
Phytochemicals from Ruta graveolens Activate TAS2R Bitter Taste Receptors and TRP Channels Involved in Gustation and Nociception



Abstract
:1. Introduction
1.1. Taste Chemoreception and Bioactive Compounds
1.2. Ruta graveolens, Medicinal Uses and Taste
1.3. Aim of This Work

2. Results and Discussion
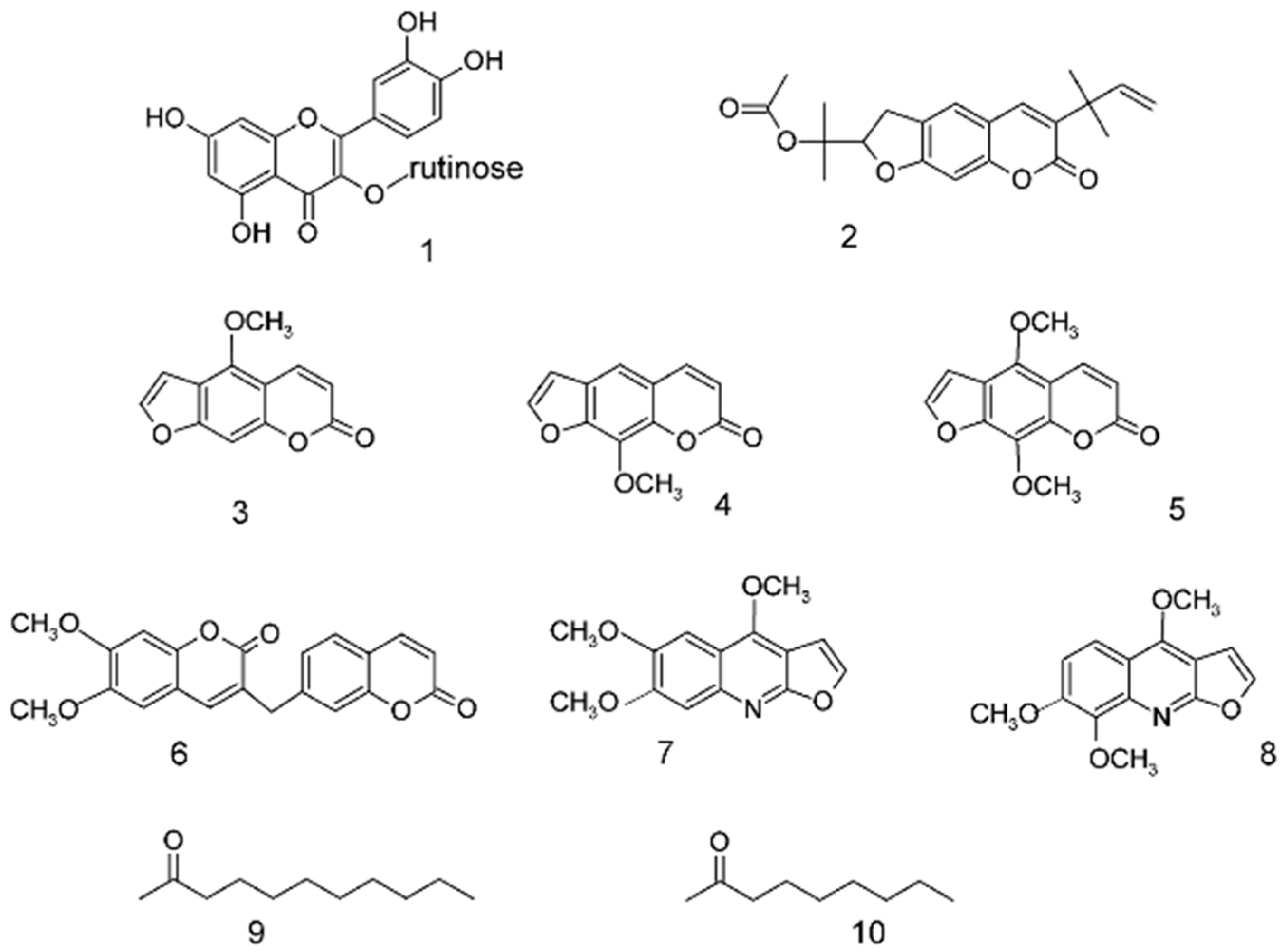
2.1. Isolation of Phytochemicals
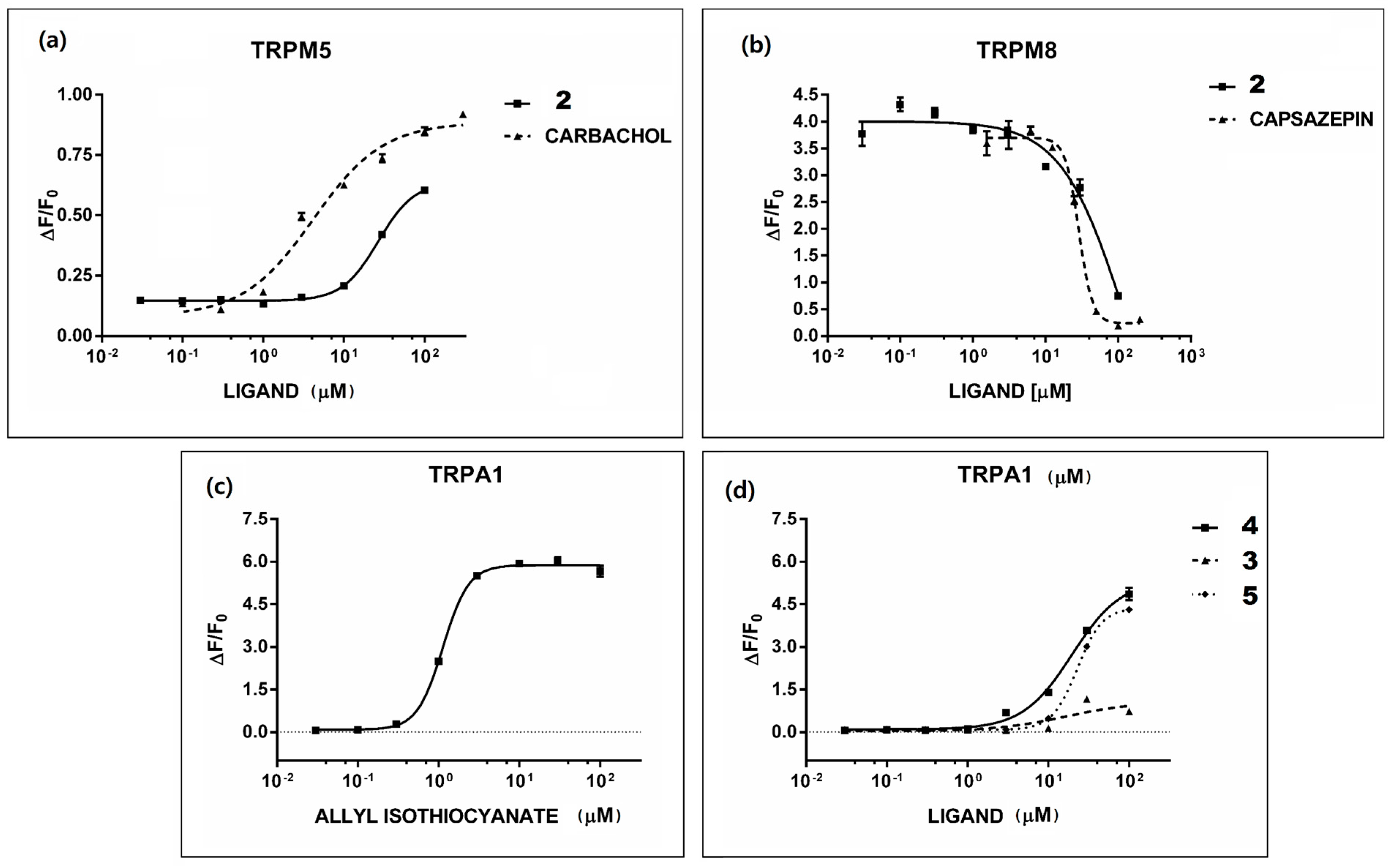
2.2. In Vitro Assays of Rue Phytochemicals on TAS2R and TRP Receptors

{kind=link}
{kind=link}
{kind=link}
| CPD | TAS2R Assays | TRP Assays | |||||
|---|---|---|---|---|---|---|---|
| TAS2R10 | TAS2R14 | TAS2R49 | TRPA1 | TRPM5 | TRPM8 | TRPV1 | |
| 1 | − | − | − | − | − | − | − |
| 2 | − | − | − | − | + | § | + |
| 3 | +++ | − | − | − | − | − | − |
| 4 | + | + | ++ | + | − | − | − |
| 5 | ++ | + | − | + | − | − | − |
| 6 | − | − | − | − | − | − | − |
| 7 | − | − | − | − | − | − | − |
| 8 | − | + | − | − | − | − | − |
| 9 | − | − | − | − | − | − | − |
| 10 | − | − | − | − | − | − | − |
| EO | − | ++ | − | − | − | − | − |
| TAS2R Referenced Agonists | ||
|---|---|---|
| Receptor | Compound | EC50 |
| TAS2R10 | Denatonium Benzoate | 8.3 µM |
| TAS2R10 | 3 | 2.8 µM |
| TAS2R10 | 4 | ≥20.6 µM |
| TAS2R10 | 5 | 12 µM |
| TAS2R14 | Aristolochic Acid | 2.3 µM |
| TAS2R14 | 4 | ≥10.8 µM |
| TAS2R14 | 5 | 11.1 µM |
| TAS2R14 | 8 | ≥15.8 µM |
| TAS2R14 | EO | 3.9 mg/L |
| TAS2R49 | Ritanserin | 6 µM |
| TAS2R49 | 4 | 12 µM |
| TRPA1 | Allyl Isothiocyanate | 1.1 µM |
| TRPA1 | 3 | n.d. |
| TRPA1 | 4 | ≥19.7 µM |
| TRPA1 | 5 | ≥22.8 µM |
| TRPM5 | Carbachol | 4.3 µM |
| TRPM5 | 2 | ≥26.7 µM |
| TRPM8 | WS3 | 4.0 µM |
2.3. Rue Phytochemicals and Bitter Taste TAS2 Receptors

2.4. Structure-Activity Relationship
2.5. Rue Phytochemicals and Somatosensory TRP Channels

3. Experimental Section
3.1. Plant Material
3.2. Extraction and Isolation
3.3. Spectroscopic Analysis of Compounds
3.4. In Vitro Assays with TAS2R and TRP Receptors
3.4.1. Compounds and References
3.4.2. Vectors and cDNAs
| TAS2R Referenced Agonists | ||
|---|---|---|
| Receptor | Compound | Concentration |
| TAS2R1 | Menthol | 300 µM |
| TAS2R3 | Chloroquine | 10 mM |
| TAS2R4 | Colchicine | 3 mM |
| TAS2R5 | Phenanthroline | 300 µM |
| TAS2R7 | Chloroquine | 10 mM |
| TAS2R8 | Chloramphenicol | 300 µM |
| TAS2R9 | Ofloxacin | 3 mM |
| TAS2R10 | Denatonium Benzoate | 300 µM |
| TAS2R13 | Denatonium Benzoate | 3 mM |
| TAS2R14 | Aristolochic Acid | 10 µM |
| TAS2R16 | Salicin | 3 mM |
| TAS2R38 | Phenylethyl Isothiocyanate | 300 µM |
| TAS2R39 | Denatonium Benzoate | 3 mM |
| TAS2R40 | Cinchinone | 100 µM |
| TAS2R43 | Aristolochic Acid | 1 µM |
| TAS2R44 | Aristolochic Acid | 10 µM |
| TAS2R46 | Strychnine | 10 µM |
| TAS2R47 | Denatonium Benzoate | 30 µM |
| TAS2R49 | Ritanserin | 100 µM |
| TAS2R50 | Andrographolide | 30 µM |
3.4.3. Cell Culture and Transfection
3.4.4. Calcium Imaging Analysis and Quantification
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Appendino, G.; Minassi, A.; Pagani, A.; Ech-Chahad, A. the role of natural products in the ligand deorphanization of TRP channels. Curr. Pharm. Des. 2008, 14, 2–17. [Google Scholar] [CrossRef] [PubMed]
- Roland, W.S.U.; van Buren, L.; Gruppen, H.; Driesse, M.; Gouka, R.J.; Smit, G.; Vincken, J.P. Bitter Taste receptor activation by flavonoids and isoflavonoids: modelled structural requirements for activation of hTAS2R14 and hTAS2R39. J. Agric. Food Chem. 2013, 61, 10454–10466. [Google Scholar] [CrossRef] [PubMed]
- Behrens, M.; Meyerhof, W.; Hellfritsch, C.; Hofmann, T. Sweet and umami taste: Natural products, their chemosensory targets, and beyond. Angew. Chem. Int. Ed. 2011, 50, 2220–2242. [Google Scholar] [CrossRef] [PubMed]
- Meyerhof, W.; Batram, C.; Kuhn, C.; Brockhoff, A.; Chudoba, E.; Bufe, B.; Appendino, G.; Behrens, M. The molecular receptive ranges of human TAS2R bitter taste receptors. Chem. Senses 2010, 35, 157–170. [Google Scholar] [CrossRef] [PubMed]
- Brockhoff, A.; Behrens, M.; Roudnitzky, N.; Appendino, G.; Avonto, C.; Meyerhof, W. Receptor agonism and antagonism of dietary bitter compounds. J. Neurosci. 2011, 31, 14775–14782. [Google Scholar] [CrossRef] [PubMed]
- Soares, S.; Kohl, S.; Thalmann, S.; Mateus, N.; Meyerhof, W.; de Freitas, V. Different phenolic compounds activate distinct human bitter taste receptors. J. Agric. Food Chem. 2013, 61, 1525–1533. [Google Scholar] [CrossRef] [PubMed]
- Clapham, D.E. TRP channels as cellular sensors. Nature 2003, 426, 517–524. [Google Scholar] [CrossRef] [PubMed]
- Viana, F. Chemosensory properties of the trigeminal system. ACS Chem. Neurosci. 2011, 2, 38–50. [Google Scholar] [CrossRef] [PubMed]
- Green, B.G. Chemesthesis and the chemical senses as components of a “chemofensor complex”. Chem. Senses 2012, 37, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Benigni, R.; Capra, C.; Cattorini, P.E. Piante Medicinali, Chimica, Farmacologia E Terapia; Inverni & Della: Milano, Italy, 1964; pp. 1372–1385. [Google Scholar]
- Hale, A.L.; Meepagala, K.M.; Oliva, A.; Aliotta, G.; Duke, S.O. Phytotoxins from the leaves of Ruta graveolens. J. Agric. Food Chem. 2004, 52, 3345–3349. [Google Scholar] [CrossRef] [PubMed]
- Haddouchi, F.; Chaouche, T. M.; Zaouali, Y.; Ksouri, R.; Attou, A.; Benmansour, A. Chemical composition and antimicrobial activity of the essential oils from four Ruta species growing in Algeria. Food Chem. 2013, 141, 253–258. [Google Scholar] [CrossRef] [PubMed]
- Ghirardini, M.P.; Carli, M.; del Vecchio, N.; Rovati, A.; Cova, O.; Valigi, F.; Agnetti, G.; Macconi, M.; Adamo, D.; Traina, M.; et al. The importance of a taste. A comparative study on wild food plant consumption in twenty-one local communities in Italy. J. Ethnobiol. Ethnomed. 2007, 3. [Google Scholar] [CrossRef] [PubMed]
- Sallal, A.J.; Alkofahi, A. Inibithion of the hemolytic activities of snake and scorpion venoms in vitro with plant extracts. Biomed. Lett. 1996, 53, 211–215. [Google Scholar]
- Kataki, M.S.; Kakoti, B.B.; Bhuyan, B.; Rajkumari, A.; Rajak, P. Garden rue inhibits the arachidonic acid pathway, scavenges free radicals, and elevates FRAP: Role in inflammation. Chin. J. Nat. Med. 2014, 12, 172–179. [Google Scholar] [CrossRef]
- Fadlalla, K.; Watson, A.; Yehualaeshet, T.; Turner, T.; Samuel, T. Ruta graveolens extract induces dna damage pathways and blocks akt activation to inhibit cancer cell proliferation and survival. Anticancer Res. 2011, 31, 233–241. [Google Scholar] [PubMed]
- Gasparro, F.P. Extracorporeal Photochemotherapy: Clinical Aspects and the Molecular Basis for Efficacy; RG Landes Company, CRC Press: Georgetown, TX, USA, 1994. [Google Scholar]
- Browne, F.; Turner, D.; Goulden, V. Psoralen and ultraviolet A in the treatment of granuloma annulare. Photodermatol. Photo 2011, 27, 81–84. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, H.; Yao, X.-G.; Shen, H.; Chen, J.; Li, C.; Zheng, M.; Ye, J.; Hu, L.; Shen, X.; Jiang, H. (+)-Rutamarin as a dual inducer of both GLUT4 traslocation and expression efficiently ameliorates glucose homeostasis in insulin resistant mice. PLoS ONE 2012, 7, e31811. [Google Scholar] [CrossRef] [PubMed]
- Ratheesh, M.; Sindhu, G.; Helen, A. Anti-inflammatory effect of quinoline alkaloid skimmianine isolated from Ruta graveolens L. Inflamm. Res. 2013, 62, 367–376. [Google Scholar] [CrossRef]
- Park, S.-H.; Sim, Y.-B.; Kim, S.-M.; Lee, J.-K.; Lim, S.-M.; Kim, J.-K.; Suh, H.-W. Antinociception effect and mechanism of Ruta graveolens L. In mice. J. Korean Soc. Appl. Biol. Chem. 2010, 53, 593–597. [Google Scholar] [CrossRef]
- Oliva, A.; Lahoz, E.; Contillo, R.; Aliotta, G. Fungistatic activity of Ruta graveolens extracts and its allelochemicals. J. Chem. Ecol. 1999, 25, 519–526. [Google Scholar] [CrossRef]
- Oliva, A.; Meepagala, K.M.; Wedge, D.E.; Harries, D.; Hale, A.L.; Aliotta, G.; Duke, S.O. Natural fungicides from Ruta graveolens L. Leaves, including a new quinolone alkaloid. J. Agric. Food Chem. 2003, 51, 890–896. [Google Scholar] [CrossRef] [PubMed]
- Astelbauer, F.; Gruber, M.; Brem, B.; Greger, H.; Obwaller, A.; Wernsdorfer, G.; Congpuong, K.; Wersdorfer, W.H.; Walochnik, J. Activity of selected phytochemicals against Plasmodium falciparum. Acta Trop. 2012, 123, 96–100. [Google Scholar] [CrossRef] [PubMed]
- Mann, R.S.; Tiwari, S.; Smoot, J.M.; Rouseff, R.L.; Stelinski, L.L. Repellency and toxicity of plant-based essential oils and their constituents against Diaphorina citri Kuwayama (Hemiptera: Psyllidae). J. Appl. Entomol. 2012, 136, 87–96. [Google Scholar] [CrossRef]
- Innocent, E.; Gikonyo, N.K.; Nkunya, M.H. Repellency property of long chain aliphatic methyl ketones against Anopheles gambiae s.s. Tanzan J. Health Res. 2008, 10, 50–54. [Google Scholar] [CrossRef] [PubMed]
- Bassoli, A.; Borgonovo, G.; Busnelli, G. Alkaloids and the bitter taste. In Modern Alkaloids, Structure, Isolation, Synthesis and Biology; Fattorusso, E., Taglialatela Scafati, O., Fattorusso, E., Taglialatela-Scafati, O., Eds.; Wiley VCH: Weinheim, Germany, 2007; pp. 53–70. [Google Scholar]
- Born, S.; Levit, A.; Niv, M.Y.; Meyerhof, W.; Behrens, M. The human bitter taste receptor TAS2R10 is tailored to accommodate numerous diverse ligands. J. Neurosci. 2013, 33, 201–213. [Google Scholar] [CrossRef] [PubMed]
- Bassoli, A.; Borgonovo, G.; Caimi, S.; Scaglioni, L.; Morini, G.; Schiano Moriello, A.; di Marzo, V.; de Petrocellis, L. Taste-guided identification of high potency TRPA1 agonists from Perilla frutescens. J. Biorg. Med. Chem. 2009, 17, 1636–1639. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Barrit, G.J. Evidence that TRPM8 is an androgen dependent Ca2+ channel required for the survival of the prostate cancer cells. Cancer Res. 2004, 64, 8365–8373. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Barrit, G.J. TRPM8 is prostate cancer cells: A potential diagnostic and prognostic marker with a secretory function? Endocr. Relat. 2006, 13, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Chianese, G.; Fattorusso, E.; Putra, M.Y.; Calcinai, B.; Bavestrello, G.; Schiano Moriello, A.; de Petrocellis, L.; di Marzo, V.; Taglialatela Scafati, O. Leuccettamols, bifunctionalised marine sphingoids, act as modulators of TRPA1 and TRPM8 channels. Mar. Drugs 2012, 10, 2435–2447. [Google Scholar] [CrossRef] [PubMed]
- Terada, Y.; Horie, S.; Takayama, H.; Uchida, K.; Tominaga, M.; Watanabe, T. Activation and Inhibition of Thermosensitive TRP Channels by Voacangine, an Alkaloid Present in Voacanga africana, an African Tree. J. Nat. Prod. 2014, 77, 285–297. [Google Scholar] [CrossRef] [PubMed]
- Baraldi, P.G.; Preti, D.; Materazzi, S.; Geppetti, P. Transient receptor potential ankyrin 1 (TRPA1) channel as emerging target for novel analgesics and anti-inflammatory agents. J. Med. Chem. 2010, 53, 5085–5107. [Google Scholar] [CrossRef] [PubMed]
- Knowlton, W.M.; Bifolk-Fisher, A.; Bautista, D.M.; McKemy, D.D. TRPM8 but not TRPA1 is required for neural and behavioural responses to acute noxious cold temperature and cold-mimetic in vivo. Pain 2010, 150, 340–350. [Google Scholar] [CrossRef] [PubMed]
- Brignell, J.L.; Chapman, V.; Kendall, D.A. Comparison of icilin- and cold- evoked responses of spinal neurones, and their modulation of mechanical activity, in a model of neurophatic pain. Brain Res. 2008, 1215, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.N.; Zhang, S.L.; Ma, L.; Zhang, Y.; Shen, X.; Wang, W.; Hu, L.H. Total synthesis of (+)-rutamarin. Adv. Synt. Catal. 2008, 350, 2373–2379. [Google Scholar]
- Muller, M.; Byres, M.; Jaspars, M.; Kumarasamy, Y.; Middleton, M.; Nahar, L.; Shoeb, M.; Sarker, S.D. 2D NMR Spectroscopic Analyses of Archangelicin from the Seed of Angelica archangelica. Acta Pharm. 2004, 54, 277–285. [Google Scholar] [PubMed]
- Liu, R.; Feng, L.; Sun, A.; Kong, L. Preparative isolation and purification of coumarins from Cnidium monnieri (L.) Cusson by high-speed counter-current chromatography. J. Chromatog. A 2004, 1055, 71–76. [Google Scholar] [CrossRef]
- Wu, T.S.; Shi, L.S.; Wang, J.J.; Iou, S.C.; Chang, H.C.; Chen, Y.P.; Kuo, Y.H.; Chang, Y.L.; Teng, C.M. Cytotoxic and Antiplatelet Aggregation Principles of Ruta graveolens. J. Chin. Soc. 2003, 50, 171–178. [Google Scholar] [CrossRef]
- Phadke, P.C.; Kelkar, S.L.; Wadia, M.S. A general synthesis of bis[coumarinyl]ethers: Synthesis of daphnoretin methyl ether. Synthesis 1986, 5, 413–415. [Google Scholar] [CrossRef]
- Bassoli, A.; Borgonovo, G.; Caremoli, F.; Mancuso, G. The taste of d- and l-amino acids: In vitro binding assays with cloned human bitter (TAS2Rs) and sweet (TAS1R2/TAS1R3) receptors. Food Chem. 2014, 150, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the compounds 1–10 are not available from the authors.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mancuso, G.; Borgonovo, G.; Scaglioni, L.; Bassoli, A. Phytochemicals from Ruta graveolens Activate TAS2R Bitter Taste Receptors and TRP Channels Involved in Gustation and Nociception. Molecules 2015, 20, 18907-18922. https://doi.org/10.3390/molecules201018907
Mancuso G, Borgonovo G, Scaglioni L, Bassoli A. Phytochemicals from Ruta graveolens Activate TAS2R Bitter Taste Receptors and TRP Channels Involved in Gustation and Nociception. Molecules. 2015; 20(10):18907-18922. https://doi.org/10.3390/molecules201018907
Chicago/Turabian StyleMancuso, Giuseppe, Gigliola Borgonovo, Leonardo Scaglioni, and Angela Bassoli. 2015. "Phytochemicals from Ruta graveolens Activate TAS2R Bitter Taste Receptors and TRP Channels Involved in Gustation and Nociception" Molecules 20, no. 10: 18907-18922. https://doi.org/10.3390/molecules201018907
APA StyleMancuso, G., Borgonovo, G., Scaglioni, L., & Bassoli, A. (2015). Phytochemicals from Ruta graveolens Activate TAS2R Bitter Taste Receptors and TRP Channels Involved in Gustation and Nociception. Molecules, 20(10), 18907-18922. https://doi.org/10.3390/molecules201018907