
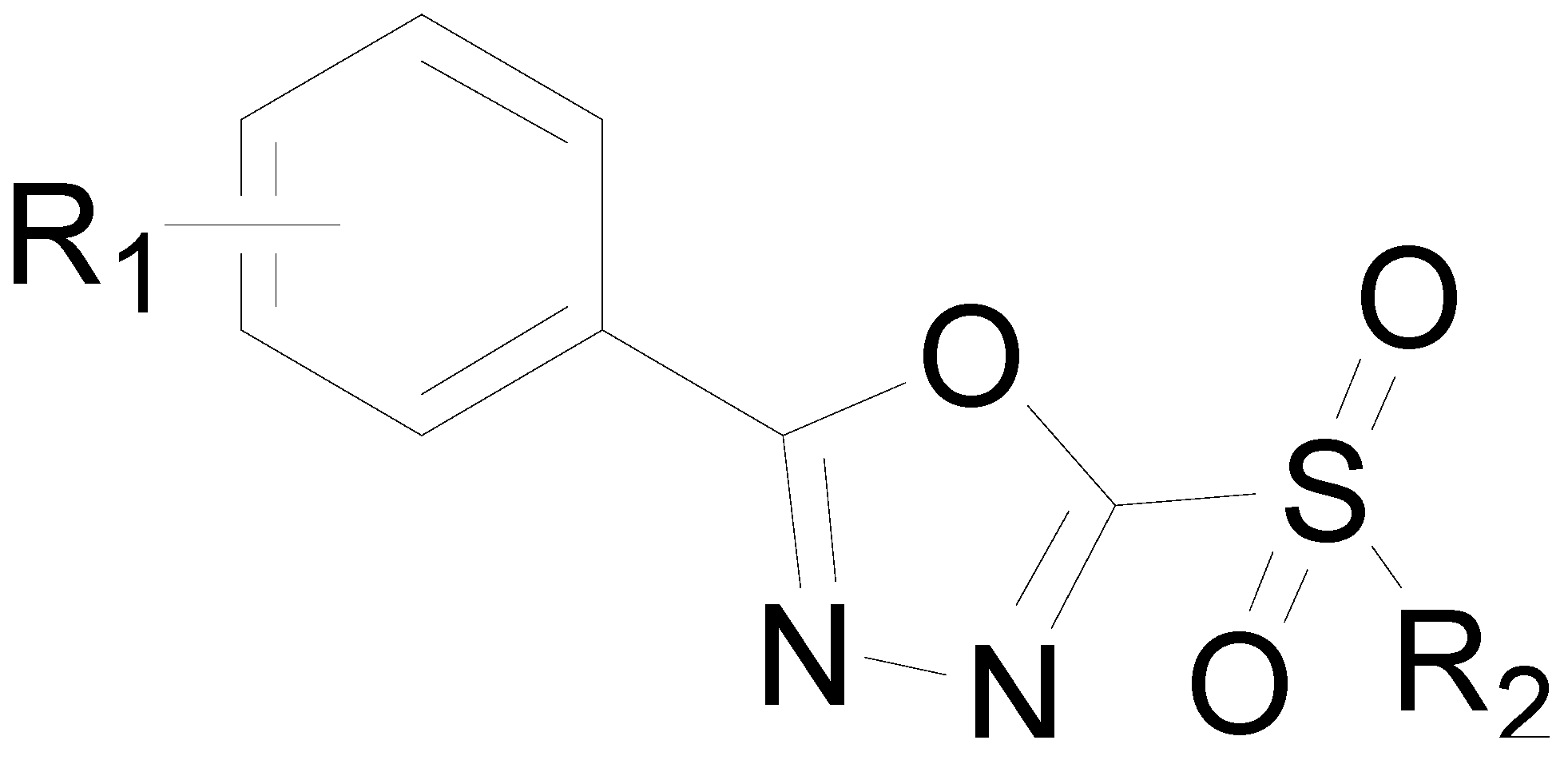
Antibacterial Activity and Mechanism of Action of Sulfone Derivatives Containing 1,3,4-Oxadiazole Moieties on Rice Bacterial Leaf Blight


Abstract
:1. Introduction

2. Results and Discussion
2.1. In Vitro Antibacterial Bioassay

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compds. | Toxic Regression Equation | R | EC50 (μg/mL) |
|---|---|---|---|
| 2-(Methylsulfonyl)-5-phenyl-1,3,4-oxadiazole | y = 2.16x + 2.18 | 0.98 | 20.07 ± 1.21 |
| 2-(Ethylsulfonyl)-5-phenyl-1,3,4-oxadiazole | y = 1.52x + 2.77 | 0.98 | 29.00 ± 1.25 |
| 2-(Methylsulfonyl)-5-(4-fluorophenyl)-1,3,4-oxadiazole | y = 4.13x + 0.89 | 0.95 | 9.89 ± 1.52 |
| 2-(Ethylsulfonyl)-5-(4-fluorophenyl)-1,3,4-oxadiazole | y = 3.28x + 1.61 | 0.96 | 10.80 ± 1.43 |
| 2-(Methylsulfonyl)-5-(4-chlorophenyl)-1,3,4-oxadiazole | y = 1.72x + 2.64 | 0.99 | 23.21 ± 0.98 |
| 2-(Ethylsulfonyl)-5-(4-chlorophenyl)-1,3,4-oxadiazole | y = 1.60x + 2.25 | 0.99 | 52.61 ± 1.08 |
| 2-(Methylsulfonyl)-5-(2,4-dichlorophenyl)-1,3,4-oxadiazole | y = 1.04x + 3.21 | 0.99 | 52.14 ± 1.05 |
| 2-(Ethylsulfonyl)-5-(2,4-dichlorophenyl)-1,3,4-oxadiazole | y = 1.43x + 2.42 | 0.97 | 63.95 ± 1.05 |
| Bismerthiazole | y = 1.50x + 2.05 | 0.98 | 92.61 ± 2.15 |
| Thiodiazole copper | y = 1.54x + 1.79 | 0.98 | 121.82 ± 3.59 |
2.2. In Vivo Antibacterial Bioassay
| Compds. | 15 Days after Spraying | 28 Days after Spraying | ||
|---|---|---|---|---|
| Disease Index (%) | Curative Activity (%) c | Disease Index (%) | Protection Activity (%) c | |
| 2-(Methylsulfonyl)-5-(4-fluorophenyl)-1,3,4-oxadiazole | 57.63 ± 3.51 | 38.17 ± 2.15 A | 54.79 ± 2.78 | 41.82 ± 2.45 A |
| Bismerthiazole a | 65.05 ± 2.26 | 30.21 ± 3.43 B | 58.85 ± 3.12 | 37.51 ± 2.54 C |
| Thiodiazole copper b | 65.70 ± 2.73 | 29.51 ± 4.76 C | 70.08 ± 3.67 | 25.58 ± 2.42 B |
| Untreated blank control | 93.21 ± 1.79 | 0 | 94.17 ± 2.55 | 0 |
2.3. Field Trial against Rice Bacterial Leaf Blight
| Compds. | Dosage (g ai/ha) d | 15 Days after the Third Spraying | |
|---|---|---|---|
| Disease Index (%) | Control Efficiency (%) e | ||
| 2-(Methylsulfonyl)-5-(4-fluorophenyl)-1,3,4-oxadiazole a | 150 | 2.03 ± 1.22 | 81.34 ± 2.76 A |
| Bismerthiazole b | 375 | 2.59 ± 1.54 | 76.19 ± 3.54 C |
| Zhongshengmycin c | 45 | 3.06 ± 1.86 | 71.87 ± 4.33 B |
| Untreated blank control | 0 | 10.88 ± 2.32 | 0 |
2.4. Determination of SOD and POD Activities

2.5. Determination of Chlorophyll Content in Rice

2.6. Determination of MDA Content in Rice

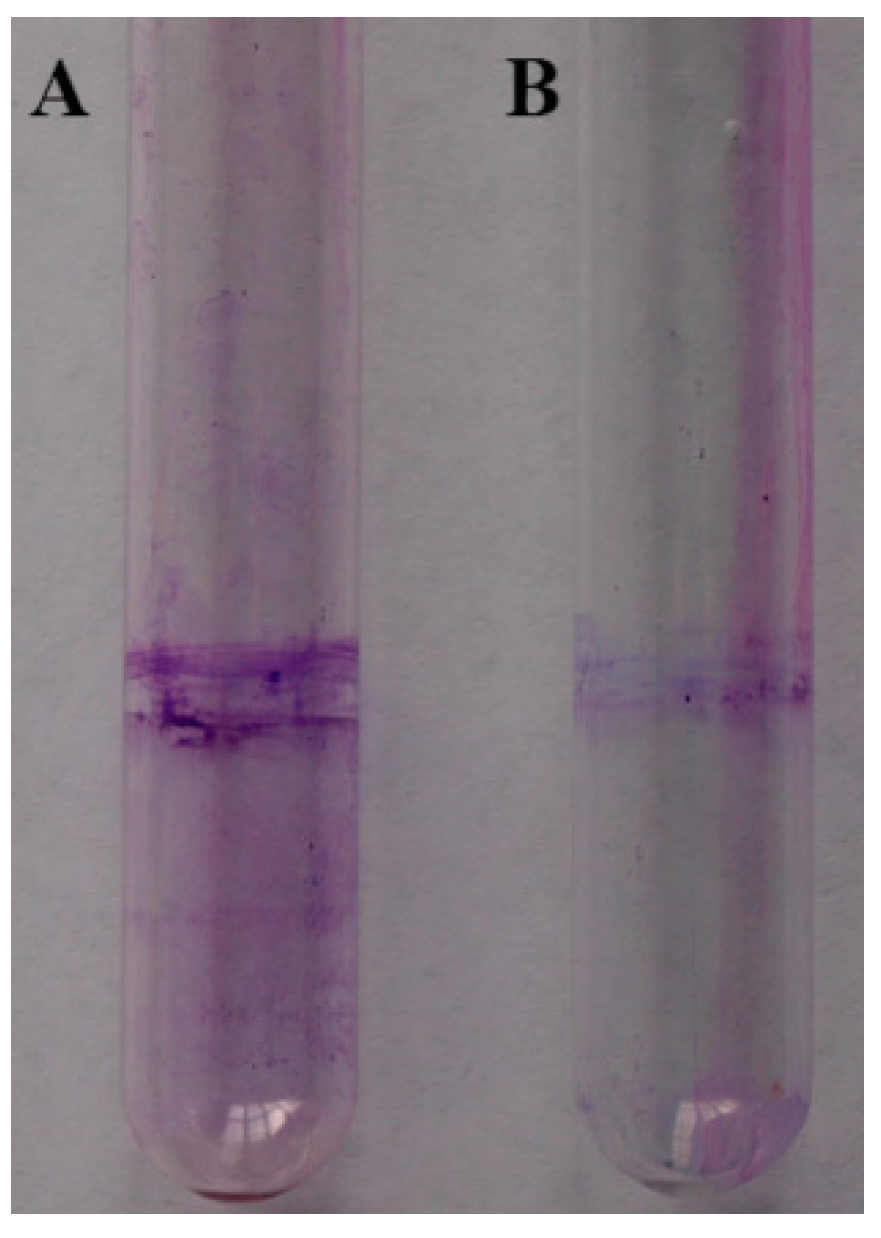
2.7. Biofilm Formation

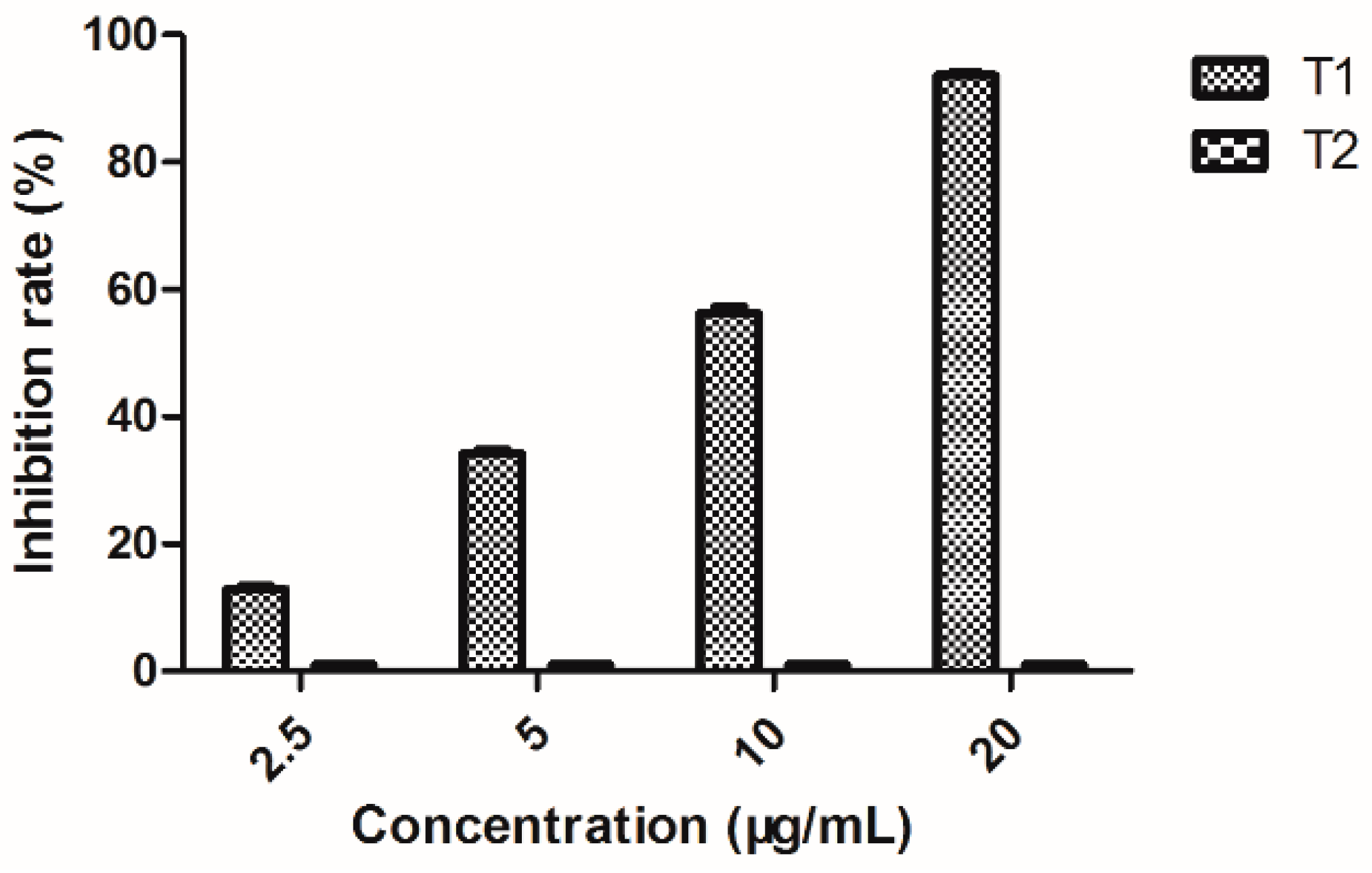
2.8. Quantitative Determination of EPS Production

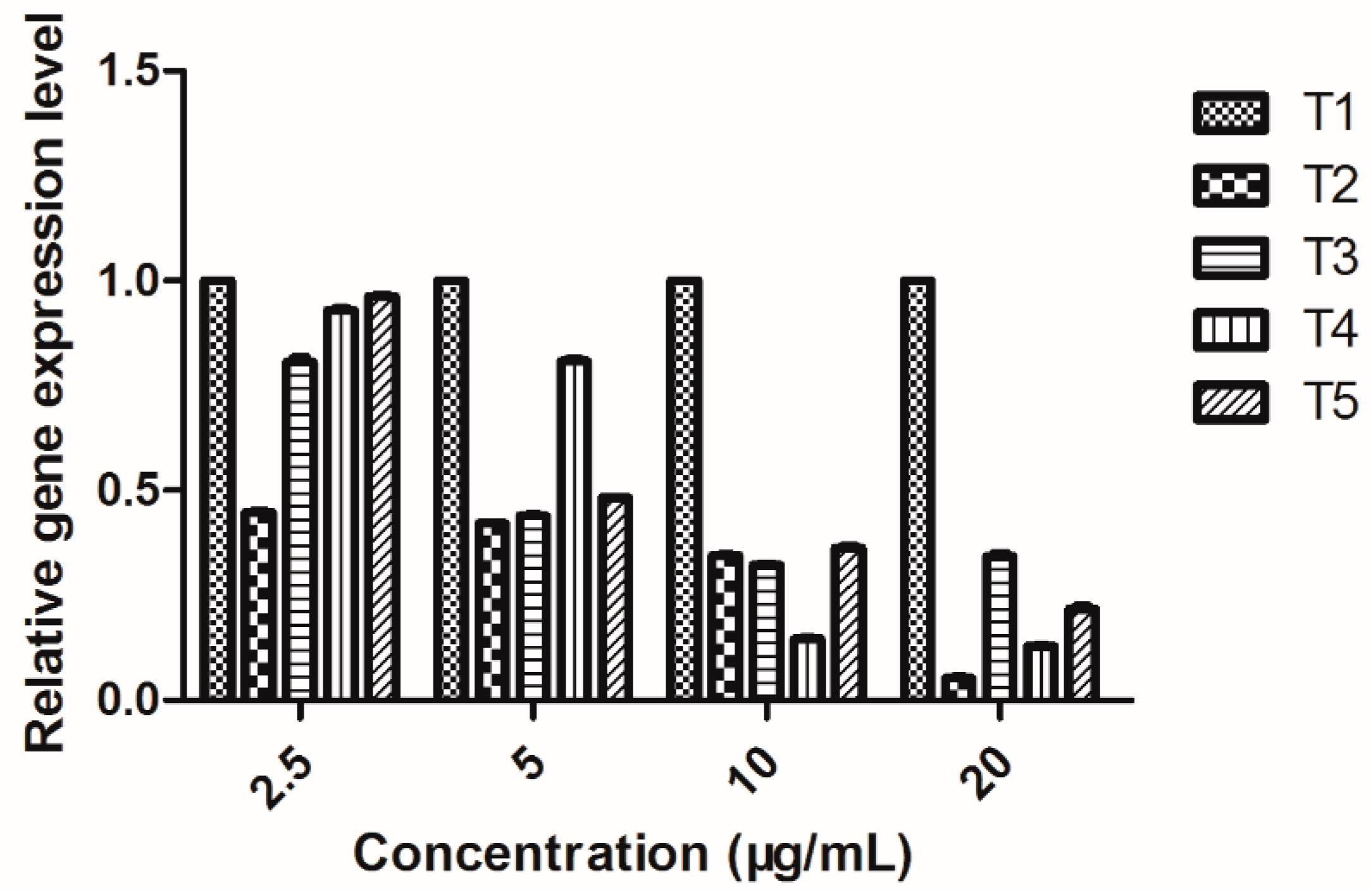
2.9. EPS Gene Expression Level in Xoo

| Genes | Inhibition Rate (%) |
|---|---|
| gumB | 94.88 ± 0.39 |
| gumG | 68.14 ± 1.15 |
| gumM | 86.76 ± 0.28 |
| xanA | 79.21 ± 1.14 |
3. Experimental Section
3.1. Bacterial Strains and Culture Conditions
3.2. In Vitro Antibacterial Activity
3.3. In Vivo Antibacterial Activity
3.4. Field Trial against Rice Bacterial Leaf Blight
3.5. Determination of SOD Activity
3.6. Determination of POD Activity
3.7. Determination of Chlorophyll Content
3.8. Determination of MDA Content
3.9. Biofilm Assays
3.10. Quantitative Determination of EPS Production
3.11. RNA Extraction, cDNA Synthesis, and RT-qPCR Analysis
| Gene Name | GenBank Acc. No. | Primers (5′–3′) |
|---|---|---|
| gumB [32] | 3265197 | Forward: CTGACCGAAATCGAGAAGGGCACCAATC |
| Reverse: GCGCCACACCATCACAAGAGGAGTCAGTTC | ||
| gumG | AF147035 | Forward: GTCACAATGCTTGCTTACA |
| Reverse: ATGGCGATGAAGAACAAC | ||
| gumM | AF231924 | Forward: GTTCTTCGCCAATACCAAT |
| Reverse: TCTCACGACACAGATACG | ||
| xanA | 999354 | Forward: GCAGCGGCGAGATCAACT |
| Reverse: AAACGCCATTCGCCAAAA |
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Mew, T.W. Current status and future prospects of research on bacterial blight of rice. Annu. Rev. Phytopathol. 1987, 25, 359–382. [Google Scholar] [CrossRef]
- Nayak, D.; Shanti, M.L.; Bose, L.K.; Singh, U.D.; Nayak, P. Pathogenicity association in Xanthomonas oryzae pv. oryzae the causal organism of rice bacterial blight disease. ARPN J. Agric. Biol. Sci. 2008, 3, 12–26. [Google Scholar]
- Shanti, M.L.; Devi, G.L.; Kumar, G.N.; Shashidhar, H.E. Molecular marker-assisted selection: A tool for insulating parental lines of hybrid rice against bacterial leaf blight. Int. J. Plant Pathol. 2010, 1, 114–123. [Google Scholar] [CrossRef]
- Evidente, A.; Venturi, V.; Masi, M.; Degrassi, G.; Cimmino, A.; Maddau, L.; Andolfi, A. In vitro antibacterial activity of sphaeropsidins and chemical derivatives toward Xanthomonas oryzae pv. oryzae, the causal agent of rice bacterial blight. J. Nat. Prod. 2011, 74, 2520–2525. [Google Scholar] [CrossRef] [PubMed]
- Khoshkdaman, M.; Ebadi, A.A.; Kahrizi, D. Evaluation of pathogencity and race classification of Xanthomonas oryzae pv. oryzae in guilan province-Iran. Agric. Sci. 2012, 3, 557–561. [Google Scholar]
- Zhu, X.F.; Xu, Y.; Peng, D.; Zhang, Y.; Huang, T.T.; Wang, J.X.; Zhou, M.G. Detection and characterization of bismerthiazol-resistance of Xanthomonas oryzae pv. oryzae. Crop Prot. 2013, 47, 24–29. [Google Scholar] [CrossRef]
- Xu, Y.; Zhu, X.F.; Zhou, M.G.; Kuang, J.; Zhang, Y.; Shang, Y.; Wang, J.X. Status of streptomycin resistance development in Xanthomonas oryzae pv. oryzae and Xanthomonas oryzae pv. oryzicola in China and their resistance characters. J. Phytopathol. 2010, 158, 601–608. [Google Scholar]
- Li, P.; Yin, J.; Xu, W.M.; Wu, J.; He, M.; Hu, D.Y.; Song, B.A. Synthesis, antibacterial activities, and 3D-QSAR of sulfone derivatives containing 1,3,4-oxadiazole moiety. Chem. Biol. Drug Des. 2013, 82, 546–556. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Shi, L.; Yang, X.; Yang, L.; Chen, X.W.; Wu, F.; Song, B.A. Design, synthesis, and antibacterial activity against rice bacterial leaf blight and leaf streak of 2,5-substituted-1,3,4-oxadiazole/thiadiazole sulfone derivative. Bioorg. Med. Chem. Lett. 2014, 24, 1677–1680. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.M.; Han, F.F.; He, M.; Hu, D.Y.; He, J.; Yang, S.; Song, B.A. Inhibition of tobacco bacterial wilt with sulfone derivatives containing 1,3,4-oxadiazole moiety. J. Agric. Food Chem. 2012, 60, 1036–1041. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.M.; He, J.; He, M.; Han, F.F.; Chen, X.H.; Pan, Z.X.; Wang, J.; Tong, M.G. Synthesis and antifungal activity of novel sulfone derivatives containing 1,3,4-oxadiazole moieties. Molecules 2011, 16, 9129–9141. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.M.; Yang, S.; Bhadury, P.; He, J.; He, M.; Gao, L.L.; Song, B.A. Synthesis and bioactivity of novel sulfone derivatives containing 2,4-dichlorophenyl substituted 1,3,4-oxadiazole/thiadiazole moiety as chitinase inhibitors. Pestic. Biochem. Physiol. 2011, 101, 6–15. [Google Scholar] [CrossRef]
- Scarf, M.E.; Siegfried, B.D.; Meinke, L.J.; Chandler, L.D. Fipronil metabolism, oxidative sulfone formation and toxicity among organophosphate- and carbamate- resistant and susceptible western corn rootworm populations. Pest Manag. Sci. 2000, 56, 757–766. [Google Scholar] [CrossRef]
- Oliveira, C.S.; Lira, B.F.; Barbosa-Filho, J.M.; Fernandez Lorenzo, J.G.; Athayde-Filho, P.F. Synthetic approaches and pharmacological activity of 1,3,4-oxadiazoles: A review of the literature from 2000–2012. Molecules 2012, 17, 10192–10231. [Google Scholar] [CrossRef] [PubMed]
- Vedula, M.S.; Pulipaka, A.B.; Venna, C.V.; Chintakunta, K.; Jinnapally, S.; Kattuboina, V.A. New styryl sulfones as anticancer agents. Eur. J. Med. Chem. 2003, 38, 811–824. [Google Scholar] [CrossRef]
- Silvestri, R.; Artico, M.; la Regina, G.; de Martino, G.; la Colla, M.; Loddo, R.; la Colla, P. Anti-HIV-1 activity of pyrryl aryl sulfone (PAS) derivatives: Synthesis and SAR studies of novel esters and amides at the position 2 of the pyrrole nucleus. Farmaco 2004, 59, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Hermes-Lima, M. Oxygen in biology and biochemistry: Role of free radicals. In Functional Metabolism: Regulation and Adaptation; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2004; Volume 1, pp. 319–366. [Google Scholar]
- Moura, J.C.M.S.; Bonine, C.A.V.; de Oliveira Fernandes Viana, J.; Dornelas, M.C.; Mazzafera, P. Abiotic and biotic stresses and changes in the lignin content and composition in plants. J. Integr. Plant Biol. 2010, 52, 360–376. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.T.; Song, B.A.; Bhadury, P.S.; Jin, L.H.; Hu, D.Y.; Yang, S. Antiviral activity and mechanism of action of novel thiourea containing chiral phosphonate on tobacco mosaic virus. Int. J. Mol. Sci. 2011, 12, 4522–4535. [Google Scholar] [CrossRef] [PubMed]
- Ghasemie, E.; Kazempour, M.; Padasht, F. Isolation and identification of Xathomonas oryzae pv. oryzae the causal agent of bacterial blight of rice in Iran. Eur. J. Plant Pathol. 2008, 48, 53–62. [Google Scholar] [CrossRef]
- Janero, D.R. Malondialdehyde and thiobarbituric acid-reactivity as diagnostic indices of lipid peroxidation and peroxidative tissue injury. Free Radic. Biol. Med. 1990, 9, 515–540. [Google Scholar] [CrossRef]
- Ielpi, L.; Couso, R.O.; Dankert, M.A. Sequential assembly and polymerization of the polyprenol-linked pentasaccharide repeating unit of the xanthan polysaccharide in Xanthomonas campestris. J. Bacteriol. 1993, 175, 2490–2500. [Google Scholar] [PubMed]
- Hung, C.H.; Wu, H.C.; Tseng, Y.H. Mutation in the Xanthomonas campestrisxan A gene required for synthesis of xanthan and lipopolysaccharide drastically reduces the efficiency of bacteriophage φL7 adsorption. Biochem. Biophys. Res. Commun. 2002, 291, 338–343. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.M.; Park, Y.J.; Park, D.S.; Kang, H.W.; Kim, J.G.; Song, E.S.; Go, S.J. The genome sequence of Xanthomonas oryzae pathovar oryzae KACC10331, the bacterial blight pathogen of rice. Nucleic Acids Res. 2005, 33, 577–586. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Kim, J.G.; Lee, B.M.; Cho, J.Y. Mutational analysis of the gum gene cluster required for xanthan biosynthesis in Xanthomonas oryzae pv oryzae. Biotechnol. Lett. 2009, 31, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Dalgaard, P.; Ross, T.; Kamperman, L.; Neumeyer, K.; McMeekin, T.A. Estimation of bacterial growth rates from turbidimetric and viable count data. Int. J. Food Microbiol. 1994, 23, 91–404. [Google Scholar] [CrossRef]
- Schaad, N.W.; Wang, Z.K.; Di, M.; McBeath, J.; Peterson, G.L.; Bonde, M.R. An improved infiltration technique to test the pathogenicity of Xanthomonas oryzae pv. oryzae in rice seedlings. Seed Sci. Technol. 1996, 24, 449–456. [Google Scholar]
- Constantine, N.G.; Stanley, K.R. Superoxide dismutases. Plant Physiol. 1977, 59, 309–314. [Google Scholar]
- Rathmell, W.G.; Sequeira, L. Soluble peroxidase in fluid from the intercellular spaces of tobacco leaves. Plant Physiol. 1974, 53, 317–318. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.S.; Liu, E. Studies of method on extract chlorophyll a and b. Acta Agric. Univ. Pekin. 1992, 18, 247–250. [Google Scholar]
- Draper, H.H.; Hadley, M. Malondialdehyde determination as index of lipid Peroxidation. Methods Enzymol. 1990, 186, 421–431. [Google Scholar] [PubMed]
- Pratt, L.A.; Kolter, R. Genetic analysis of Escherichia coli biofilm formation: Roles of flagella, motility, chemotaxis and type I pili. Mol. Microbiol. 1998, 30, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhang, K.; Zhou, X.; Xu, N.; Xu, H.H.K.; Weir, M.D.; Ge, Y.; Wang, S.; Li, M.; Li, Y.; et al. Antibacterial Effect of Dental Adhesive Containing Dimethylaminododecyl Methacrylate on the Development of Streptococcus mutans Biofilm. Int. J. Mol. Sci. 2014, 12791, 12791–12806. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Sagaram, U.S.; Kim, J.S.; Wang, N. Requirement of the galU gene for polysaccharide production by and pathogenicity and growth in planta of Xanthomonascitri subsp. Appl. Environ. Microbiol. 2010, 76, 2234–2242. [Google Scholar] [CrossRef] [PubMed]
- Vojnov, A.A.; Zorreguieta, A.; Dow, J.M.; Daniels, M.J.; Dankert, M.A. Evidence for a role for the gumB and gumC gene products in the formation of xanthan from its pentasaccharide repeating unit by Xanthomonas campestris. Microbiology 1998, 144, 1487–1493. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Pan, X.; Luo, J.; Wu, J.; Zhou, Z.; Liang, X.; Zhou, M.G. Effects of phenazine-1-carboxylic acid on the biology of the plant-pathogenic bacterium Xanthomonas oryzae pv. oryzae. Pestic. Biochem. Physiol. 2015, 117, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the compounds are available from the authors.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, L.; Li, P.; Wang, W.; Gao, M.; Wu, Z.; Song, X.; Hu, D. Antibacterial Activity and Mechanism of Action of Sulfone Derivatives Containing 1,3,4-Oxadiazole Moieties on Rice Bacterial Leaf Blight. Molecules 2015, 20, 11660-11675. https://doi.org/10.3390/molecules200711660
Shi L, Li P, Wang W, Gao M, Wu Z, Song X, Hu D. Antibacterial Activity and Mechanism of Action of Sulfone Derivatives Containing 1,3,4-Oxadiazole Moieties on Rice Bacterial Leaf Blight. Molecules. 2015; 20(7):11660-11675. https://doi.org/10.3390/molecules200711660
Chicago/Turabian StyleShi, Li, Pei Li, Wenli Wang, Manni Gao, Zengxue Wu, Xianpeng Song, and Deyu Hu. 2015. "Antibacterial Activity and Mechanism of Action of Sulfone Derivatives Containing 1,3,4-Oxadiazole Moieties on Rice Bacterial Leaf Blight" Molecules 20, no. 7: 11660-11675. https://doi.org/10.3390/molecules200711660
APA StyleShi, L., Li, P., Wang, W., Gao, M., Wu, Z., Song, X., & Hu, D. (2015). Antibacterial Activity and Mechanism of Action of Sulfone Derivatives Containing 1,3,4-Oxadiazole Moieties on Rice Bacterial Leaf Blight. Molecules, 20(7), 11660-11675. https://doi.org/10.3390/molecules200711660