
Anti-Inflammatory Activity Comparison among Scropoliosides—Catalpol Derivatives with 6-O-Substituted Cinnamyl Moieties

Abstract
:1. Introduction

2. Results and Discussion
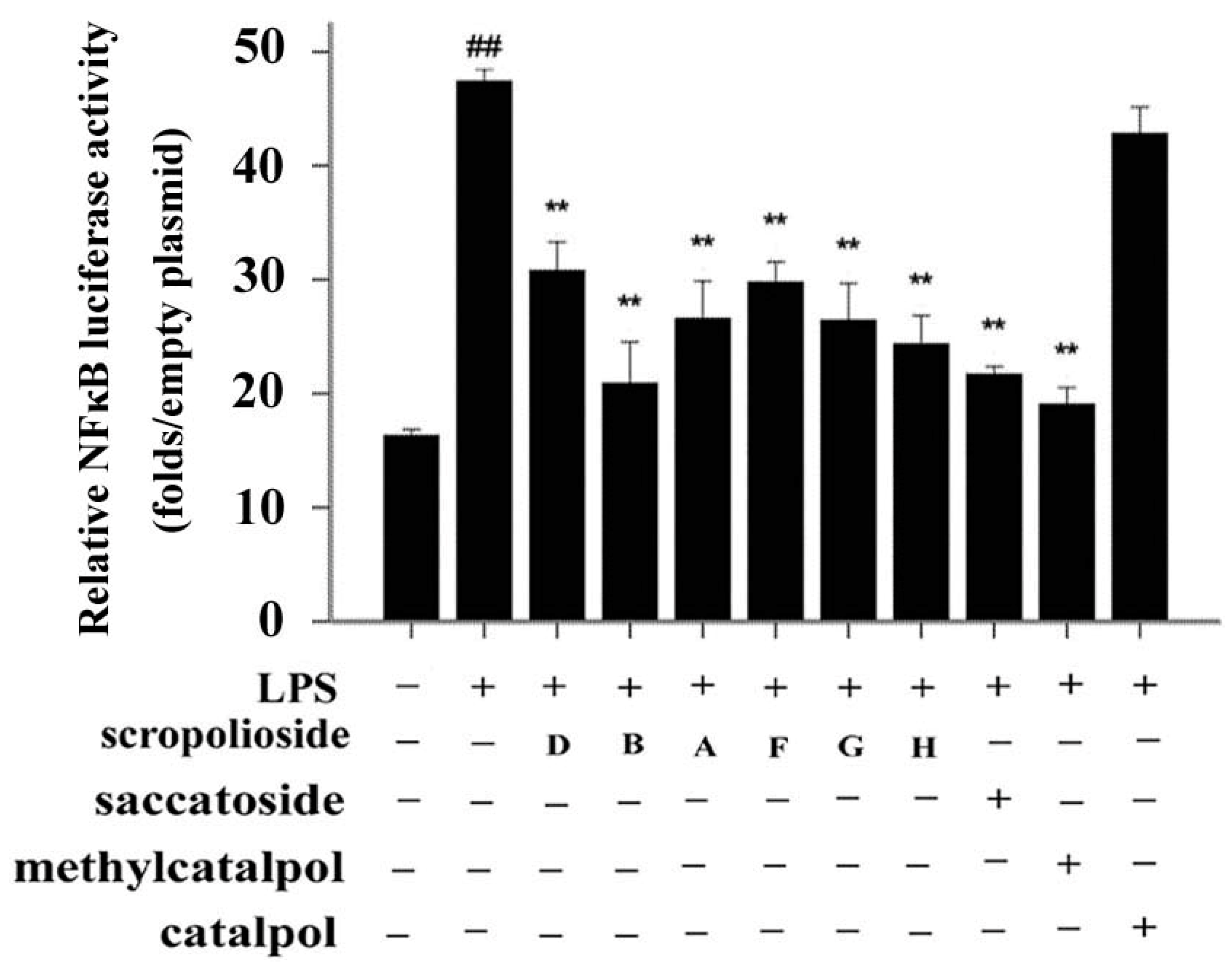
2.1. Effect of Iridoid Glycosides on NF-κB Activation

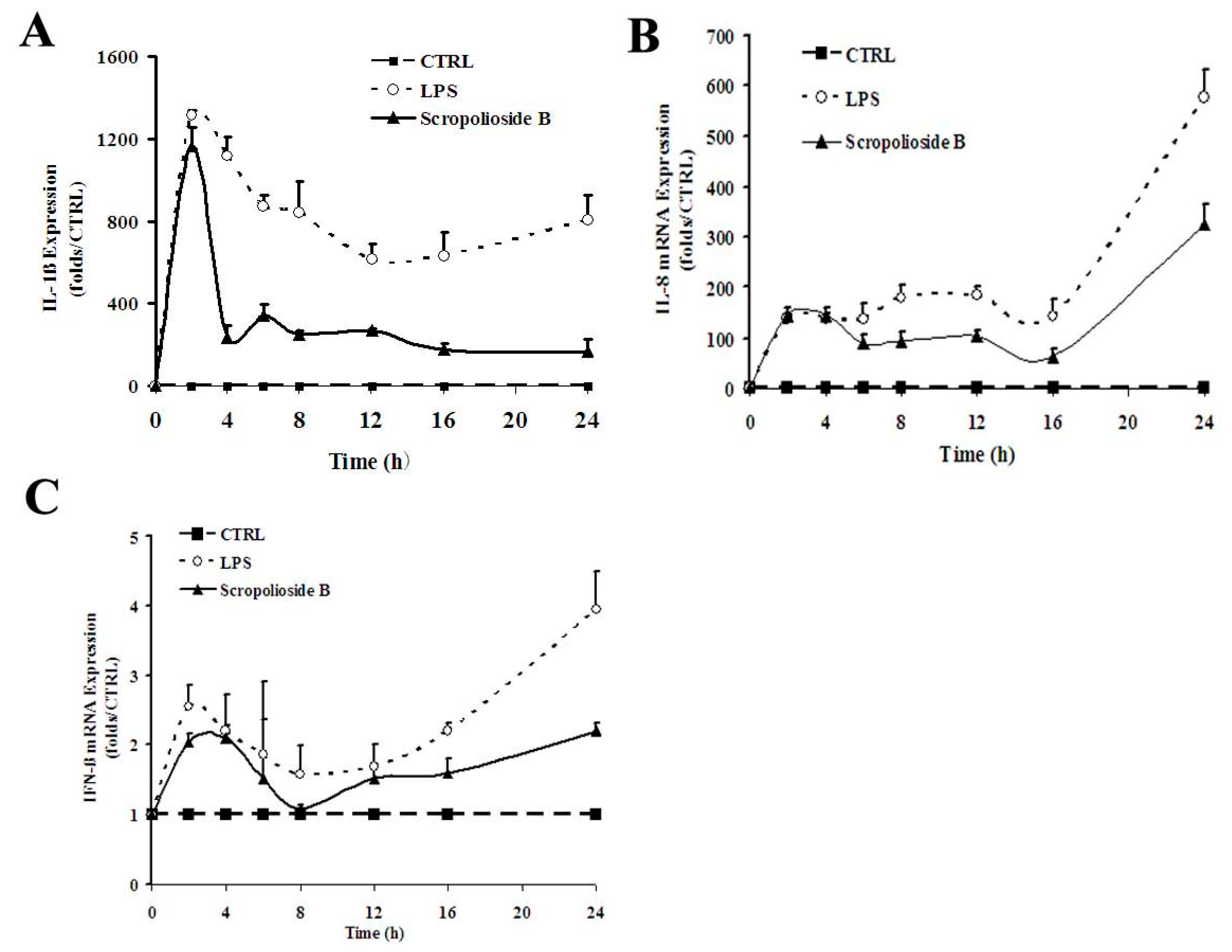
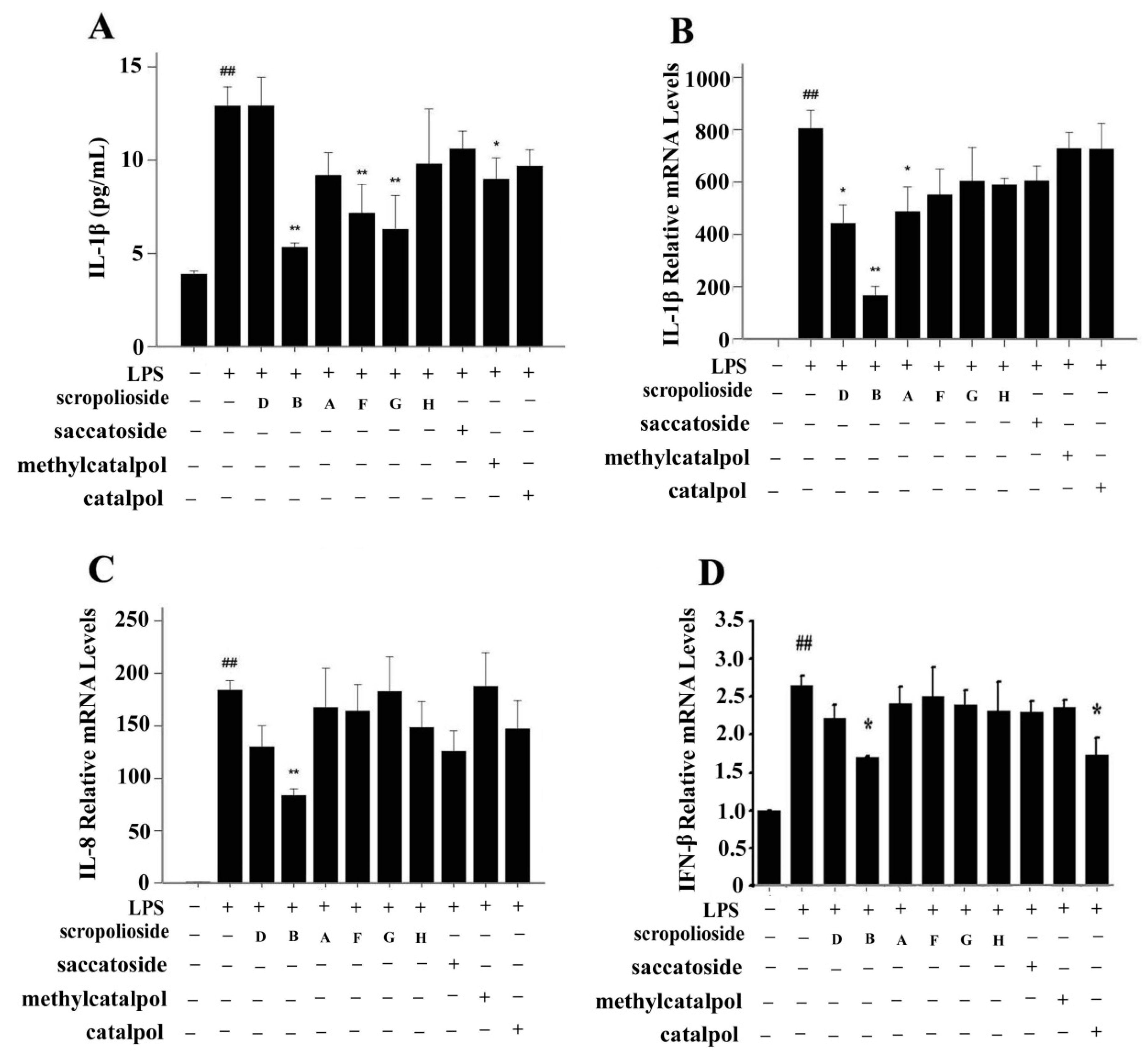
2.2. Cytokine Expression


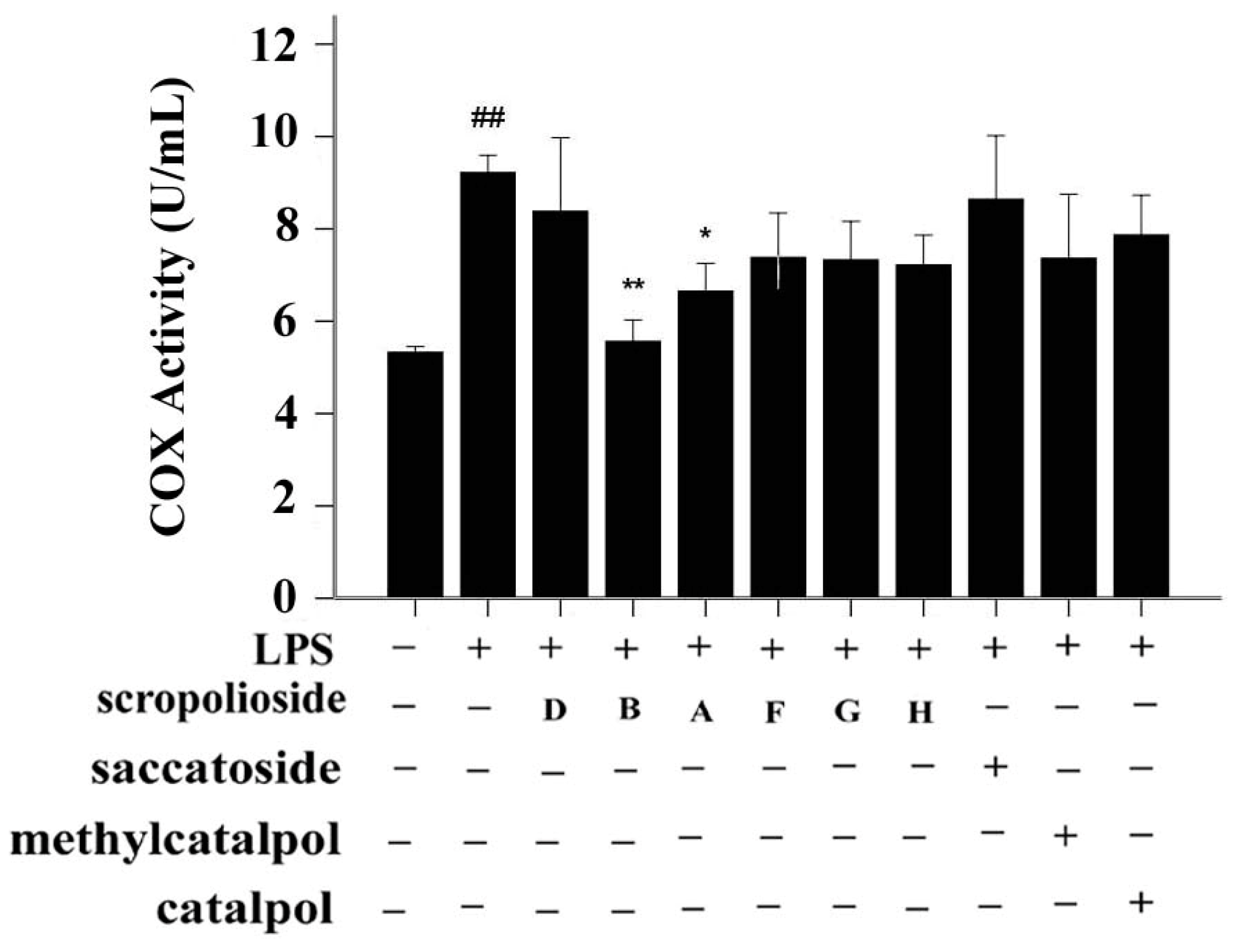
2.3. Activity of Arachidonic-Acid-Metabolizing Enzymes

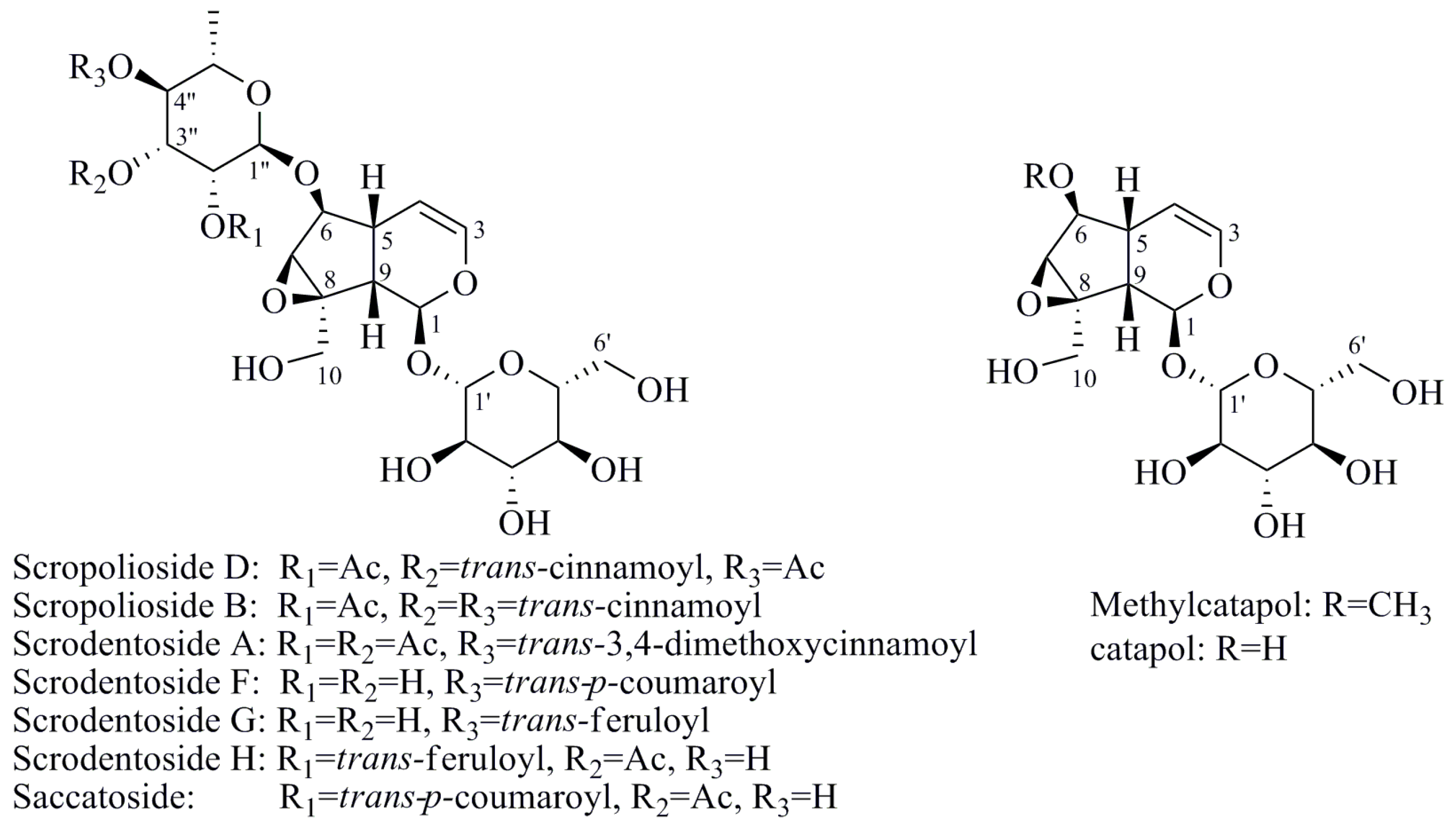
2.4. Structure-Activity Relationship of the Eight Catalpol Derivatives and Catalpol

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Structure | Inhibitiory Ratio of NF-κB Activity | Inhibitiory Ratio of IL-1β mRNA Expression | Inhibitiory Ratio of IL-1β Protein Expression | Inhibitiory Ratio of IL-8 mRNA Expression | Inhibitiory Ratio of IFN-β mRNA Expression | Inhibitiory Ratio of COX-2 Activity |
|---|---|---|---|---|---|---|---|
| Scropolioside B |  | 49.1 | 79.5 | 58.8 | 54.6 | 35.9 | 32.3 |
| Scropolioside D |  | 40.9 | 45.1 | −0.1 | 29.4 | 16.5 | −1.9 |
| Scrodentoside A |  | 40.2 | 39.5 | 28.9 | 8.8 | 9.25 | 19.1 |
| Scrodentoside F |  | 41.9 | 31.6 | 44.5 | 10.8 | 5.6 | 10.2 |
| Scrodentoside G |  | 50.4 | 25 | 51.3 | 0.8 | 10.1 | 10.8 |
| Scrodentoside H |  | 52.8 | 26.8 | 24.1 | 19.3 | 13.5 | 12.1 |
| Saccatoside |  | 58.1 | 24.9 | 17.9 | 31.7 | 11.2 | −5.1 |
| Methylcatapol |  | 59.2 | 9.6 | 51.3 | -2 | 8.9 | 10.4 |
| Catalpol |  | −2.3 | 9.8 | 25 | 19.9 | 34.7 | 4.2 |
2.5. Effect of Cinnamyl Moieties in Scropoliosides on Anti-inflammatory Activity
2.6. NF-κB Activity, Cytokine Expression and Release, and the Inhibitory Effect of Scropoliosides
3. Experimental Section
3.1. General Information
3.2. Cell Culture and Reagents
3.3. Extraction and Isolation of Iridoid Glycosides from S. dentata Royle ex Benth
3.3.1. 6-O-α-l-(4′′-O-trans-p-Coumaroyl)rhamnopyranosylcatalpol (Scropolioside F)
3.3.2. 6-O-α-l-(4′′-O-Feruloyl)rhamnopyranosylcatalpol (Scropolioside G)
3.3.3. 6-O-α-l-(2′′-O-Feruloyl)rhamnopyranosylcatalpol (Scropolioside H)
3.3.4. Saccatoside
3.3.5. 6-O-Methylcatapol
3.4. Luciferase Assay
3.5. Quantitative Real-Time PCR
| Gene | Direction | Primer Sequences |
|---|---|---|
| IL-1β | Forward Reverse | 5-AAACAGATGAAGTGCTCCTTCCAGG-3 5-TGGAGAACACCACTTGTTGCTCCA-3 |
| IL-8 | Forward Reverse | 5-ATGGCTGCTGAACCAGTAGA-3 5-CTAGTCTTCGTTTTGAACAG-3 |
| IFN-β | Forward Reverse | 5-GCCTCAAGGACAGGATGAAC-3 5-AGCCAGGAGGTTCTCAACAA-3 |
| GAPDH | Forward Reverse | 5-AGAAGGCTGGGGCTCATTTG-3 5-AGGGGCCATCCACAGTCTTC-3 |
3.6. ELISA
3.7. Screening Assay for 15-Lipoxygenase Inhibitor
3.8. COX-2 Activity Assay
3.9. Data Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
References
- Viljoen, A.; Mncwangi, N.; Vermaak, I. Anti-inflammatory iridoids of botanical origin. Curr. Med. Chem. 2012, 19, 2104–2127. [Google Scholar] [CrossRef] [PubMed]
- Tundis, R.; Loizzo, M.R.; Menichini, F.; Statti, G.A.; Menichini, F. Biological and pharmacological activities of iridoids: Recent developments. Mini Rev. Med. Chem. 2008, 8, 399–420. [Google Scholar] [CrossRef] [PubMed]
- Giner, R.M.; Villalba, M.L.; Recio, M.C.; Máñez, S.; Cerdá-Nicolás, M.; Ríos, J. Anti-inflammatory glycoterpenoids from Scrophularia auriculata. Eur. J. Pharmacol. 2000, 389, 243–252. [Google Scholar] [CrossRef]
- Dinda, B.; Debnath, S.; Banik, R. Naturally occurring iridoids and secoiridoids. An updated review, part 4. Chem. Pharm. Bull. 2011, 59, 803–833. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Feng, L.; Jia, Q.; Xu, J.; Wang, R.; Wang, Z.; Wu, Y.; Li, Y. Effects of β-glucosidase hydrolyzed products of harpagide and harpagoside on cyclooxygenase-2 (COX-2) in vitro. Bioorg. Med. Chem. 2011, 19, 4882–4886. [Google Scholar] [CrossRef] [PubMed]
- Park, K.S.; Kim, B.H.; Chang, I.M. Inhibitory Potencies of Several Iridoids on Cyclooxygenase-1, Cyclooxygnase-2 Enzymes Activities, Tumor Necrosis factor-α and Nitric Oxide Production in Vitro. Evid. Based. Complement. Altern. Med. 2010, 7, 41–45. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.J.; Jang, H.J.; Chung, T.W.; Jeong, S.I.; Cha, J.; Choi, J.Y.; Han, C.W.; Jang, Y.S.; Joo, M.; Jeong, H.S.; et al. Catalpol suppresses advanced glycation end-products-induced inflammatory responses through inhibition of reactive oxygen species in human monocytic THP-1 cells. Fitoterapia 2013, 86, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Fu, K.; Piao, T.; Wang, M.; Zhang, J.; Jiang, J.; Wang, X.; Liu, H. Protective effect of catalpol on lipopolysaccharide-induced acute lung injury in mice. Int. Immunopharmacol. 2014, 23, 400–406. [Google Scholar] [CrossRef] [PubMed]
- Zhu, T.; Zhang, L.; Ling, S.; Duan, J.; Qian, F.; Li, Y.; Xu, J.W. Scropolioside B inhibits IL-1β and cytokines expression through NF-κB and inflammasome NLRP3 pathways. Mediat. Inflamm. 2014, 2014, 819053. [Google Scholar] [CrossRef] [PubMed]
- Bas, E.; Recio, M.C.; Máñez, S.; Giner, R.M.; Escandell, J.M.; López-Ginés, C.; Ríos, J.L. New insight into the inhibition of the inflammatory response to experimental delayed-type hypersensitivity reactions in mice by scropolioside A. Eur. J. Pharmacol. 2007, 555, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Helfrich, E.; Rimpler, H. Iridoid glycosides and phenolic glycosides from Holmskioldia sanguinea. Phytochemistry 1999, 50, 619–627. [Google Scholar] [CrossRef]
- Warashina, T.; Miyase, T.; Ueno, A. Iridoid glycosides from Verbascum thapsus L. Chem. Pharm. Bull. 1991, 39, 3261–3264. [Google Scholar] [CrossRef]
- Otsuka, H.; Kubo, N.; Sasaki, Y.; Yamasaki, K.; Takeda, Y.; Seki, T. Iridoid diglycoside monoacyl esters from stems of Premna japonica. Phytochemistry 1991, 30, 1917–1920. [Google Scholar] [CrossRef]
- Otsuka, H.; Sasaki, Y.; Yamasaki, K.; Takeda, Y.; Seki, T. Iridoid Diglycoside Monoacyl Esters from the Leaves of Premna japonica. J. Nat. Prod. 1990, 53, 107–111. [Google Scholar] [CrossRef]
- Zhang, W.J.; Liu, Y.Q.; Li, X.C.; Pu, X.Y.; Jin, Y.Q.; Yang, C.R. Chemical constituents from Scrophularia ningpoensis. Acta Bot. Yunnanica 1994, 16, 407–412. [Google Scholar]
- Recio, M.C.; Giner, R.M.; Máñez, S.; Ríos, J.L. Structural considerations on the iridoids as anti-inflammatory agents. Planta Med. 1994, 60, 232–234. [Google Scholar] [CrossRef] [PubMed]
- Park, K.S.; Chang, I.M. Anti-inflammatory activity of aucubin by inhibition of tumor necrosis factor-alpha production in RAW 264.7 cells. Planta Med. 2004, 70, 778–779. [Google Scholar] [CrossRef] [PubMed]
- Koo, H.J.; Lim, K.H.; Jung, H.J.; Park, E.H. Anti-inflammatory evaluation of gardenia extract, geniposide and genipin. J. Ethnopharmacol. 2006, 103, 496–500. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, B.; Al-Rehaily, A.J.; Al-Howiriny, T.A.; El-Sayed, K.A.; Ahmad, M.S. Scropolioside-D2 and harpagoside-B: Two new iridoid glycosides from Scrophularia deserti and their antidiabetic and antiinflammatory activity. Biol. Pharm. Bull. 2003, 26, 462–467. [Google Scholar] [CrossRef] [PubMed]
- Cartron, E.; Carbonneau, M.A.; Fouret, G.; Descomps, B.; Léger, C.L. Specific antioxidant activity of caffeoyl derivatives and other natural phenolic compounds: LDL protection against oxidation and decrease in the proinflammatory lysophosphatidylcholine production. J. Nat. Prod. 2001, 64, 480–486. [Google Scholar] [CrossRef] [PubMed]
- De Cássia da Silveira E Sá, R.; Andrade, L.N.; Dos Reis Barreto de Oliveira, R.; de Sousa, D.P. A review on anti-inflammatory activity of phenylpropanoids found in essential oils. Molecules 2014, 19, 1459–1480. [Google Scholar] [CrossRef] [PubMed]
- Kurokawa, M.; Brown, J.; Kagawa, Y.; Shiraki, K. Cytokine-regulatory activity and therapeutic efficacy of cinnamyl derivatives in endotoxin shock. Eur. J. Pharmacol. 2003, 474, 283–293. [Google Scholar] [CrossRef]
- Ropert, C.; Closel, M.; Chaves, A.C.; Gazzinelli, R.T. Inhibition of a p38/stress-activated protein kinase-2-dependent phosphatase restores function of IL-1 receptor-associate kinase-1 and reverses Toll-like receptor 2- and 4-dependent tolerance of macrophages. J. Immunol. 2003, 171, 1456–1465. [Google Scholar] [CrossRef] [PubMed]
- Basak, C.; Pathak, S.K.; Bhattacharyya, A.; Mandal, D.; Pathak, S.; Kundu, M. NF-κB- and C/EBPβ-driven interleukin-1beta gene expression and PAK1-mediated caspase-1 activation play essential roles in interleukin-1beta release from Helicobacter pylori lipopolysaccharide-stimulated macrophages. J. Biol. Chem. 2005, 280, 4279–4288. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.T.; Cohen, P.; Rousseau, S. IL-1β-stimulated activation of ERK1/2 and p38α MAPK mediates the transcriptional up-regulation of IL-6, IL-8 and GRO-α in HeLa cells. Cell Signal. 2008, 20, 375–380. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Ye, F.; Xiong, H.; Hu, D.; Limb, G.A.; Xie, T.; Peng, L.; Yang, W.; Sun, Y.; Zhou, M.; et al. IL-1β Upregulates IL-8 Production in Human Müller Cells through Activation of the p38 MAPK and ERK1/2 Signaling Pathways. Inflammation 2014, 37, 1486–1495. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, C.; Kamata, H.; Takeshita, H.; Yagisawa, H.; Hirata, H. Redox regulation of lipopolysaccharide (LPS)-induced interleukin-8 (IL-8) gene expression mediated by NF kappa B and AP-1 in human astrocytoma U373 cells. Biochem. Biophys. Res. Commun. 1997, 232, 568–573. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Bai, N.; Chang, A.; Zhang, Z.; Yin, J.; Shen, W.; Tian, Y.; Xiang, R.; Liu, C. ATF4 is directly recruited by TLR4 signaling and positively regulates TLR4-trigged cytokine production in human monocytes. Cell. Mol. Immunol. 2013, 10, 84–94. [Google Scholar] [CrossRef] [PubMed]
- Toshchakov, V.; Jones, B.W.; Perera, P.Y.; Thomas, K.; Cody, M.J.; Zhang, S.; Williams, B.R.; Major, J.; Hamilton, T.A.; Fenton, M.J.; et al. TLR4, but not TLR2, mediates IFN-β-induced STAT1α/β-dependent gene expression in macrophages. Nat. Immunol. 2002, 3, 392–398. [Google Scholar] [CrossRef] [PubMed]
- Eskan, M.A.; Rose, B.G.; Benakanakere, M.R.; Lee, M.J.; Kinane, D.F. Sphingosine 1-phosphate 1 and TLR4 mediate IFN-β expression in human gingival epithelial cells. J. Immunol. 2008, 180, 1818–1825. [Google Scholar] [CrossRef] [PubMed]
- Kato, A.; Ogasawara, T.; Homma, T.; Saito, H.; Matsumoto, K. Lipopolysaccharide-binding protein critically regulates lipopolysaccharide-induced IFN-beta signaling pathway in human monocytes. J. Immunol. 2004, 172, 6185–6194. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Liu, Y.P.; Sha, H.; Chen, H.; Qi, L.; Smith, J.A. XBP-1 couples endoplasmic reticulum stress to augmented IFN-beta induction via a cis-acting enhancer in macrophages. J. Immunol. 2010, 185, 2324–2330. [Google Scholar] [CrossRef] [PubMed]
- Lamkanfi, M.; Dixit, V.M. Mechanisms and functions of inflammasomes. Cell 2014, 157, 1013–1022. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Callaway, J.B.; Ting, J.P. Inflammasomes: Mechanism of action, role in disease, and therapeutics. Nat. Med. 2015. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhu, T.; Qian, F.; Xu, J.; Dorje, G.; Zhao, Z.; Guo, F.; Li, Y. Iridoid glycosides isolated from Scrophularia dentata Royle ex Benth. and their anti-inflammatory activity. Fitoterapia 2014, 98, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Not avaliable.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, T.; Zhang, L.; Ling, S.; Qian, F.; Li, Y.; Xu, J.-W. Anti-Inflammatory Activity Comparison among Scropoliosides—Catalpol Derivatives with 6-O-Substituted Cinnamyl Moieties. Molecules 2015, 20, 19823-19836. https://doi.org/10.3390/molecules201119659
Zhu T, Zhang L, Ling S, Qian F, Li Y, Xu J-W. Anti-Inflammatory Activity Comparison among Scropoliosides—Catalpol Derivatives with 6-O-Substituted Cinnamyl Moieties. Molecules. 2015; 20(11):19823-19836. https://doi.org/10.3390/molecules201119659
Chicago/Turabian StyleZhu, Tiantian, Liuqiang Zhang, Shuang Ling, Fei Qian, Yiming Li, and Jin-Wen Xu. 2015. "Anti-Inflammatory Activity Comparison among Scropoliosides—Catalpol Derivatives with 6-O-Substituted Cinnamyl Moieties" Molecules 20, no. 11: 19823-19836. https://doi.org/10.3390/molecules201119659
APA StyleZhu, T., Zhang, L., Ling, S., Qian, F., Li, Y., & Xu, J. -W. (2015). Anti-Inflammatory Activity Comparison among Scropoliosides—Catalpol Derivatives with 6-O-Substituted Cinnamyl Moieties. Molecules, 20(11), 19823-19836. https://doi.org/10.3390/molecules201119659