
Targeted and Untargeted Metabolomics to Explore the Bioavailability of the Secoiridoids from a Seed/Fruit Extract (Fraxinus angustifolia Vahl) in Human Healthy Volunteers: A Preliminary Study

 , ,
, , 

Abstract
:
1. Introduction

2. Results and Discussion
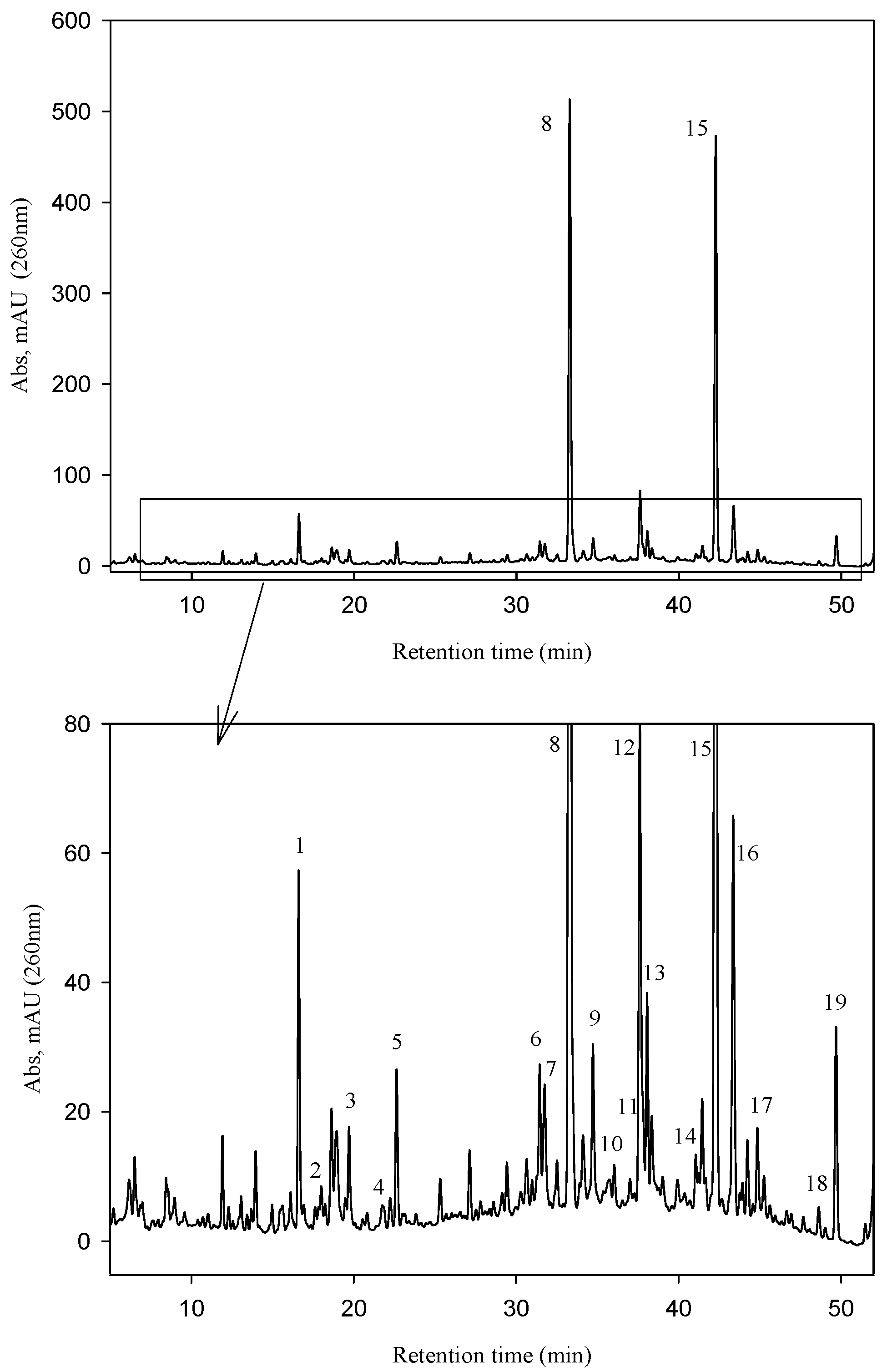
2.1. Fraxinus Extract Content in Secoiridoids


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak | Compounds | Rt (UV) | [M − H]− | MS/MS |
|---|---|---|---|---|
| 1 | Salidroside | 16.60 | 299 | 179,119 |
| 2 | Hydroxycinnamic acid derivative | 16.92 | 487 | 295, 179, 135 |
| 3 | Oleoside | 19.70 | 389 | 371, 345, 209, 179 |
| 4 | Oleoside 11-methyl ester | 22.63 | 403 | 223, 179 |
| 5 | Hydroxycinnamic acid derivative | 21.75 | 369 | 207, 192, 179 |
| 6 | Verbascoside | 31.45 | 623 | 461, 315, 251 |
| 7 | Nuzhenide isomer | 31.75 | 685 | 523, 453, 421, 299, 223 |
| 8 | Nuzhenide | 33.28 | 685 | 523, 453, 421, 299, 223 |
| 9 | Nuzhenide isomer | 34.72 | 685 | 523, 453, 421, 299, 223 |
| 10 | 1-O-β-d-Glucosylformoside | 36.03 | 685 | 523, 453, 385, 299, 223 |
| 11 | Nuzhenide derivative | 37.01 | 727 | 685, 565, 523, 453, 341, 299 |
| 12 | Excelside B | 37.61 | 685 | 565, 361, 291, 260 |
| 13 | GL3 isomer * | 41.06 | 1071 | 909, 685, 523, 385 |
| 14 | GL3 isomer * | 41.45 | 1071 | 909, 771, 685, 609, 421 |
| 15 | GL3 | 42.20 | 1071 | 909, 771, 685, 453, 385 |
| 16 | GL3 isomer * | 43.36 | 1071 | 909, 685, 478, 361 |
| 17 | Nuzhenide di (11-methyloleoside) | 44.96 | 1457 | 1157, 1071, 934, 771, 685 |
| 18 | GL5 isomer | 48.70 | 909 | 771, 747, 523, 361, 259 |
| 19 | GL5 isomer | 49.77 | 909 | 771, 747, 645, 523, 361 |
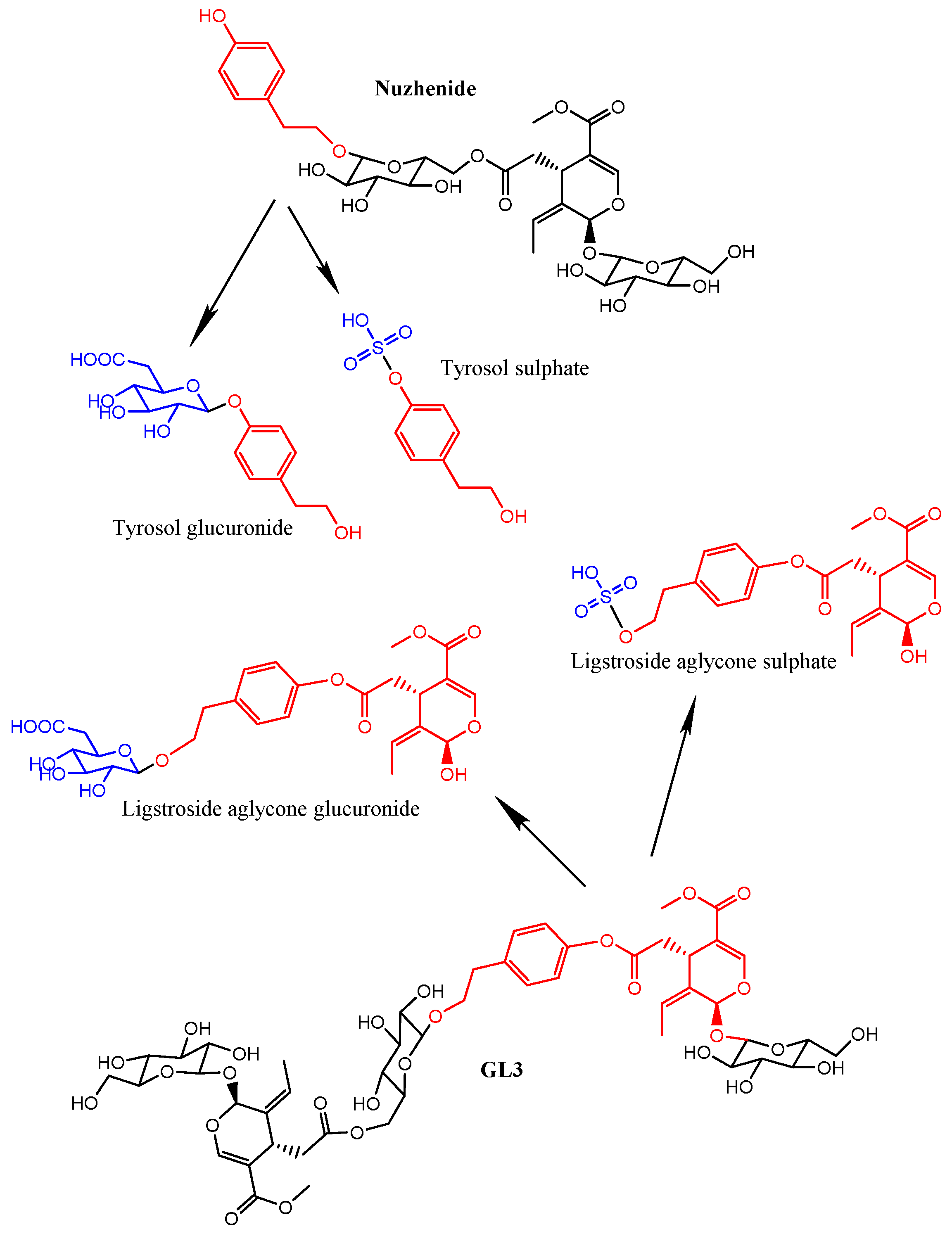
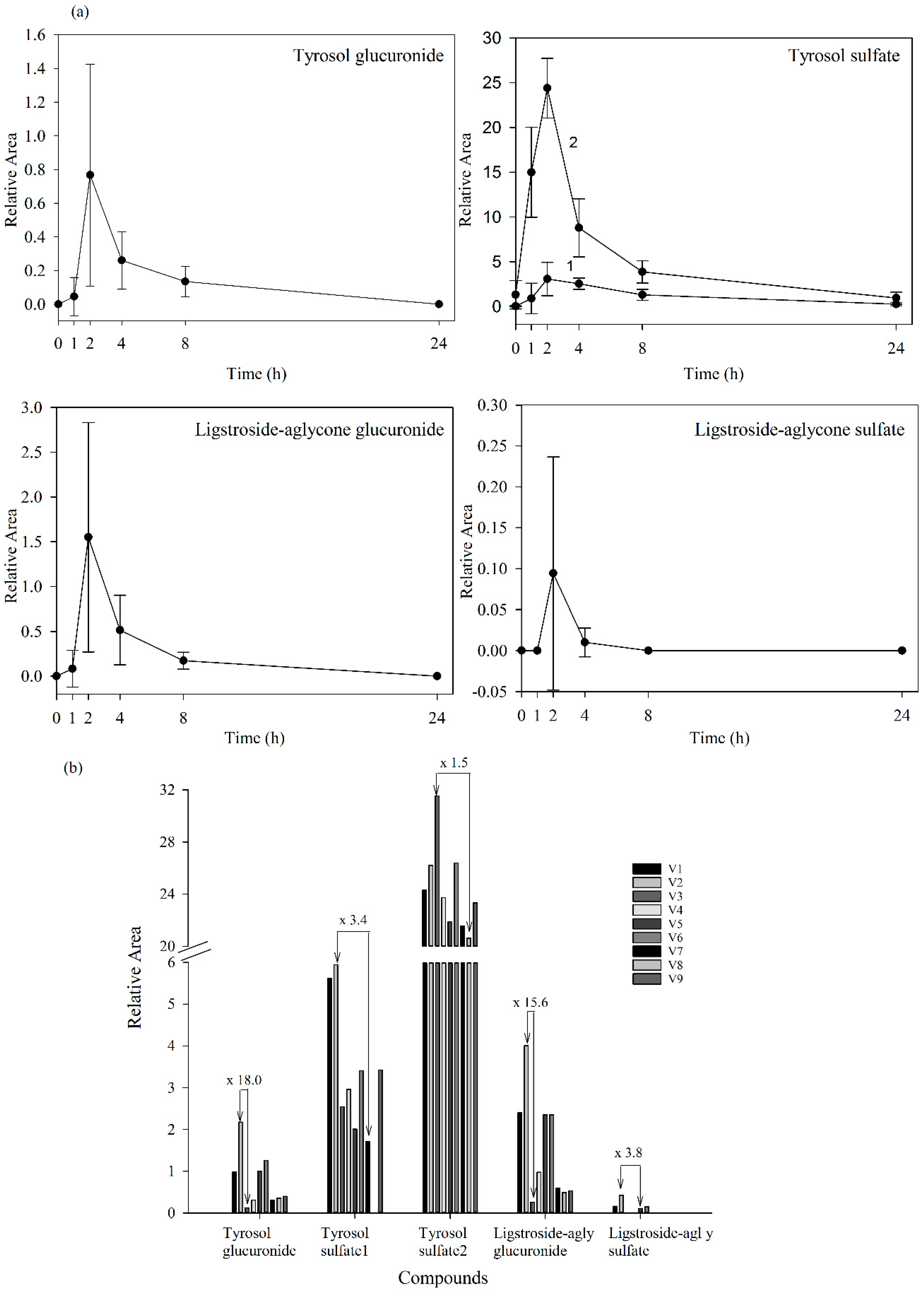
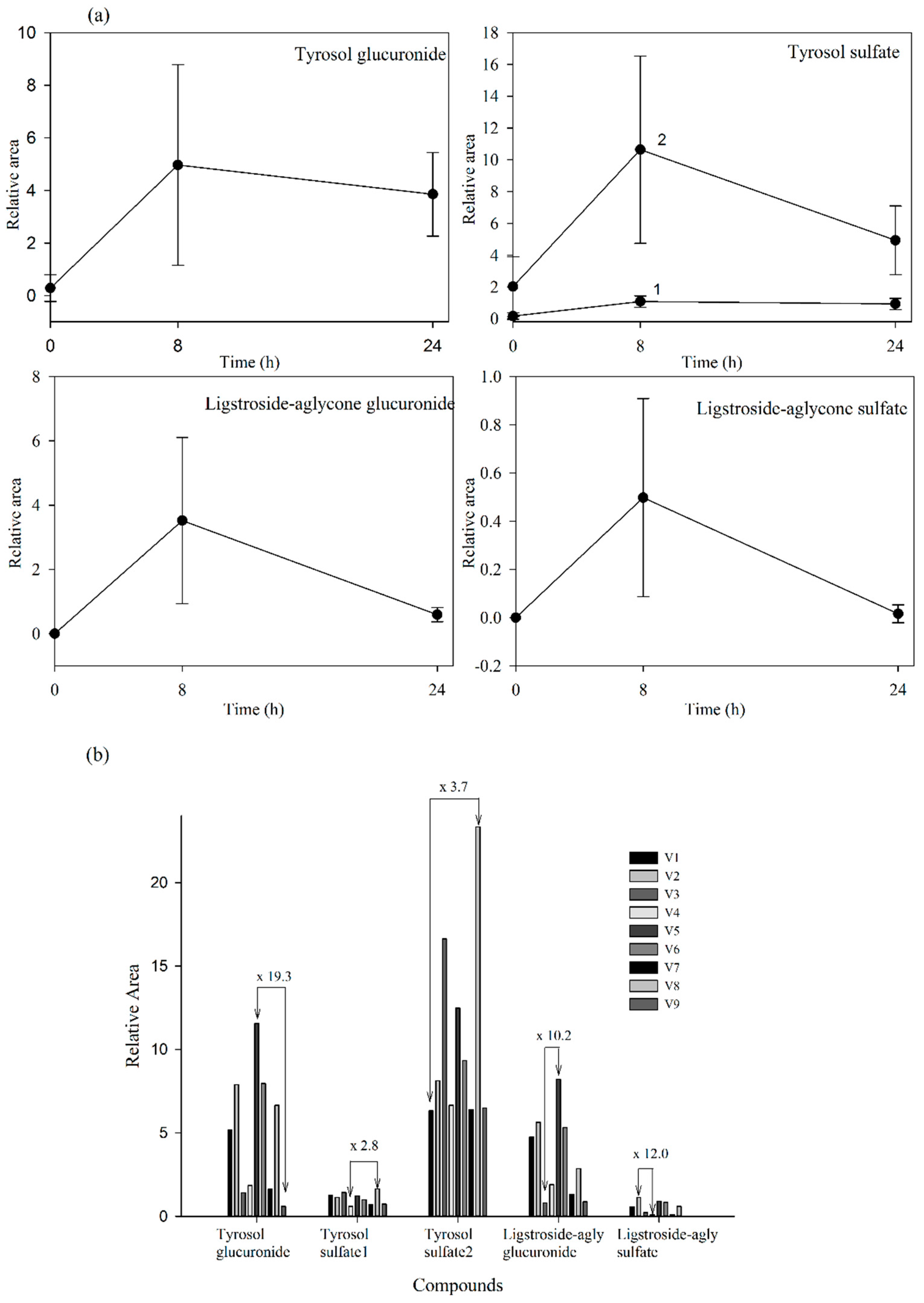
2.2. Fraxinus Extract Derived Specific Plasma and Urine Metabolites: Targeted Analysis
| Compounds | Retention Time (min) | m/z Experimental | Score | Error | Molecular Formulae | MS/MS Fragments |
|---|---|---|---|---|---|---|
| Tyrosol glucuronide | 5.30 | 313.0937 | 91.75 | −3.13 | C14H18O8 | 137.0608, 175.0247 |
| Tyrosol sulfate 1 | 5.79 | 217.0168 | 96.09 | 3.80 | C8H10O5S | 137.0604, 79.9572 |
| Tyrosol sulfate 2 | 13.00 | 217.0174 | 95.35 | 1.78 | C8H10O5S | 137.0606, 79.9574 |
| Ligstroside-aglycone * glucuronide | 14.66 | 537.1613 | 97.9 | −0.05 | C25H30O13 | 361.1288, 175.0246 |
| Ligstroside-aglycone * sulfate | 15.02 | 441.0868 | 95.6 | −1.74 | C19H22O10S | 361.1295, 79.9577 |



2.3. Untargeted Approach to Study Non-Predicted Secoiridoid Metabolites Appearing in Plasma and Urine Samples


| Exact Mass | Retention Time (min) | Error (ppm) | Score | MS/MS | Molecular Formula | Tentatively Identified Metabolite | Time-Points (h) * | p(corr) |
|---|---|---|---|---|---|---|---|---|
| Plasma | ||||||||
| 259.9985 | 8.28 | 0.02 | 98.12 | 179.0352, 135.0455, 96.9592 | C9H8O7S | Caffeic acid sulfate | 1, 2, 4, 8 | 3.85 × 10−8 |
| 274.014 | 8.58 | 1.92 | 98.53 | 193.0505 | C10H10O7S | Ferulic acid sulfate | 2, 4 | 1.70 × 10−3 |
| 204.0087 | 10.41 | 3.98 | 94.87 | 123.0449, 79.9575 | C7H8O5S | 4-hydroxybenzyl alcohol sulfate | 1 ,2, 4 | 3.50 × 10−6 |
| 274.0142 | 10.98 | 2.01 | 97.79 | 193.0505 | C10H10O7S | Ferulic acid sulfate | 1, 2 | 1.04 × 10−15 |
| 216.0087 | 12.20 | 1.55 | 98.84 | 135.0047 | C8H8O5S | 4-hydroxyphenylacetaldehyde sulfate | 2, 4 | 9.55 × 10−6 |
| 218.024 | 13.01 | 2.3 | 97.58 | 137.0606, 122.0374, 79.9574 | C8H10O5S | Tyrosol sulfate | 1, 2, 4 | 1.98 × 10−12 |
| 538.1688 | 14.64 | −0.02 | 99.04 | 493.1712, 401.8835, 361.1288, 175.0246, 153.0917, 113.0240 | C25H30O13 | Ligstroside-aglycone glucuronide | 1, 2, 4, 8 | 2.75 × 10−3 |
| Urine | ||||||||
| 232.0048 | 8.23 | −1.6 | 98.03 | 203.2221, 151.0399, 108.0213 | C8H8O6S | 3,4-dihydroxyphenylacetaldehyde (DOPAL)-sulfate * | 8, 24 | 1.25 × 10−2 |
| 368.1114 | 8.39 | −0.63 | 98.69 | 193.0505,134.0376 | C17H20O9 | Ferulic acid derivative | 8, 24 | 1.98 × 10−3 |
| 274.0152 | 10.98 | −1.71 | 97.52 | 193.0508, 149.0243, 121.0285, 93.0347, 65.0396 | C10H10O7S | Ferulic acid sulfate | 8, 24 | 2.24 × 10−3 |
| 538.1688 | 14.65 | −0.02 | 99.04 | 493.1712, 401.8835, 361.1288, 175.0246, 153.0917, 113.0240 | C25H30O13 | Ligstroside-aglycone glucuronide | 8, 24 | 1.08 × 10−2 |
3. Experimental Section
3.1. Chemicals
3.2. Fraxinus Extract Characterization
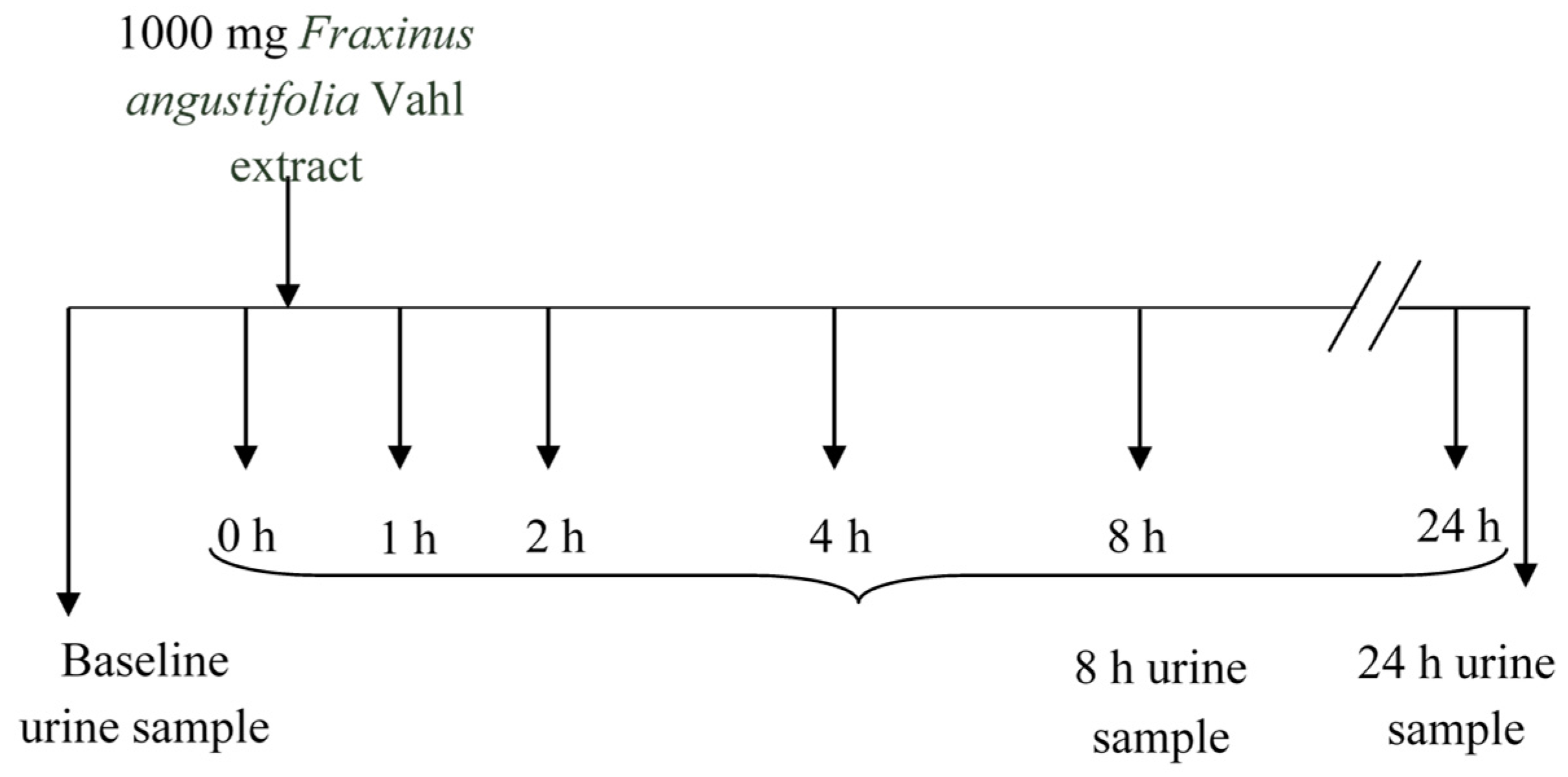
3.3. Human Intervention Study
| N | 10 |
|---|---|
| Gender (M/F) | (4/6) |
| Age (years old) | 23.5 ± 1.3 |
| Weight (kg) | (M) 81.0 ± 12.0 |
| (F) 69.1 ± 7.2 | |
| BMI (kg/m2) | (M) 25.1 ± 2.8 |
| (F) 21.5 ± 2.1 |

3.4. Urine and Plasma Samples Preparation
3.5. UPLC-ESI-QTOF MS Analysis of Plasma and Urine Samples
3.5.1. Targeted Metabolomics Analysis
3.5.2. Untargeted Metabolomics Analysis Steps
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kostova, I.; Iossifova, T. Chemical components of Fraxinus species. Fitoterapia 2007, 78, 85–106. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Garcia, F.; Flanagan, J.; García-Molina, O.; Vilaplana-Vivo, V.; García-Carrillo, N.; Fança-Berthon, P.; Bily, A.; Roller, M.; Vicente-Ortega, V.; Issaly, N. Preventive effect of a Fraxinus excelsior L seeds/fruits extract on hepatic steatosis in obese type 2 diabetic mice. J. Diabetes Metab. 2015, 6. [Google Scholar] [CrossRef]
- Montó, F.; Arce, C.; Noguera, M.A.; Ivorra, M.D.; Flanagan, J.; Roller, M.; Issaly, N.; D’Ocon, P. Action of an extract from the seeds of Fraxinus excelsior L. on metabolic disorders in hypertensive and obese animal models. Food Funct. 2014, 5, 786–796. [Google Scholar] [CrossRef] [PubMed]
- Ibarra, A.; Baia, N.; Hea, K.; Bily, A.; Cases, J.; Roller, M.; Sang, S. Fraxinus excelsior seed extract FraxiPure™ limits weight gains and hyperglycemia in high-fat diet-induced obese mice. Phytomedicine 2011, 18, 479–485. [Google Scholar] [CrossRef] [PubMed]
- López-Carreras, N.; Fernández-Vallinas, S.; Miguel, M.; Aleixandre, A. Long-term effect of an aqueous Fraxinus excelsior L. seed extract in spontaneously hypertensive rats. Int. J. Hypertens. 2014. [Google Scholar] [CrossRef] [PubMed]
- Visen, P.; Saraswat, B.; Visen, A.; Roller, M.; Bily, A.; Mermet, C.; He, K.; Bai, N.; Lemaire, B.; Lafay, S.; et al. Acute effects of Fraxinus excelsior L. seed extract on postprandial glycemia and insulin secretion on healthy volunteers. J. Ethnopharmacol. 2009, 126, 226–232. [Google Scholar] [CrossRef] [PubMed]
- Zulet, M.A.; Navas-Carretero, S.; Lara y Sánchez, D.; Abete, I.; Flanagan, J.; Issaly, N.; Fanca-Berthon, P.; Bily, A.; Martinez, J.A. A Fraxinus excelsior L. seeds/fruits extract benefits glucose homeostasis and adiposity related markers in elderly overweight/obese subjects: A longitudinal, randomized, crossover, double-blind, placebo-controlled nutritional intervention study. Phytomedicine 2014, 21, 1162–1169. [Google Scholar] [CrossRef] [PubMed]
- Flanagan, J.; Meyer, M.; Pasamar, M.A.; Ibarra, A.; Roller, M.; Alvarez i Genoher, N.; Leiva, S.; Gómez-García, F.; Alcaraz, M.; Martínez-Carrasco, A.; et al. Safety evaluation and nutritional composition of a Fraxinus excelsior seed extract, FraxiPure™. Food Chem. Toxicol. 2013, 53, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Bai, N.; He, K.; Ibarra, A.; Bily, A.; Roller, M.; Chen, X.; Rühl, R. Iridoids from Fraxinus excelsior with adipocyte differentiation-inhibitory and PPARα activation activity. J. Nat. Prod. 2010, 73, 2–6. [Google Scholar] [CrossRef] [PubMed]
- Suárez, M.; Valls, R.M.; Romero, M.P.; Maciá, A.; Fernández, S.; Giralt, M.; Solá, R.; Motilva, M.J. Bioavailability of phenols from a phenol-enriched olive oil. Br. J. Nutr. 2011, 106, 1691–1701. [Google Scholar] [CrossRef] [PubMed]
- García-Villalba, R.; Carrasco-Pancorbo, A.; Nevedomskaya, E.; Mayboroda, O.A.; Deelder, A.M.; Segura-Carretero, A.; Fernández-Gutiérrez, A. Exploratory analysis of human urine by LC-ESI-TOF MS after high intake of olive oil: Understanding the metabolism of polyphenols. Anal. Bioanal. Chem. 2010, 398, 463–475. [Google Scholar] [CrossRef] [PubMed]
- Pinto, J.; Paiva-Martins, F.; Corona, G.; Debnam, E.S.; Oruna-Concha, M.J.; Vauzour, D.; Gordon, M.H.; Spencer, J.P.E. Absorption and metabolism of olive oil secoiridoids in the small intestine. Br. J. Nutr. 2011, 105, 1607–1618. [Google Scholar] [CrossRef] [PubMed]
- De Bock, M.; Thorstensen, E.B.; Derraik, J.G.B.; Henderson, H.V.; Hofman, P.L.; Cutfield, W.S. Human absorption and metabolism of oleuropein and hydroxytyrosol ingested as olive (Olea europaea L.) leaf extract. Mol. Nutr. Food Res. 2013, 57, 2079–2085. [Google Scholar] [CrossRef] [PubMed]
- Lockyer, S.; Corona, G.; Yaqoob, P.; Spencer, J.P.E.; Rowland, I. Secoiridoids delivered as olive leaf extract induce acute improvements in human vascular function and reduction of an inflammatory cytokine: A randomized, double-blind, placebo-controlled, cross-over trial. Br. J. Nutr. 2015. [Google Scholar] [CrossRef] [PubMed]
- Kendall, M.; Batterham, M.; Callahan, D.L.; Jardine, D.; Prenzler, P.D.; Robards, K.; Ryan, D. Randomized controlled study of the urinary excretion of biophenols following acute and chronic intake of olive leaf supplements. Food Chem. 2012, 130, 651–659. [Google Scholar] [CrossRef]
- García-Villalba, R.; Larrosa, M.; Possemiers, S.; Tomás-Barberán, F.A.; Espín, J.C. Bioavailability of phenolics from an oleuropein-rich olive (Olea europaea) leaf extract and its acute effect on plasma antioxidant status: Comparison between pre- and postmenopausal women. Eur. J. Nutr. 2014, 53, 1015–1027. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Kim, J.; Yun, E.J.; Kim, K.H. Food metabolomics: From farm to human. Curr. Opin. Biotechnol. 2016, 37, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Primrose, S.; Draper, J.; Elsom, R.; Kirkpatrick, V.; Mathers, J.C.; Seal, C.; Beckmann, M.; Haldar, S.; Beattie, J.H.; Lodge, J.K.; et al. Metabolomics and human nutrition. Br. J. Nutr. 2011, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Nuñez-Sanchez, M.A.; García-Villalba, R.; Monedero-Saiz, T.; García-Talavera, N.V.; Gómez-Sánchez, M.B.; Sánchez-Álvarez, C.; García-Albert, A.M.; Rodríguez-Gil, F.J.; Ruiz-Marín, M.; Pastor-Quirante, F.A.; et al. Targeted metabolic profiling of pomegranate polyphenols and urolithins in plasma, urine and colon tissues from colorectal cancer patients. Mol. Nutr. Food Res. 2014, 58, 1199–1211. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, M.; Engelsen, S.B.; Dragsted, L.O. LC-MS metabolomics top-down approach reveals new exposure and effect biomarkers of apple and apple-pectin intake. Metabolomics 2012, 8, 64–73. [Google Scholar] [CrossRef]
- Xie, G.; Zhao, A.; Zhao, L.; Chen, T.; Chen, H.; Qi, X.; Zheng, X.; Ni, Y.; Cheng, Y.; Lan, K.; et al. Metabolic fate of tea polyphenols in humans. J. Proteome Res. 2012, 11, 3449–3457. [Google Scholar] [CrossRef] [PubMed]
- Llorach, R.; Garrido, I.; Monagas, M.; Urpi-Sarda, M.; Tulipani, S.; Bartolome, B.; Andres-Lacueva, C. Metabolomics study of human urinary metabolome modifications after intake of almond (Prunus dulcis (Mill.) D.A. Webb) skin polyphenols. J. Proteome Res. 2010, 9, 5859–5867. [Google Scholar] [CrossRef] [PubMed]
- Silva, S.; Gomes, L.; Leitao, F.; Bronze, M.; Coelho, A.V.; Vilas-Boas, L. Secoirioids in olive seed: Characterization of nuzhenide and 11-methyl oleosides by liquid chromatography with diode array and mass spectrometry. Grasas Aceites 2010, 61, 157–164. [Google Scholar] [CrossRef]
- Del Boccio, P.; di Deo, A.; de Curtis, A.; Celli, N.; Iacoviello, L.; Rotilio, D. Liquid chromatography-tandem mass spectrometry analysis of oleuropein and its metabolite hydroxytyrosol in rat plasma and urine after oral administration. J. Chromatogr. B 2003, 785, 47–56. [Google Scholar] [CrossRef]
- Guo, N.; Zhu, M.; Han, X.; Sui, D.; Wang, Y.; Yang, Q. The metabolism of salidroside to its aglycone p-tyrosol in rats following the administration of salidroside. PLoS ONE 2014, 9, e103648. [Google Scholar] [CrossRef] [PubMed]
- Akao, T.; Kobashi, K.; Aburada, M. Enzymic studies on the animal and intestinal bacterial metabolism of geniposide. Biol. Pharm. Bull. 1994, 17, 1573–1576. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Mateos, A.; Cifuentes-Gomez, T.; Gonzalez-Salvador, I.; Ottaviani, J.I.; Schroeter, H.; Kelm, M.; Heiss, C.; Spencer, J. Influence of age on the absorption, metabolism, and excretion of cocoa flavanols in healthy subjects. Mol. Nutr. Food Res. 2015, 1–9. [Google Scholar] [CrossRef] [PubMed]
- García-Conesa, M.T. Dietary polyphenols against metabolic disorders: How far have we progressed in the understanding of the molecular mechanisms of action of these compounds? Crit. Rev. Food Sci. Nutr. 2015. [Google Scholar] [CrossRef] [PubMed]
- Mosele, J.I.; Martín-Peláez, S.; Maciá, A.; Farrás, M.; Valls, R.M.; Catalán, U.; Motilva, M.J. Faecal microbial metabolism of olive oil phenolic compounds: In vitro and in vivo approaches. Mol. Nutr. Food Res. 2014, 58, 1809–1819. [Google Scholar] [CrossRef] [PubMed]
- D’Angelo, S.; Manna, C.; Migliardi, V.; Mazzoni, O.; Morrica, P.; Capasso, G.; Pontoni, G.; Galleti, P.; Zappia, V. Pharmacokinetics and metabolism of hydroxytyrosol, a natural antioxidant from olive oil. Drug Metab. Dispos. 2001, 29, 1492–1498. [Google Scholar] [PubMed]
- Chandramohan, R.; Pari, L.; Rathinam, A.; Ahmad Sheikh, B. Tyrosol, a phenolic compound, ameliorates hyperglycemia by regulating key enzymes of carbohydrate metabolism in streptozotocin induced diabetic rats. Chem. Biol. Interact. 2015, 229, 44–54. [Google Scholar] [CrossRef] [PubMed]
- Bohn, T. Dietary factors affecting polyphenol bioavailability. Nutr. Rev. 2014, 72, 429–452. [Google Scholar] [CrossRef] [PubMed]
- Romo-Vaquero, M.; Selma, M.V.; Larrosa, M.; Obiol, M.; García-Villalba, R.; González-Barrio, R.; Issaly, N.; Flanagan, J.; Roller, M.; Tomás-Barberán, F.A.; et al. A rosemary extract rich in carnosic acid selectively modulates caecum microbiota and inhibits β-glucosidase activity, altering fiber and short chain fatty acids fecal excretion in lean and obese female rats. PLoS ONE 2014, 9, e94687. [Google Scholar] [CrossRef] [PubMed]
- He, K.; Roller, M.; Bily, A.; Bai, N.; Dikansky, J.; Ibarra, A. Extract of Fraxinus Excelsior Seeds and Therapeutic Applications Therefor. Patent Number US 8293292 B2, 23 November 2012. [Google Scholar]
- Del Rio, D.; Rodriguez-Mateos, A.; Spencer, J.P.E.; Tognolini, M.; Borges, G.; Crozier, A. Dietary (poly) phenolics in human health: Structures, bioavailability, and evidence of protective effects against chronic diseases. Antioxid. Redox Signal 2013, 18, 1818–1892. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the compounds are available from the Naturex´s authors.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Villalba, R.; Tomás-Barberán, F.A.; Fança-Berthon, P.; Roller, M.; Zafrilla, P.; Issaly, N.; García-Conesa, M.-T. Targeted and Untargeted Metabolomics to Explore the Bioavailability of the Secoiridoids from a Seed/Fruit Extract (Fraxinus angustifolia Vahl) in Human Healthy Volunteers: A Preliminary Study. Molecules 2015, 20, 22202-22219. https://doi.org/10.3390/molecules201219845
García-Villalba R, Tomás-Barberán FA, Fança-Berthon P, Roller M, Zafrilla P, Issaly N, García-Conesa M-T. Targeted and Untargeted Metabolomics to Explore the Bioavailability of the Secoiridoids from a Seed/Fruit Extract (Fraxinus angustifolia Vahl) in Human Healthy Volunteers: A Preliminary Study. Molecules. 2015; 20(12):22202-22219. https://doi.org/10.3390/molecules201219845
Chicago/Turabian StyleGarcía-Villalba, Rocío, Francisco A. Tomás-Barberán, Pascale Fança-Berthon, Marc Roller, Pilar Zafrilla, Nicolas Issaly, and María-Teresa García-Conesa. 2015. "Targeted and Untargeted Metabolomics to Explore the Bioavailability of the Secoiridoids from a Seed/Fruit Extract (Fraxinus angustifolia Vahl) in Human Healthy Volunteers: A Preliminary Study" Molecules 20, no. 12: 22202-22219. https://doi.org/10.3390/molecules201219845
APA StyleGarcía-Villalba, R., Tomás-Barberán, F. A., Fança-Berthon, P., Roller, M., Zafrilla, P., Issaly, N., & García-Conesa, M. -T. (2015). Targeted and Untargeted Metabolomics to Explore the Bioavailability of the Secoiridoids from a Seed/Fruit Extract (Fraxinus angustifolia Vahl) in Human Healthy Volunteers: A Preliminary Study. Molecules, 20(12), 22202-22219. https://doi.org/10.3390/molecules201219845