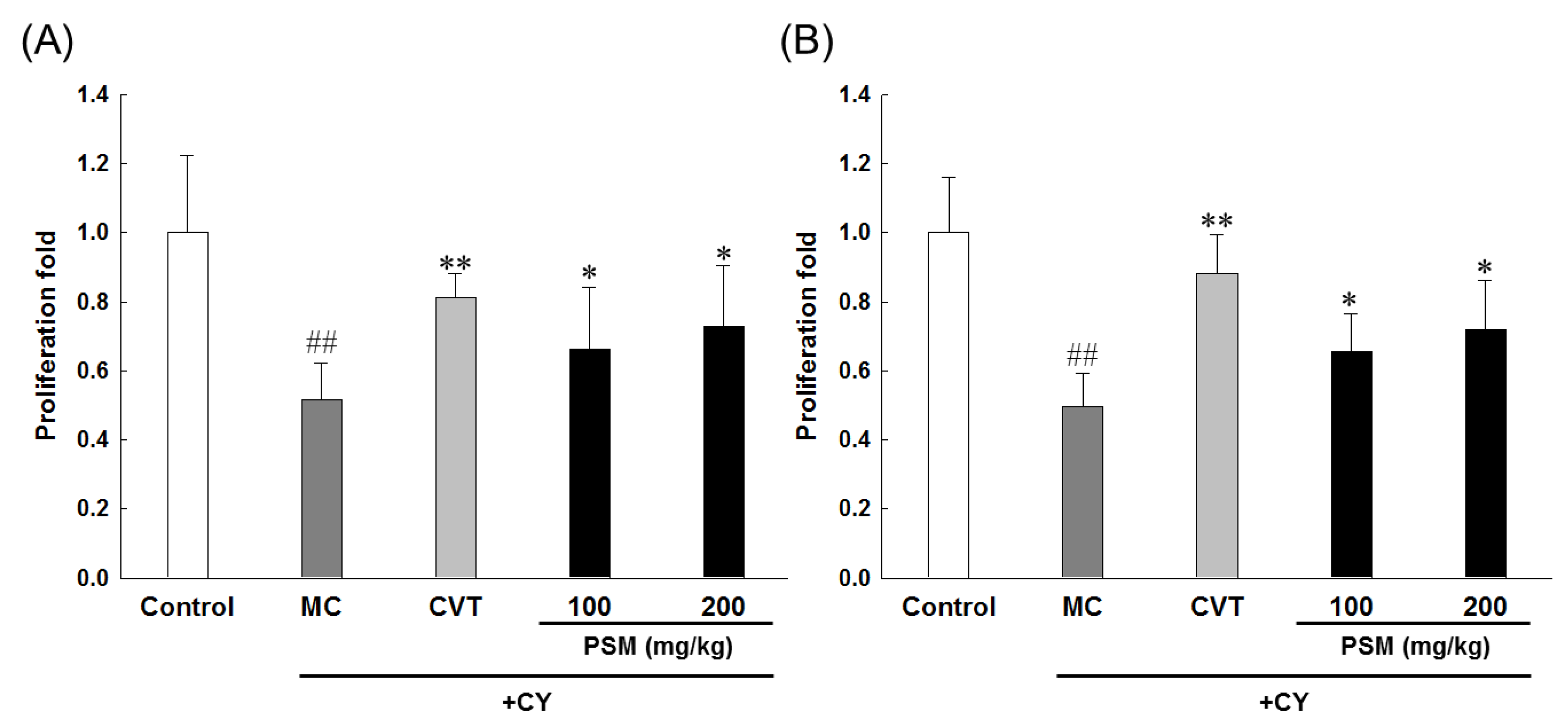
2.3.1. Effects on Lymphocytes Proliferation


2.3.2. Effects on NK Cell Activity

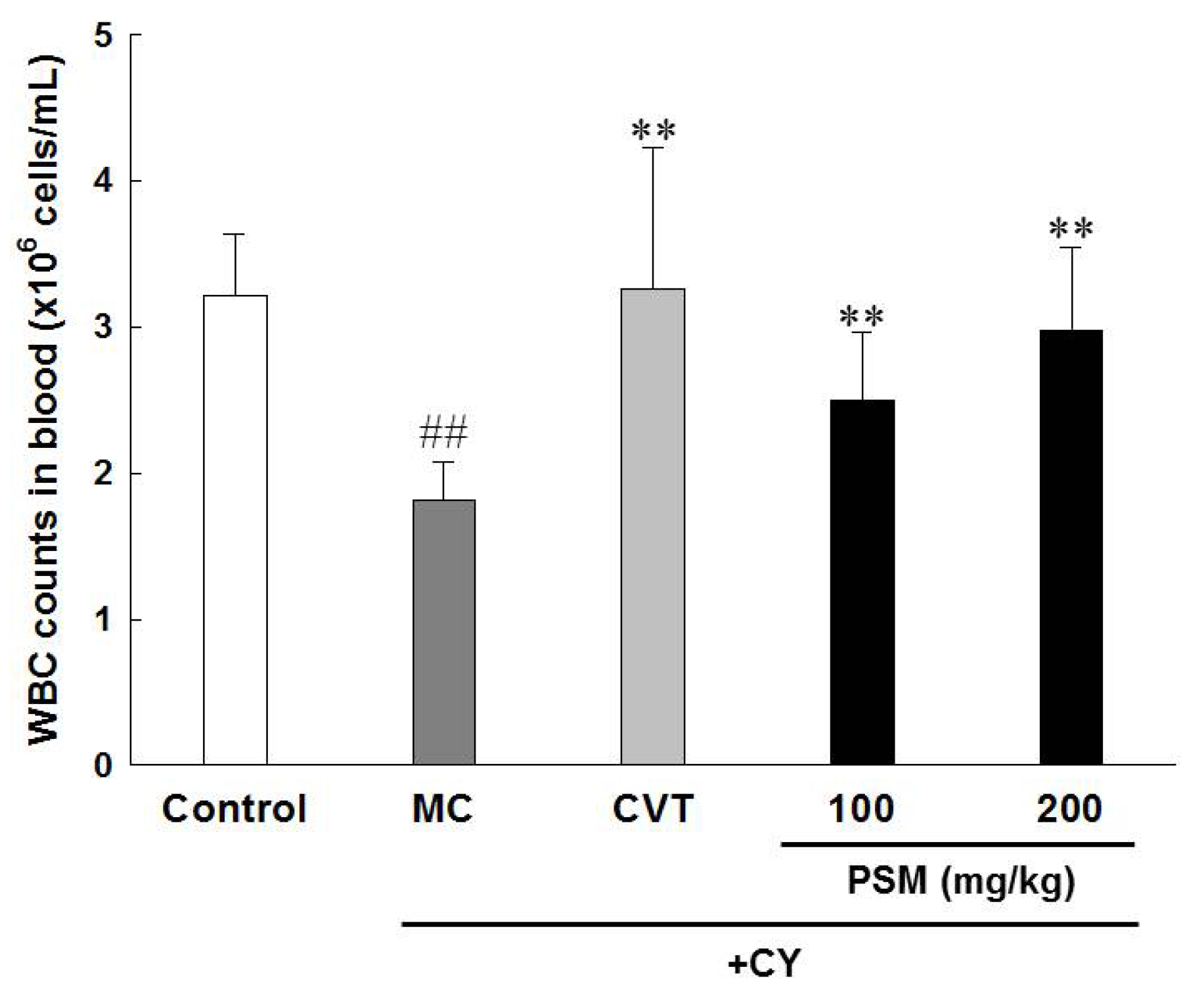
2.3.3. Effects on White Blood Cell (WBC) Count

3. Experimental
3.1. Materials
3.2. Preparation of PSM
3.3. Characterization of PSM
3.4. Animals
3.5. Animals Experiments
3.6. Splenocyte Proliferation Assay
3.7. NK Cell-Mediated Cytotoxicity Assay
3.8. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ha, J.; Wang, Y.; Jang, H.; Seog, H.; Chen, X. Determination of E,E-farnesol in Makgeolli (rice wine) using dynamic headspace sampling and stir bar sorptive extraction coupled with gas chromatography-mass spectrometry. Food Chem. 2014, 142, 79–86. [Google Scholar] [CrossRef]
- Kang, B.S.; Lee, J.E.; Park, H.J. Electronic tongue-based discrimination of Korean rice wines (makgeolli) including prediction of sensory evaluation and instrumental measurements. Food Chem. 2014, 151, 317–323. [Google Scholar] [CrossRef]
- Shin, M.O.; Kang, D.Y.; Kim, M.H.; Bae, S.J. Effect of growth inhibition and quinone reductase activity stimulation of makgeolli fractions in various cancer cells. J. Korean Soc. Food Sci. Nutr. 2008, 37, 288–293. [Google Scholar] [CrossRef]
- Lee, S.J.; Kim, G.W. Hypolipidemic effect of hexane fraction from Rhizopus oryzae KSD-815 through peroxisome proliferator-activated receptor-α. J. Korean Soc. Appl. Biol. Chem. 2010, 53, 761–765. [Google Scholar] [CrossRef]
- Lee, S.J.; Kim, J.H.; Jung, Y.W.; Park, S.Y.; Shin, W.C.; Park, C.S.; Hong, S.Y.; Kim, G.W. Compositions of organic acids and physiological functionality of commercial makgeolli. Korean J. Food Sci. Technol. 2011, 43, 206–212. [Google Scholar]
- Shin, M.O.; Kim, M.H.; Bae, S.J. The effect of makgeolli on blood flow, serum lipid improvement and inhibition of ACE in vitro. J. life Sci. 2010, 20, 710–716. [Google Scholar] [CrossRef]
- Park, S.Y.; Lee, J.; Yu, K.W.; Han, N.S.; Lee, H.; Koh, J.H.; Shin, K.S. Anti-complementary polysaccharides in grape wines. J. Korean Soc. Food Sci. Nutr. 2006, 35, 1232–1236. [Google Scholar] [CrossRef]
- Hwang, Y.C.; Shin, K.S. Characterization of immune-stimulating polysaccharides isolated from Korean persimmon vinegar. Korean J. Food Sci. Technol. 2008, 40, 220–227. [Google Scholar]
- Park, H.R.; Lee, M.S.; Jo, S.Y.; Won, H.J.; Lee, H.S.; Lee, H.; Shin, K.S. Immuno-stimulating activities of polysaccharides isolated from commercial soy sauce and traditional Korean soy sauce. Korean J. Food Sci. Technol. 2012, 44, 228–234. [Google Scholar] [CrossRef]
- Bae, S.H.; Jung, E.Y.; Kim, S.Y.; Shin, K.S.; Suh, H.J. Antioxidant and immune-modulating activities of Korean traditional rice wine, takju. J. Food Biochem. 2010, 34, 233–248. [Google Scholar] [CrossRef]
- Bae, S.H.; Choi, J.W.; Ra, K.S.; Yu, K.W.; Shin, K.S.; Park, S.S.; Suh, H.J. Anti-complementary activity of enzyme-treated traditional Korean rice wine (Makgeolli) hydrolysates. J. Sci. Food Agric. 2012, 92, 1765–1770. [Google Scholar] [CrossRef]
- Hong, H.D.; Cho, C.W.; Rhee, Y.K.; Choi, H.D.; Lee, H.S. Status on technology development using immune-modulating polysaccharide. Food Sci. Ind. 2012, 45, 1–11. [Google Scholar]
- Jiang, M.H.; Zhu, L.; Jiang, J.G. Immunoregulatory actions of polysaccharides from Chinese herbal medicine. Expert Opin. Ther. Targets 2010, 14, 1367–1402. [Google Scholar] [CrossRef]
- Zhao, C.; Li, M.; Luo, Y.; Wu, W. Isolation and structural characterization of an immunostimulating polysaccharide from fuzi, Aconitum carmichaeli. Carbohydr. Res. 2006, 341, 485–491. [Google Scholar] [CrossRef]
- Chen, Y.; Tang, J.; Wang, X.; Sun, F.; Liang, S. An immunostimulatory polysaccharide (SCP-IIa) from the fruit of Schisandra chinensis (Turcz.) Baill. Int. J. Biol. Macromol. 2012, 50, 844–848. [Google Scholar] [CrossRef]
- Guo, L.; Liu, J.; Hu, Y.; Wang, D.; Li, Z.; Zhang, J.; Qin, T.; Liu, X.; Liu, C.; Zhao, X.; et al. Astragalus polysaccharide and sulphated epimedium polysaccharide synergistically resist the immunosuppression. Carbohydr. Polym. 2012, 90, 1055–1060. [Google Scholar] [CrossRef]
- O’Neill, M.; Albersheim, P.; Darvill, A. The pectic polysaccharides of primary cell walls. In Methods in Plant Biochemistry; Dey, P.M., Ed.; Academic: London, UK, 1990; Volume 2, pp. 415–441. [Google Scholar]
- Engelsen, S.B.; Cros, S.; Mackie, W.; Perez, S. A molecular builder for carbohydrates: Application to polysaccharides and complex carbohydrates. Biopolymers 1996, 39, 417–433. [Google Scholar] [CrossRef]
- Klis, F.M.; Mol, P.; Hellingwerf, K.; Brul, S. Dynamics of cell wall structure in Saccharomyces cerevisiae. FEMS Microbiol. Rev. 2002, 26, 239–256. [Google Scholar] [CrossRef]
- Biondo, P.D.; Goruk, S.; Ruth, M.R.; O’Connell, E.; Field, C.J. Effect of CVT-E002™ (COLD-fX®) versus a ginsenoside extract on systemic and gut-associated immune function. Int. Immunopharmacol. 2008, 8, 1134–1142. [Google Scholar] [CrossRef]
- Chen, X.; Nie, W.; Fan, S.; Zhang, J.; Wang, Y.; Lu, J.; Jin, L. A polysaccharide from Sargassum fusiforme protects against immunosuppression in cyclophosphamide-treated mice. Carbohydr. Polym. 2012, 90, 1114–1119. [Google Scholar] [CrossRef]
- Cerqueira, F.; Cordeiro-Da-Silva, A.; Gaspar-Marques, C.; Simões, F.; Pinto, M.M.; Nascimento, M.S. Effect of abietane diterpenes from Plectranthus grandidentatus on T- and B-lymphocyte proliferation. Bioorg. Med. Chem. 2004, 12, 217–223. [Google Scholar]
- Lee, Y.S.; Lee, G.H.; Kwon, Y.K.; Park, J.H.; Shin, S.W. Water extracted evodiae fructus possessed immunomodulatory activities on cyclophosphamide induced immunosuppression. Korean J. Ori. Med. Physiol. Pathol. 2007, 21, 485–490. [Google Scholar]
- Artym, J.; Zimecki, M.; Kruzel, M.L. Normalization of peripheral blood cell composition by lactoferrin in cyclophosphamide-treated mice. Med. Sci. Monit. 2004, 10, 84–89. [Google Scholar]
- Moynihan, J.; Cohen, N. The kinetics of recovery of leukocyte number and lymphocyte function following an injection of a single high dose of cyclophosphamide in C3H/HeJ mice. Int. J. Immunopharmacol. 1989, 11, 517–527. [Google Scholar] [CrossRef]
- Vivier, E.; Nunes, J.A.; Vely, F. Natural killer cell signalling pathways. Science 2004, 306, 1517–1519. [Google Scholar] [CrossRef]
- Kantakamalakul, W.; Jaroenpool, J.; Pattanapanyasat, K. A novel enhanced green fluorescent protein (EGFP)-K562 flow cytometric method for measuring natural killer (NK) cell cytotoxic activity. J. Immunol. Methods 2003, 272, 189–197. [Google Scholar] [CrossRef]
- Shalit, I.; Kletter, Y.; Halperin, D.; Waldman, D.; Vasserman, E.; Nagler, A.; Fabian, I. Immunomodulatory effects of moxifloxacin in comparison to ciprofloxacin and G-CSF in a murine model of cyclophosphamide-induced leukopenia. Eur. J. Haematol. 2001, 66, 287–296. [Google Scholar] [CrossRef]
- Zhou, H.; Sun, F.; Li, H.; Zhang, S.; Liu, Z.; Pei, J.; Liang, C. Effect of recombinant Ganoderma lucidum immunoregulatory protein on cyclophosphamide-induced leukopenia in mice. Immunopharmacol. Immunutoxicol. 2013, 35, 426–433. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Robers, P.A.; Smith, F. Colorimetric method for determination of sugar and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar]
- Bitter, T.; Muir, H.M. A modified uronic acid carbazole reaction. Anal. Biochem. 1962, 4, 330–334. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Jones, T.M.; Albersheim, P. A gas chromatographic method of the determination of aldose and uronic acid constituents of plant cell wall polysaccharides. Plant Physiol. 1972, 49, 926–936. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the makgeolli polysaccharides are available from the authors.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Cho, C.-W.; Han, C.-j.; Rhee, Y.K.; Lee, Y.-C.; Shin, K.-S.; Hong, H.-D. Immunostimulatory Effects of Polysaccharides Isolated from Makgeolli (Traditional Korean Rice Wine). Molecules 2014, 19, 5266-5277. https://doi.org/10.3390/molecules19045266
Cho C-W, Han C-j, Rhee YK, Lee Y-C, Shin K-S, Hong H-D. Immunostimulatory Effects of Polysaccharides Isolated from Makgeolli (Traditional Korean Rice Wine). Molecules. 2014; 19(4):5266-5277. https://doi.org/10.3390/molecules19045266
Chicago/Turabian StyleCho, Chang-Won, Chun-ji Han, Young Kyoung Rhee, Young-Chul Lee, Kwang-Soon Shin, and Hee-Do Hong. 2014. "Immunostimulatory Effects of Polysaccharides Isolated from Makgeolli (Traditional Korean Rice Wine)" Molecules 19, no. 4: 5266-5277. https://doi.org/10.3390/molecules19045266
APA StyleCho, C. -W., Han, C. -j., Rhee, Y. K., Lee, Y. -C., Shin, K. -S., & Hong, H. -D. (2014). Immunostimulatory Effects of Polysaccharides Isolated from Makgeolli (Traditional Korean Rice Wine). Molecules, 19(4), 5266-5277. https://doi.org/10.3390/molecules19045266