
Purification and Characterization of Chitinases from Ridgetail White Prawn Exopalaemon carinicauda

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussions
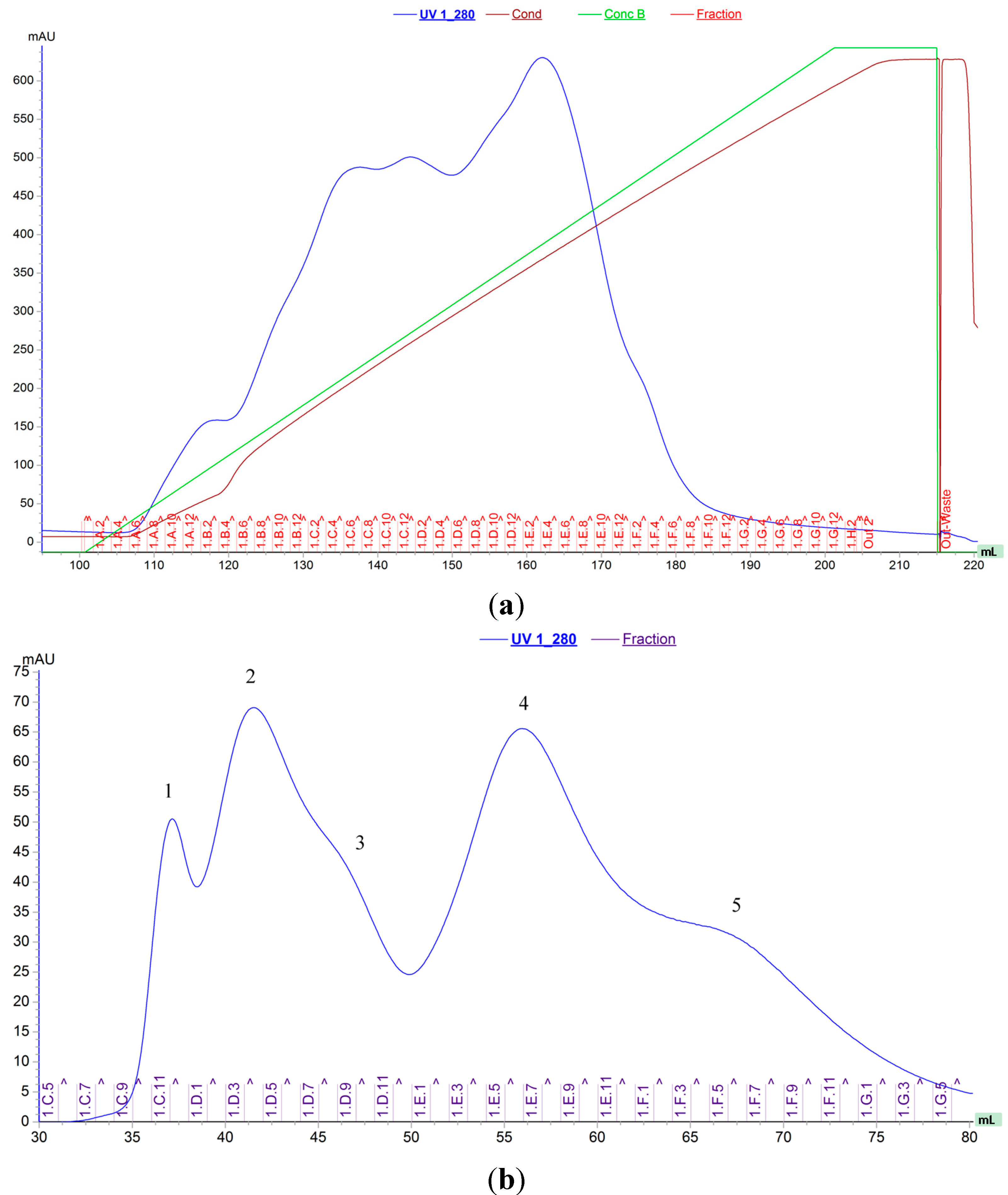
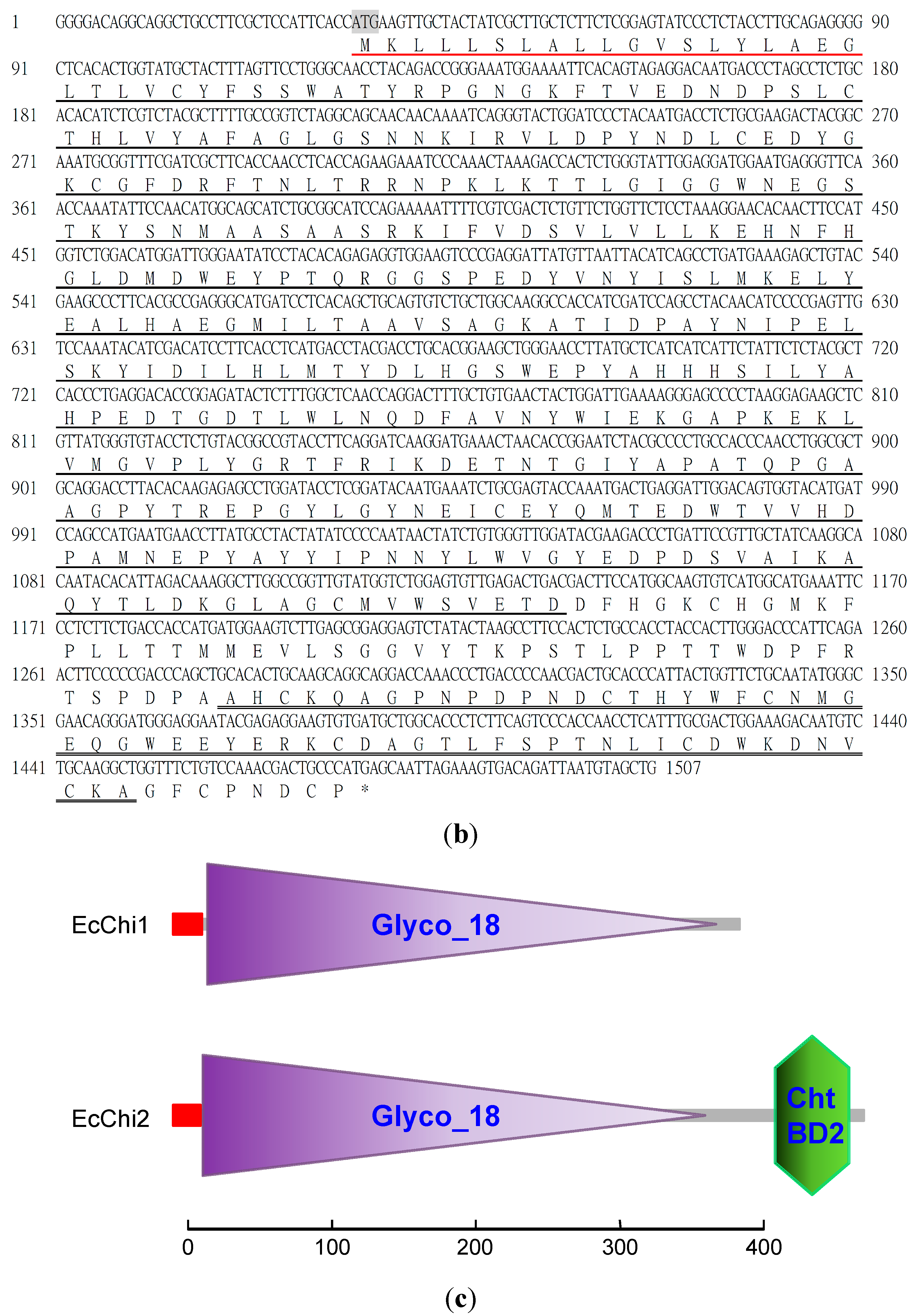
2.1. Protein Purification, Molecular Weight Determination and Amino Acid Sequence




2.2. Chitinolytic Activity, Protein Concentration and the Specific Activity
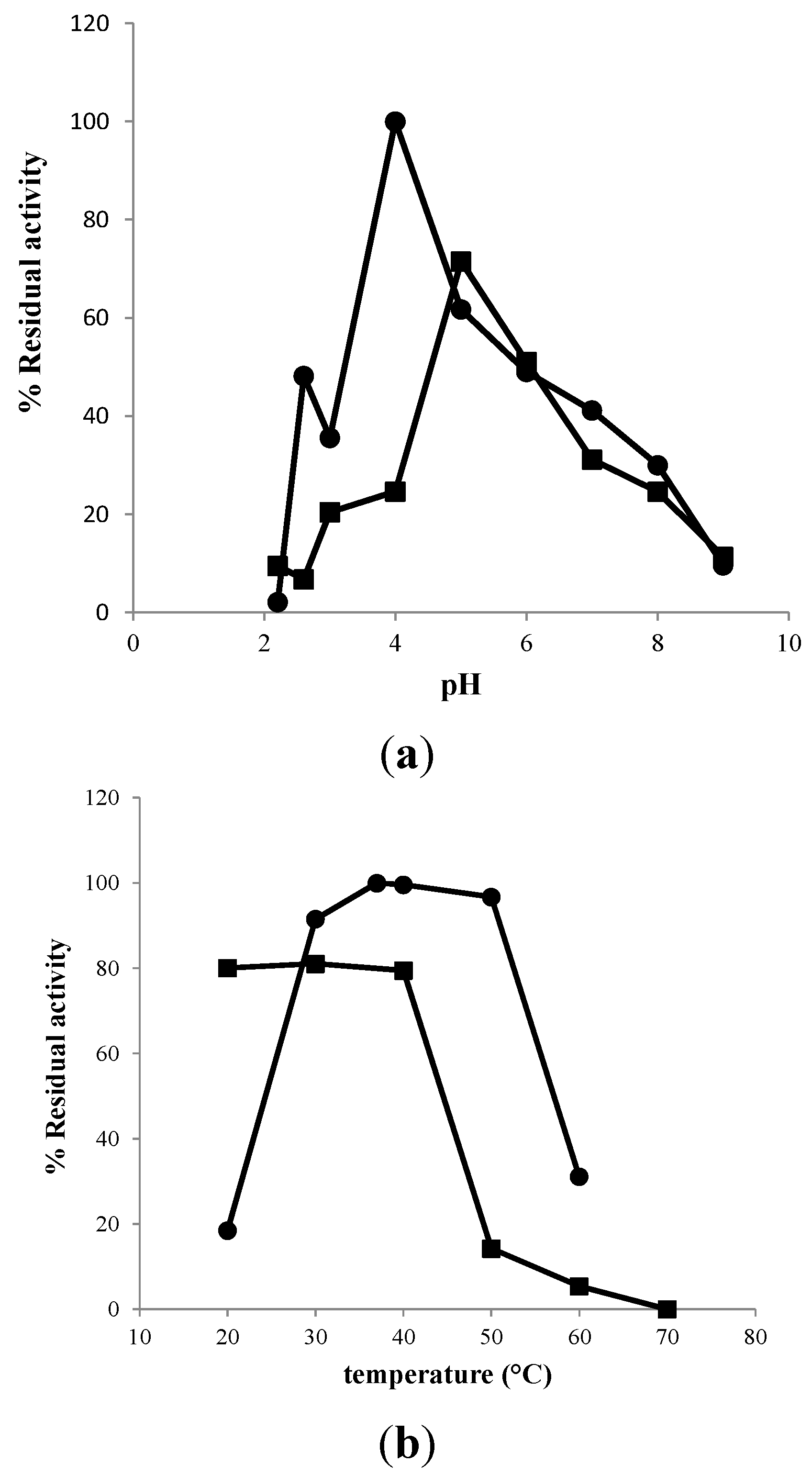
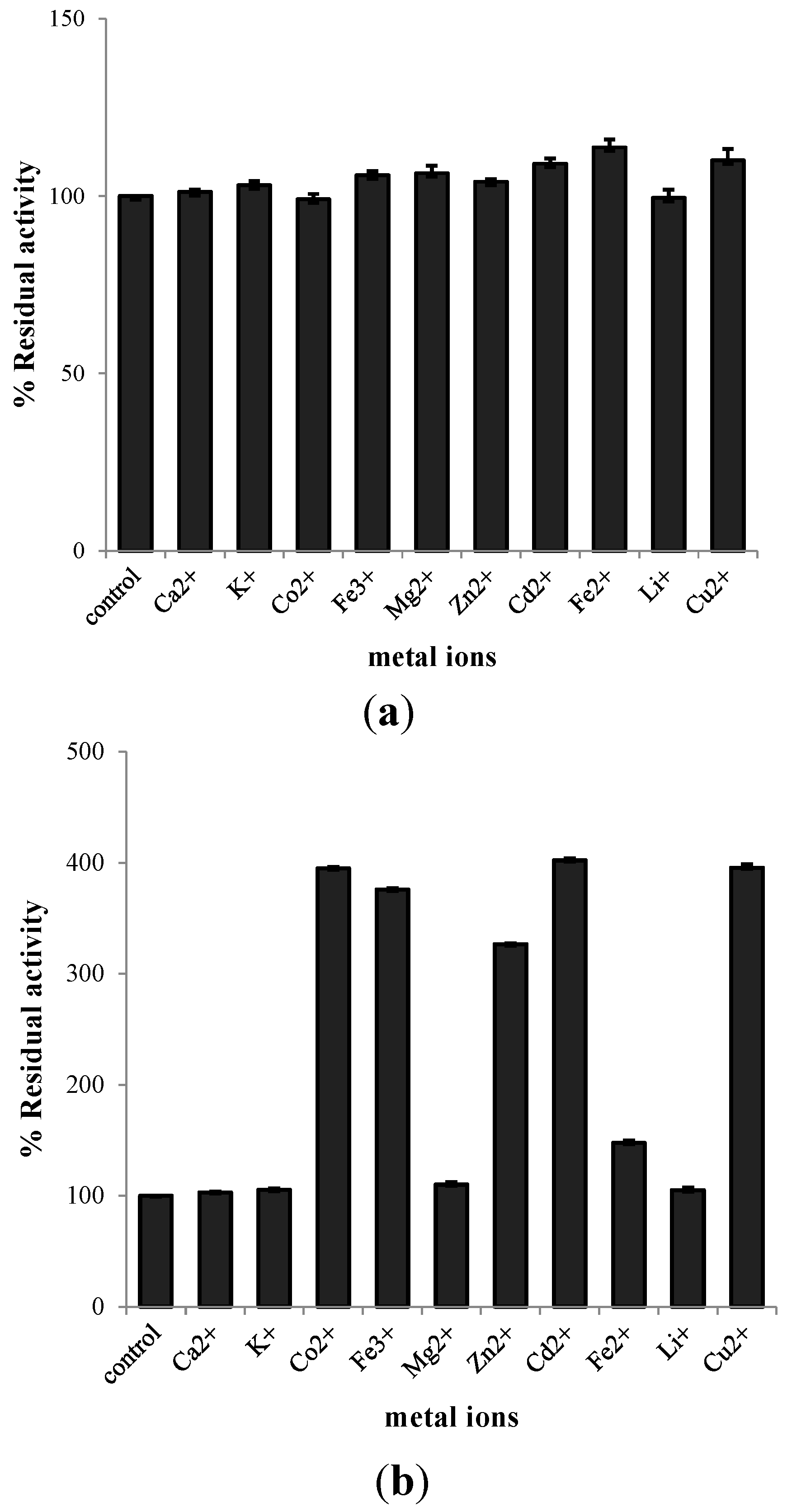
2.3. Characterization of Purified EcChi1


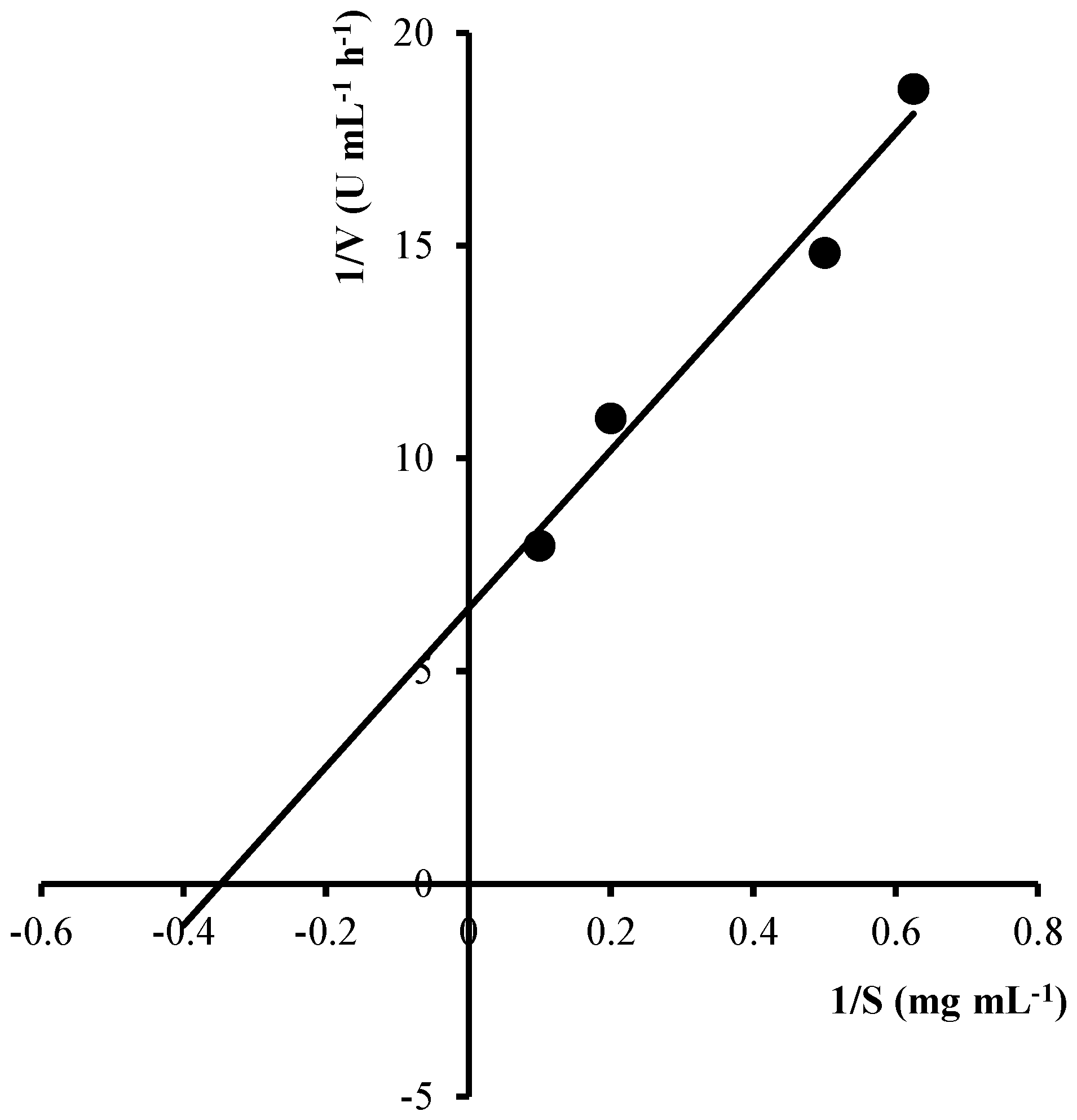
2.4. Kinetic Parameters for Colloidal Chitin Hydrolysis by EcChi1

3. Experimental Section
3.1. Materials
3.2. Chitinolytic Activity Assay
3.3. Protein Concentration Determination
3.4. Isolation and Purification of Chitinases from E. carinicauda (EcChis)
3.5. Identification of Purified EcChis by LC-ESI-MS/MS
3.6. Effect of pH Value on Chitinolytic Activity
3.7. Effect of Temperature on Chitinolytic Activity
3.8. Effect of Metal Ions on Chitinolytic Activity
3.9. Kinetic Parameters for Colloidal Chitin Hydrolysis by EcChi1
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Patil, R.S.; Ghormade, V.; Deshpande, M.V. Chitinolytic enzymes: An exploration. Enzym. Microb. Technol. 2000, 26, 473–483. [Google Scholar] [CrossRef]
- Rinaudo, M. Chitin and chitosan: Properties and applications. Prog. Polym. Sci. 2006, 31, 603–632. [Google Scholar] [CrossRef]
- Chen, J.K.; Shen, C.R.; Liu, C.L. N-acetylglucosamine: Production and applications. Mar. Drugs 2010, 8, 2493–2516. [Google Scholar] [CrossRef] [PubMed]
- Aam, B.B.; Heggset, E.B.; Norberg, A.L.; Sorlie, M.; Varum, K.M.; Eijsink, V.G. Production of chitooligosaccharides and their potential applications in medicine. Mar. Drugs 2010, 8, 1482–1517. [Google Scholar] [CrossRef] [PubMed]
- Xia, W.; Liu, P.; Zhang, J.; Chen, J. Biological activities of chitosan and chitooligosaccharides. Food Hydrocoll. 2011, 25, 170–179. [Google Scholar] [CrossRef]
- Gorbach, V.I.; Krasikova, I.N.; Luk’yanov, P.A.; Loenko, Y.N.; Solov’eva, T.F.; Ovodov, Y.S.; Deev, V.V.; Pimenov, A.A. New glycolipids (chitooligosaccharide derivatives) possessing immunostimulating and antitumor activities. Carbohydr. Res. 1994, 260, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Shen, K.T.; Chen, M.H.; Chan, H.Y.; Jeng, J.H.; Wang, Y.J. Inhibitory effects of chitooligosaccharides on tumor growth and metastasis. Food Chem. Toxicol. 2009, 47, 1864–1871. [Google Scholar] [CrossRef] [PubMed]
- Dahiya, N.; Tewari, R.; Hoondal, G.S. Biotechnological aspects of chitinolytic enzymes: A review. Appl. Microbiol. Biotechnol. 2006, 71, 773–782. [Google Scholar] [CrossRef] [PubMed]
- Aloise, P.; Lumme, M.; Haynes, C. N-acetyl-D-glucosamine production from chitin-waste using chitinases from Serratia marcescens. Chitin Enzymol. 1996, 2, 581–594. [Google Scholar]
- Merzendorfer, H.; Zimoch, L. Chitin metabolism in insects: Structure, function and regulation of chitin synthases and chitinases. J. Exp. Biol. 2003, 206, 4393–4412. [Google Scholar] [CrossRef] [PubMed]
- Merzendorfer, H. Insect chitin synthases: A review. J. Comp. Physiol. B 2006, 176, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Gooday, G. Diversity of roles of chitinases in nature. In Chitin and Chitosan; Abdullah, M.P., Zakaria, M.B., Wan Muda, W.M., Eds.; Penerbit Universiti Kebangsaan: Bangi, Malaysia, 1995; Volume 3, pp. 191–202. [Google Scholar]
- Flach, J.; Pilet, P.-E.; Jolles, P. What’s new in chitinase research? Experientia 1992, 48, 701–716. [Google Scholar] [CrossRef] [PubMed]
- Sahai, A.; Manocha, M. Chitinases of fungi and plants: Their involvement in morphogenesis and host-parasite interaction. FEMS Microbiol. Rev. 1993, 11, 317–338. [Google Scholar] [CrossRef]
- Kombrink, E.; Somssich, I.E. Defense responses of plants to pathogens. Adv. Bot. Res. 1995, 21, 1–34. [Google Scholar]
- Kramer, K.J.; Muthukrishnan, S. Insect chitinases: Molecular biology and potential use as biopesticides. Insect Biochem. Mol. Biol. 1998, 27, 887–900. [Google Scholar] [CrossRef]
- Wang, S.L.; Chang, W.T. Purification and characterization of two bifunctional chitinases/lysozymes extracellularly produced by Pseudomonas aeruginosa k-187 in a shrimp and crab shell powder medium. Appl. Environ. Microbiol. 1997, 63, 380–386. [Google Scholar] [PubMed]
- Jeuniaux, C. Chitine et chitinolyse: Un chapitre de la Biologie Moléculaire, 1st ed.; Florkin, M., Ed.; Masson: Paris, France, 1963; Volume 3, p. 181. [Google Scholar]
- Elyakova, L. Distribution of cellulases and chitinases in marine invertebrates. Comp. Biochem. Physiol. Part B 1972, 43, 67–70. [Google Scholar]
- Funke, B.; Spindler, K. Developmental changes of chitinolytic enzymes and ecdysteroid levels during the early development of the brine shrimp Artemia. Artemia Res. Appl. 1987, 2, 67–68. [Google Scholar]
- Funke, B.; Spindler, K.D. Characterization of chitinase from the brine shrimp Artemia. Comp. Biochem. Physiol. Part B 1989, 94, 691–695. [Google Scholar] [CrossRef]
- Spindler-Barth, M.; van Wormhoudt, A.; Spindler, K.-D. Chitinolytic enzymes in the integument and midgut-gland of the shrimp Palaemon serratus during the moulting cycle. Mar. Biol. 1990, 106, 49–52. [Google Scholar] [CrossRef]
- Zou, E.; Bonvillain, R. Chitinase activity in the epidermis of the fiddler crab, Uca pugilator, as an in vivo screen for molt-interfering xenobiotics. Comp. Biochem. Physiol. Part C 2004, 139, 225–230. [Google Scholar] [CrossRef]
- Watanabe, T.; Kono, M.; Aida, K.; Nagasawa, H. Isolation of cDNA encoding a putative chitinase precursor in the kuruma prawn Penaeus japonicus. Mol. Mar. Biol. Biotechnol. 1996, 5, 299–303. [Google Scholar] [PubMed]
- Watanabe, T.; Kono, M. Isolation of a cDNA encoding a chitinase family protein from cuticular tissues of the kuruma prawn Penaeus japonicus. Zool. Sci. 1997, 14, 65–68. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Kono, M.; Aida, K.; Nagasawa, H. Purification and molecular cloning of a chitinase expressed in the hepatopancreas of the penaeid prawn Penaeus japonicus. BBA-Protein Struct. Mol. Enzymol. 1998, 1382, 181–185. [Google Scholar] [CrossRef]
- Tan, S.H.; Degnan, B.M.; Lehnert, S.A. The Penaeus monodon chitinase 1 gene is differentially expressed in the hepatopancreas during the molt cycle. Mar. Biotechnol. 2000, 2, 126–135. [Google Scholar] [PubMed]
- Proespraiwong, P.; Tassanakajon, A.; Rimphanitchayakit, V. Chitinases from the black tiger shrimp Penaeus monodon: Phylogenetics, expression and activities. Comp. Biochem. Physiol. Part B 2010, 156, 86–96. [Google Scholar] [CrossRef]
- Lehnert, S.A.; Johnson, S.E. Expression of hemocyanin and digestive enzyme messenger RNAs in the hepatopancreas of the black tiger shrimp Penaeus monodon. Comp. Biochem. Physiol. Part B 2002, 133, 163–171. [Google Scholar] [CrossRef]
- Zhang, J.; Sun, Y.; Li, F.; Huang, B.; Xiang, J. Molecular characterization and expression analysis of chitinase (Fcchi-3) from Chinese shrimp, Fenneropenaeus chinensis. Mol. Biol. Rep. 2010, 37, 1913–1921. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.S.; Yan, J.H.; Tang, J.Y.; Tao, Y.M.; Xie, X.L.; Wang, Y.; Wei, X.Q.; Yan, Q.H.; Chen, Q.X. Cloning and tissue expressions of seven chitinase family genes in Litopenaeus vannamei. Fish Shellfish Immunol. 2010, 29, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Salma, U.; Uddowla, M.H.; Kim, M.; Kim, J.M.; Kim, B.K.; Baek, H.-J.; Park, H.; Mykles, D.L.; Kim, H.-W. Five hepatopancreatic and one epidermal chitinases from a pandalid shrimp (Pandalopsis japonica): Cloning and effects of eyestalk ablation on gene expression. Comp. Biochem. Physiol. Part B 2012, 161, 197–207. [Google Scholar] [CrossRef]
- Kono, M.; Matsui, T.; Shimizu, C.; Koga, D. Purifications and some properties of chitinase from the liver of a prawn, Penaeus japonicus (biological chemistry). Agric. Biol. Chem. 1990, 54, 2145–2147. [Google Scholar] [CrossRef]
- Sotelo-Mundo, R.R.; Moran-Palacio, E.F.; Garcia-Orozco, K.D.; Figueroa-Soto, C.; Yepiz-Plascencia, G.; Romo-Figueroa, M.G.; Valenzuela-Soto, E.M. Kinetic characterization, expression and molecular modeling of a chitinase from the pacific white shrimp Litopenaeus vannamei. J. Food Biochem. 2009, 33, 246–259. [Google Scholar]
- Spindler, K.D.; Buchholz, F. Partial characterization of chitin degrading enzymes from two euphausiids, Euphausia superba and Meganyctiphanes norvegica. Polar Biol. 1988, 9, 115–122. [Google Scholar] [CrossRef]
- Rodriguez-Kabana, R.; Godoy, G.; Morgan-Jones, G.; Shelby, R. The determination of soil chitinase activity: Conditions for assay and ecological studies. Plant Soil 1983, 75, 95–106. [Google Scholar] [CrossRef]
- Reissig, J.L.; Strominger, J.L.; Leloir, L.F. A modified colorimetric method for the estimation of N-acetylamino sugars. J. Biol. Chem. 1955, 217, 959–966. [Google Scholar] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Zhang, X.; Zhang, J.; Li, F.; Zhang, J.; Xiang, J. Comparative transcriptome analysis on embryo and adults of Exopalamon carincauda. In Experimental Biology, Proceedings of the 8th Symposium of World’s Chinese Scientist’s on Crustacean Aquaculture, Ningbo, China, 16–18 November 2012; Qingdao Publishing House: Qingdao, China, 2012. Abstract Number (157), Pagination (207). [Google Scholar]
- Sample Availability: Samples described in this publication are available upon request in a timely manner for noncommercial research purposes.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Zhang, J.; Song, F.; Gui, T.; Xiang, J. Purification and Characterization of Chitinases from Ridgetail White Prawn Exopalaemon carinicauda. Molecules 2015, 20, 1955-1967. https://doi.org/10.3390/molecules20021955
Wang J, Zhang J, Song F, Gui T, Xiang J. Purification and Characterization of Chitinases from Ridgetail White Prawn Exopalaemon carinicauda. Molecules. 2015; 20(2):1955-1967. https://doi.org/10.3390/molecules20021955
Chicago/Turabian StyleWang, Jing, Jiquan Zhang, Fengge Song, Tianshu Gui, and Jianhai Xiang. 2015. "Purification and Characterization of Chitinases from Ridgetail White Prawn Exopalaemon carinicauda" Molecules 20, no. 2: 1955-1967. https://doi.org/10.3390/molecules20021955
APA StyleWang, J., Zhang, J., Song, F., Gui, T., & Xiang, J. (2015). Purification and Characterization of Chitinases from Ridgetail White Prawn Exopalaemon carinicauda. Molecules, 20(2), 1955-1967. https://doi.org/10.3390/molecules20021955