
The Efficacy and Underlying Mechanism of Sulfone Derivatives Containing 1,3,4-oxadiazole on Citrus Canker

Abstract
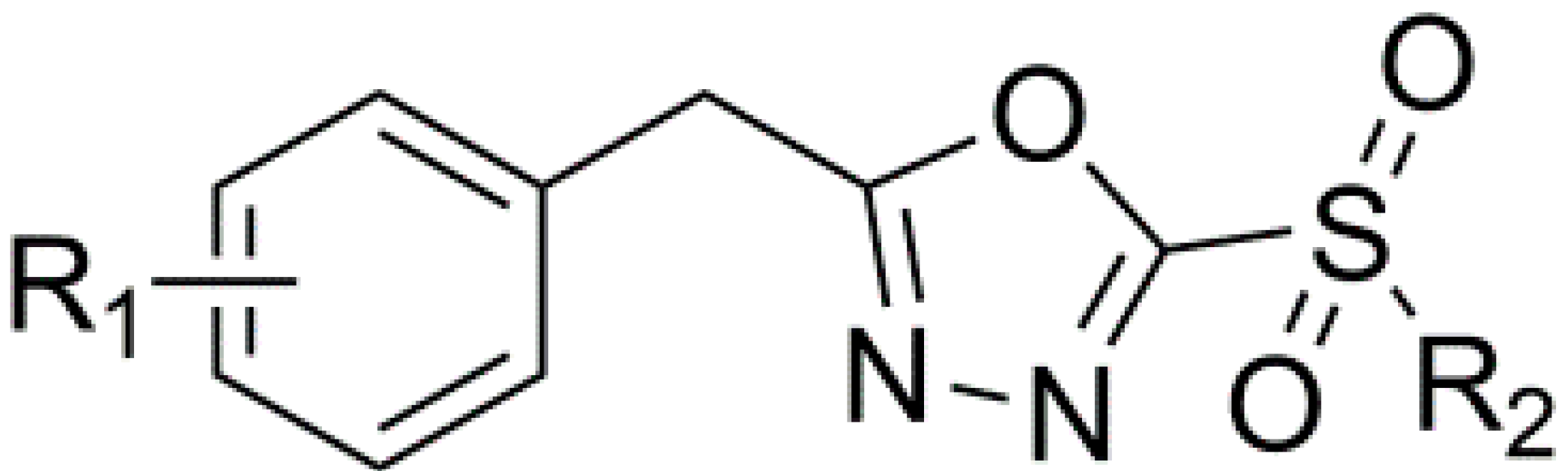
:1. Introduction

2. Results and Discussion
2.1. DNA Extraction, PCR Amplification, Sequencing, and Identification of Species

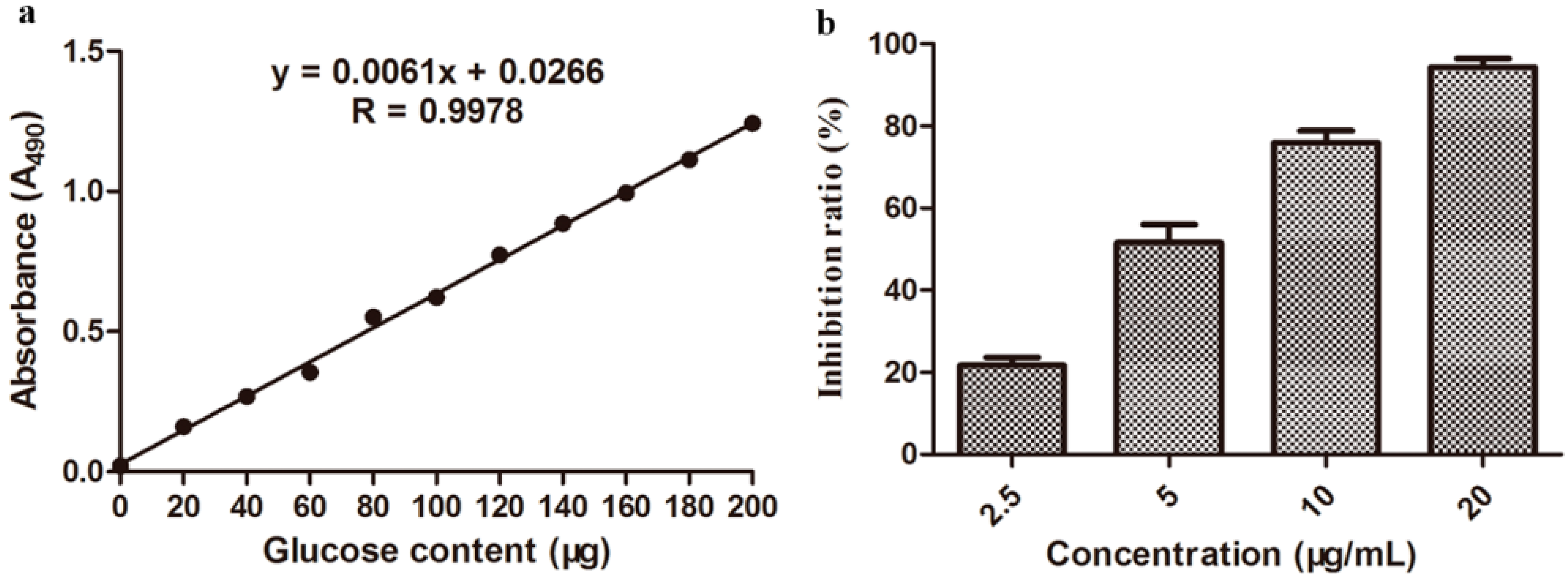
2.2. In Vitro Antibacterial Bioassay

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. |  | Toxic Regression Equation | r | EC50 (μg/mL) | |
|---|---|---|---|---|---|
| R1 | R2 | ||||
| 1 | H | –CH3 | y = 1.42x + 3.84 | 0.99 | 6.52 ± 1.19 |
| 2 | H | –CH2CH3 | y = 1.56x + 2.75 | 0.98 | 27.80 ± 2.76 |
| 3 | 4-F | –CH3 | y = 1.56x + 4.86 | 0.99 | 1.23 ± 0.97 |
| 4 | 4-F | –CH2CH3 | y = 1.42x + 3.28 | 0.99 | 16.16 ± 2.21 |
| 5 | 4-Cl | –CH3 | y = 1.56x + 3.30 | 0.99 | 12.28 ± 1.76 |
| 6 | 4-Cl | –CH2CH3 | y = 1.53x + 2.43 | 0.99 | 48.54 ± 2.78 |
| 7 | 2,4-2Cl | –CH3 | y = 1.58x + 4.38 | 0.96 | 2.47 ± 0.69 |
| 8 | 2,4-2Cl | –CH2CH3 | y = 1.40x + 2.92 | 0.97 | 30.10 ± 3.87 |
| Kocide 3000 | y = 1.61x + 2.15 | 0.98 | 58.21 ± 2.77 | ||
| Thiodiazole copper | y = 2.15x + 0.94 | 0.98 | 77.04 ± 1.96 | ||
2.3. Field Trials against Citrus Canker
| Treatment | Congjiang, Guizhou Province | Luodian, Guizhou Province | ||
|---|---|---|---|---|
| In Leaves a | In Fruits a | In Leaves a | In Fruits a | |
| 3 | 66.31 ± 2.45A | 69.03 ± 5.12A | 60.43 ± 5.67A | 64.51 ± 2.23A |
| Kocide 3000 | 55.13 ± 5.63B | 53.76 ± 4.43B | 50.93 ± 4.34C | 52.77 ± 5.98B |
| Thiodiazole copper | 61.47 ± 2.32C | 57.73 ± 3.73C | 55.72 ± 2.86B | 56.52 ± 3.76C |
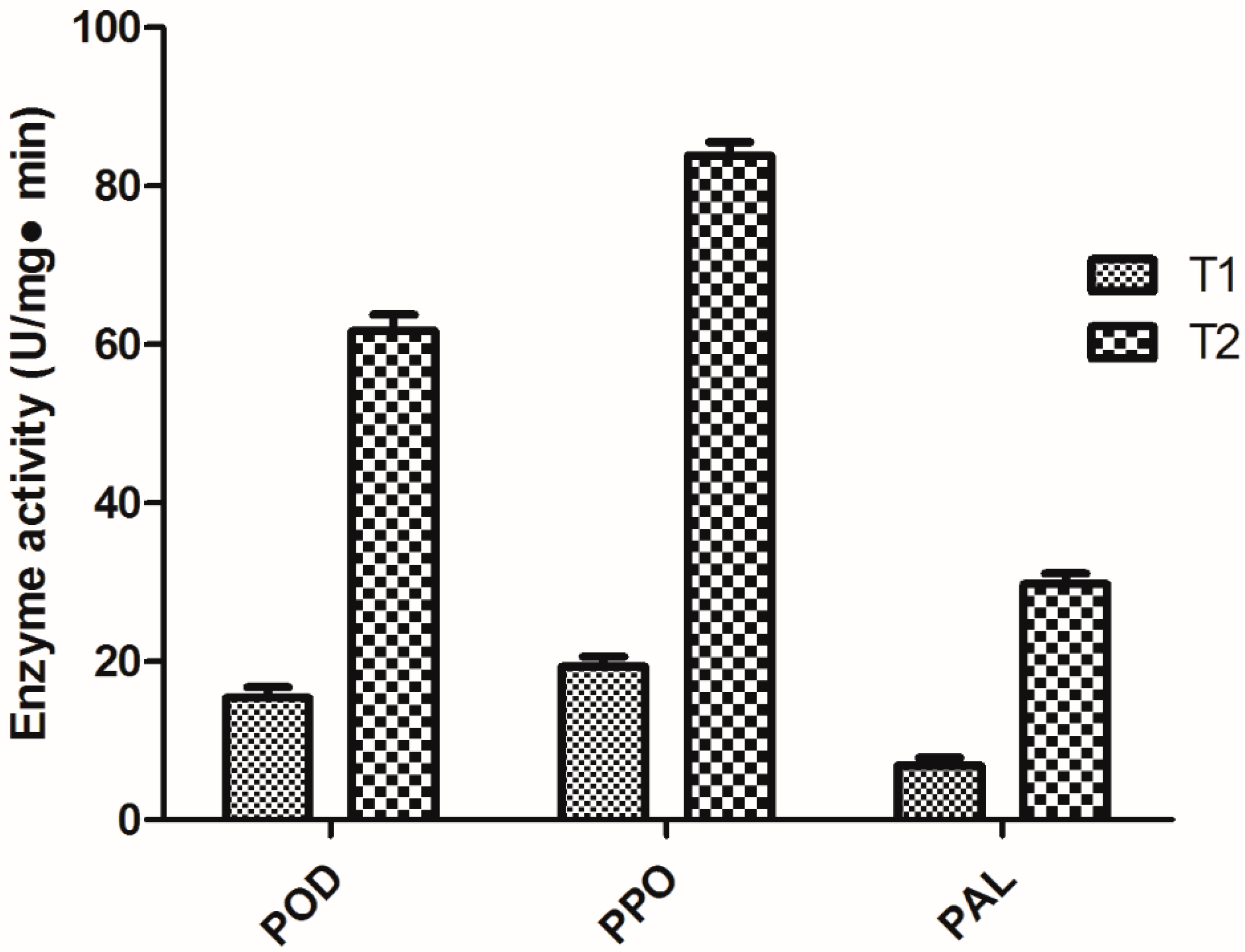
2.4. Determination of Peroxidase (POD), Polyphenol oxidase (PPO), and Phenylalanine ammonia lyase (PAL) Activities

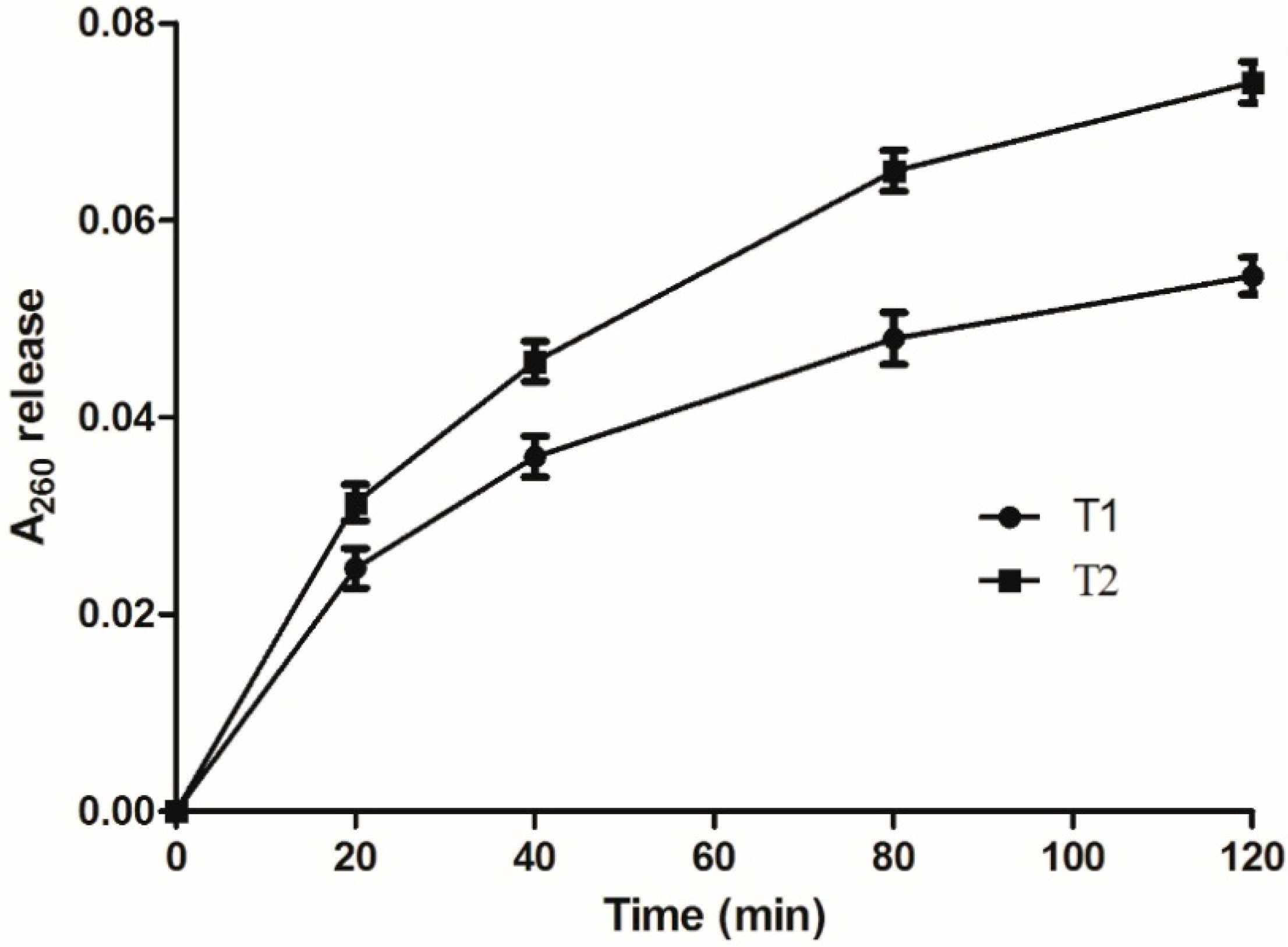
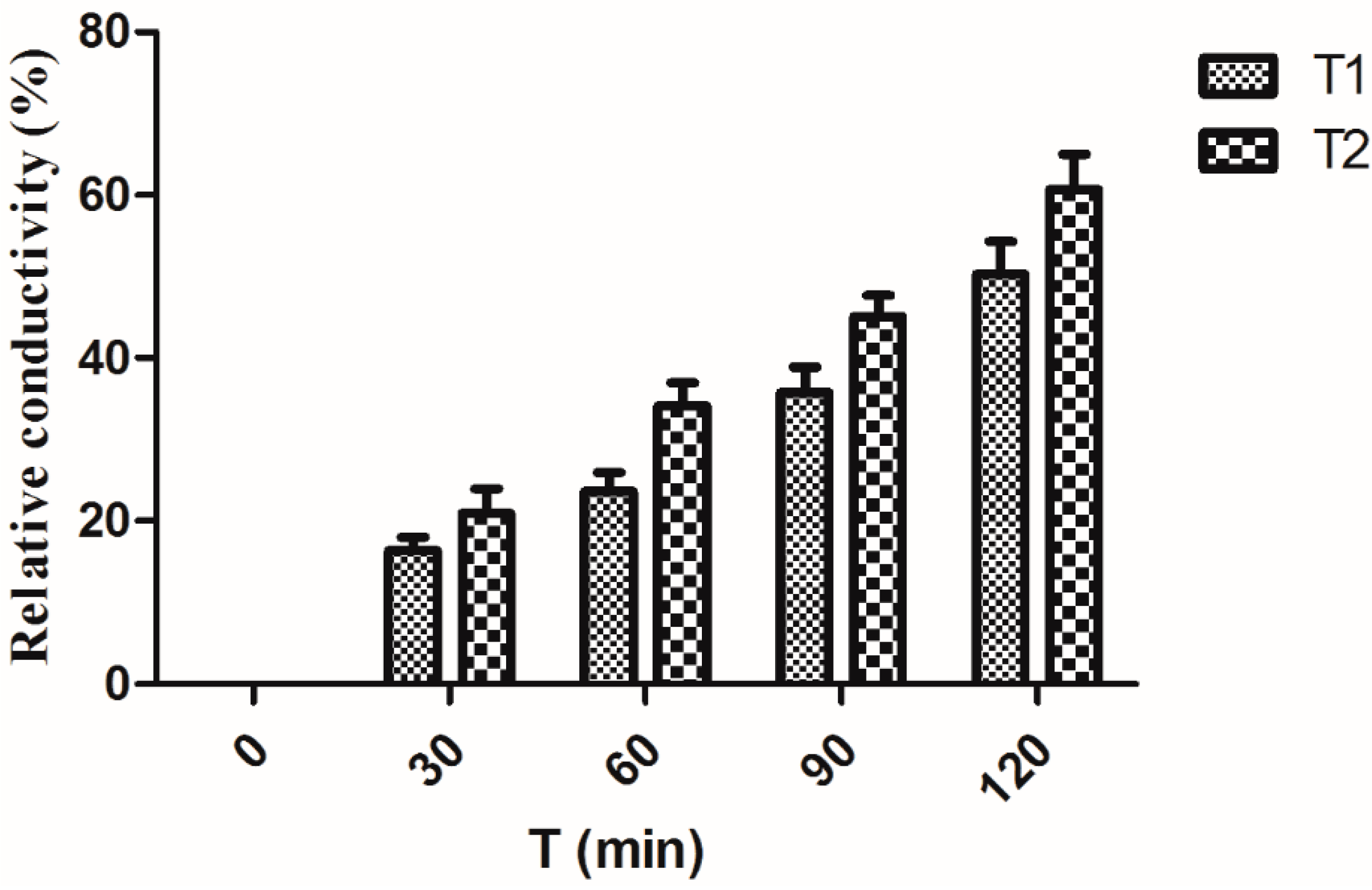
2.5. Effect on the Integrity of Bacterial Cell Membranes

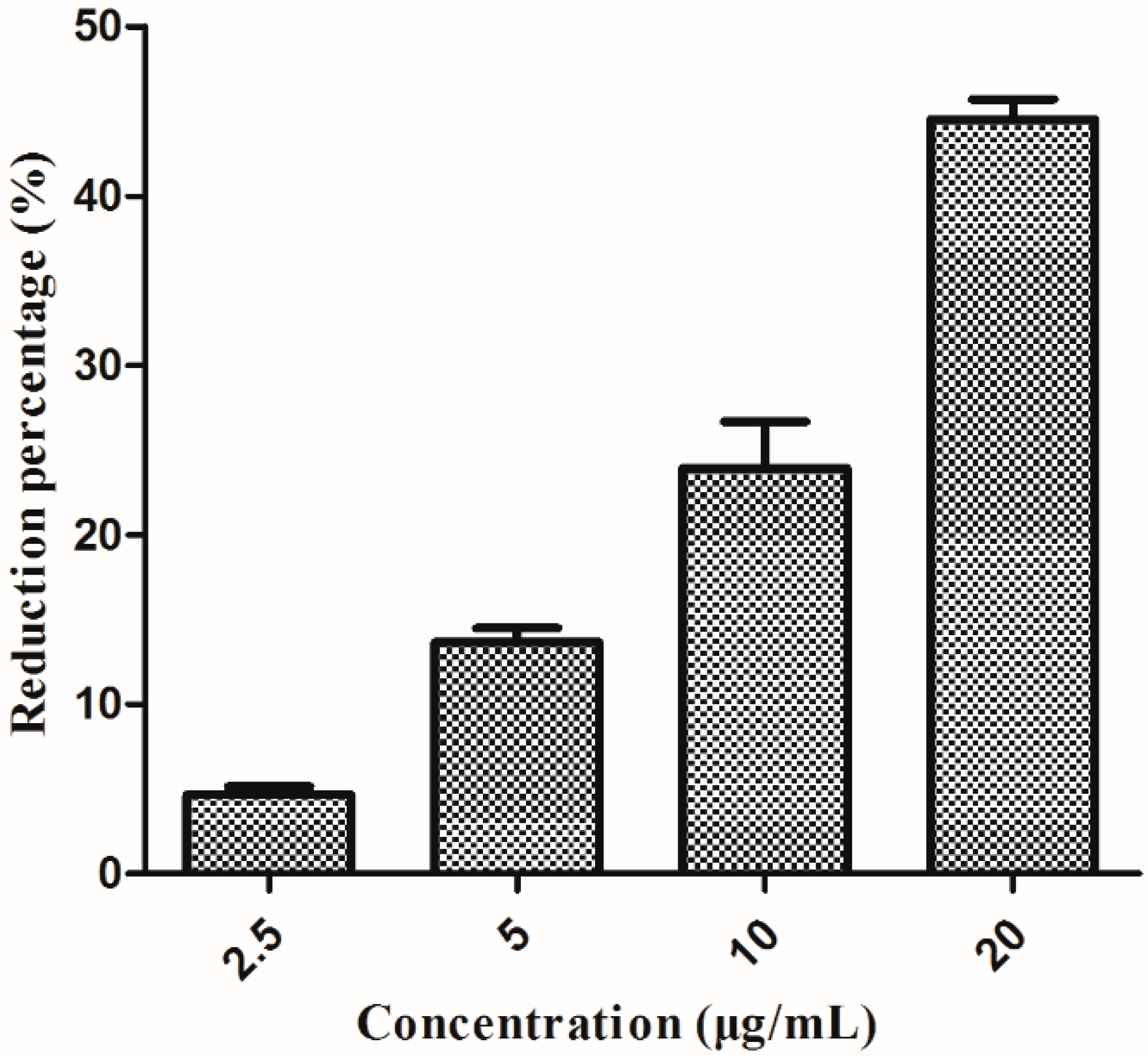
2.6. Effect on the Biofilm Formation

2.7. Effect on Cell Membrane Permeability of Xcc

2.8. Determination of Exopolysaccharide (EPS) Content

3. Experimental Section
3.1. Bacteria Isolation and Purification
3.2. DNA Extraction, PCR Amplification, and Sequencing of Species
3.3. In Vitro Antibacterial Bioassay
3.4. Field Trial against Citrus Canker
3.5. Determination of POD, PPO, and PAL Activities
3.6. Effect on the Cell Membrane Integrity
3.7. Effect on the Biofilm Formation
3.8. Effect on the Cell Membrane Permeability
3.9. EPS Content
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Das, A.K. Citrus canker-a review. J. Appl. Hortic. 2003, 5, 52–60. [Google Scholar]
- Gottwald, T.R.; Graham, J.H.; Schubert, T.S. Citrus canker: The pathogen and its impact. Plant Health Prog. 2002. [Google Scholar] [CrossRef]
- Gottwald, T.R.; Hughes, G.; Graham, J.H.; Sun, X.; Riley, T. The citrus canker epidemic in Florida: The scientific basis of regulatory eradication policy for an invasive species. Phytopathology 2001, 91, 30–34. [Google Scholar] [CrossRef] [PubMed]
- Graham, J.H.; Gottwald, T.R.; Cubero, J.; Achor, D.S. Xanthomonas axonopodis pv. citri: Factors affecting successful eradication of citrus canker. Mol. Plant Pathol. 2004, 5, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Cubero, J.; Graham, J.H. Genetic relationship among worldwide strains of Xanthomonas causing canker in citrus species and design of new primers for their identification by PCR. Appl. Environ. Microbiol. 2002, 68, 1257–1264. [Google Scholar] [CrossRef] [PubMed]
- Schaad, N.; Postnikova, E.; Lacy, G.; Sechler, A.; Agarkova, I.; Stromberg, P.; Stromberg, V.; Vidaver, A. Emended classification of Xanthomonad pathogens on citrus. Syst. Appl. Microbiol. 2006, 29, 690–695. [Google Scholar] [CrossRef] [PubMed]
- Vauterin, L.; Hoste, B.; Kersters, K.; Swings, J. Reclassification of Xanthomonas. Int. J. Syst. Bacteriol. 1995, 45, 472–489. [Google Scholar] [CrossRef]
- Gottwald, T.R.; Graham, J.H. A device for precise and nondisruptive stomatal inoculation of leaf tissue with bacterial pathogens. Phytopathology 1992, 82, 930–935. [Google Scholar] [CrossRef]
- Pruvost, O.; Boher, B.; Brocherieux, C.; Nicole, M.; Chiroleu, F. Survival of Xanthomonas axonopodis pv. citri in leaf lesions under tropical environmental conditions and simulated splash dispersal of inoculum. Phytopathology 2002, 92, 336–346. [Google Scholar] [CrossRef] [PubMed]
- Bock, C.H.; Parker, P.E.; Gottwald, T.R. The effect of simulated wind-driven rain on duration and distance of dispersal of Xanthomonas axonopodis pv. citri from canker infected citrus trees. Plant Dis. 2005, 89, 71–80. [Google Scholar] [CrossRef]
- Bock, C.H.; Graham, J.; Gottwald, T.R.; Cook, A.Z.; Parker, P.E. Wind speed effects on the quantity of Xanthomonas citri subsp. citri dispersed downwind from canopies of grapefruit trees infected with citrus canker. Plant Dis. 2010, 94, 725–736. [Google Scholar] [CrossRef]
- Gottwald, T.R.; Irey, M. Post-hurricane analysis of citrus canker II: Predictive model estimation of disease spread and area potentially impacted by various eradication protocols following catastrophic weather events. Plant Health Prog. 2007. [Google Scholar] [CrossRef]
- Li, J.Y.; Wang, N. Foliar application of biofilm formation-inhibiting compounds enhances control of citrus canker caused by Xanthomonas citri subsp. citri. Phytopathology 2014, 104, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Shi, L.; Yang, X.; Chen, X.W.; Wu, F.; Shi, Q.C.; Xu, W.M.; He, M.; Hu, D.Y.; Song, B.A. Design, synthesis, and antibacterial activity against rice bacterial leaf blight and leaf streak of 2,5-substituted-1,3,4-oxadiazole/thiadiazole sulfone derivative. Bioorg. Med. Chem. Lett. 2014, 24, 1677–1680. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.P.; Liu, X.Y.; Diao, Y.; Ding, Z.L.; Hu, Z.L. Authentication and characterization of a candidate antagonistic bacterium against soft rot of Amorphophallus konjac. Crop Prot. 2012, 34, 83–87. [Google Scholar] [CrossRef]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [PubMed]
- Paw, D.; Thomas, R.; Laura, K.; Karina, N.; Thomas, A.M. Estimation of bacterial growth rates from turbidimetric and viable count data. Int. J. Food. Microbiol. 1994, 23, 391–404. [Google Scholar]
- Vanitha, S.C.; Niranjana, S.R.; Umesha, S. Role of phenylalanine ammonia lyase and polyphenol oxidase in host resistance to bacterial wilt of tomato. J. Phytopathol. 2009, 157, 552–557. [Google Scholar] [CrossRef]
- Schurter, R.; Kunz, W.; Nyfelder, R. Benzothiadiazole und Ihre Verwendung in Verfahren und Mitteln Gegen Pflanzenkrankheiten. EU Patent 0313512, 25 November 1992. [Google Scholar]
- Schurter, R.; Kunz, W.; Nyfelder, R. Process and a Composition for Immunizing Plants against Diseases. U.S. Patent 4931581, 5 June 1990. [Google Scholar]
- Yoshida, H.; Konishi, K.; Koike, K.; Nakagawa, T.; Sekido, S.; Yamaguchi, I. Effect of N-Cyanomethyl-2-chloroisonicotinamide for control of rice blast. J. Pestic. Sci. 1990, 15, 413–417. [Google Scholar] [CrossRef]
- Yoshioka, K.; Nakashita, H.; Klessig, D.F.; Yamaguchi, I. Probenazole induces systemic acquired resistance in Arabidopsis with a novel type of action. Plant J. 2001, 25, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Durrant, W.E.; Dong, X. Systemic acquired resistance. Annu. Rev. Phytopathol. 2004, 42, 185–209. [Google Scholar] [CrossRef] [PubMed]
- Fukasawa-Akada, T.; Kung, S.; Watson, J.A. Phenylalanine ammonia lyase gene structure, expression, and evolution in Nicotiana. Plant Mol. Biol. 1996, 30, 711–722. [Google Scholar] [CrossRef] [PubMed]
- Hammerschmidt, R.; Nuckles, E.M.; Kuc, J. Association of enhanced peroxidase activity with induced systemic resistance of cucumber to Colletotrichum lagenarium. Physiol. Plant Pathol. 1982, 20, 73–83. [Google Scholar] [CrossRef]
- Mauch, F.; Mauch-Mani, B.; Boller, T. Antifungal hydrolases in pea tissue: II. Inhibition of fungal growth by combinations of chitinase and β-1,3-glucanase. Plant Physiol. 1988, 88, 936–942. [Google Scholar] [CrossRef] [PubMed]
- Hemm, M.R.; Rider, S.D.; Ogas, J.; Murry, D.J.; Chapple, C. Light induces phenylpropanoid metabolism in Arabidopsis roots. Plant J. 2004, 38, 765–778. [Google Scholar] [CrossRef] [PubMed]
- Moura, J.C.M.S.; Bonine, C.A.V.; de Oliveira Fernandes Viana, J.; Dornelas, M.C.; Mazzafera, P. Abiotic and biotic stresses and changes in the lignin content and composition in plants. J. Integr. Plant Biol. 2010, 52, 360–376. [Google Scholar] [CrossRef] [PubMed]
- Luh, B.S.; Phithakpol, B. Characteristics of polyphenol oxidase related to browning in cling peaches. J. Food Sci. 1972, 37, 264–268. [Google Scholar] [CrossRef]
- Jiang, A.L.; Tian, S.P.; Xu, Y. Effect of controlled atmospheres with high-O2 or high-CO2 concentrations on postharvest physiology and storability of “Napoleon” sweet cherry. J. Integr. Plant Biol. 1984, 44, 925–930. [Google Scholar]
- Matuschek, E.; Svanberg, U. The effect of fruit extracts with polyphenol peroxidase (PPO) activity on the in vitro accessibility of iron in high-tannin sorghum. Food Chem. 2005, 90, 765–771. [Google Scholar] [CrossRef]
- Assis, J.S.; Maldonado, R.; Muñoz, T.; Escribano, M.I.; Merodio, C. Effect of high carbon dioxide concentration on PAL activity and phenolic contents in ripening cherimoya fruit. Postharvest Biol. Technol. 2001, 23, 33–39. [Google Scholar] [CrossRef]
- Chen, C.Z.; Cooper, S.L. Interactions between dendrimer biocides and bacterial membranes. Biomaterials 2002, 23, 3359–3368. [Google Scholar] [CrossRef]
- Li, Y.H.; Tan, N.; Aspiras, M.B.; Lau, P.C.Y.; Lee, J.H.; Ellen, R.P.; Cvitkovitch, D.G. A quorum-sensing signaling system essential for genetic competence in Streptococcus mutans is involved in biofilm formation. J. Bacteriol. 2002, 184, 2699–2708. [Google Scholar] [CrossRef] [PubMed]
- Loo, C.Y.; Corliss, D.A.; Ganeshkumar, N. Streptococcus gordonii biofilm formation: Identification of genes that code for biofilm phenotypes. J. Bacteriol. 2000, 182, 1374–1382. [Google Scholar] [CrossRef] [PubMed]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Robers, P.A.; Smith, F. Calorimetric method for determinination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Rao, P.; Pattabiraman, T.N. Reevaluation of the phenol-sulfuric acid reaction for the estimation of hexoses and pentoses. Anal. Biochem. 1989, 181, 18–22. [Google Scholar] [CrossRef]
- Duan, Y.B.; Ge, C.Y.; Liu, S.M.; Chen, C.J.; Zhou, M.G. Effect of phenylpyrrole fungicide fludioxonil on morphological and physiological characteristics of Sclerotinia sclerotiorum. Pestic. Biochem. Phys. 2013, 106, 61–67. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the compounds are available from the authors.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, P.; Ma, Y.; Zhou, J.; Luo, H.; Yan, J.; Mao, Y.; Wang, Z. The Efficacy and Underlying Mechanism of Sulfone Derivatives Containing 1,3,4-oxadiazole on Citrus Canker. Molecules 2015, 20, 14103-14117. https://doi.org/10.3390/molecules200814103
Li P, Ma Y, Zhou J, Luo H, Yan J, Mao Y, Wang Z. The Efficacy and Underlying Mechanism of Sulfone Derivatives Containing 1,3,4-oxadiazole on Citrus Canker. Molecules. 2015; 20(8):14103-14117. https://doi.org/10.3390/molecules200814103
Chicago/Turabian StyleLi, Pei, Yuhua Ma, Junliang Zhou, Hui Luo, Jiawen Yan, Yongya Mao, and Zhuang Wang. 2015. "The Efficacy and Underlying Mechanism of Sulfone Derivatives Containing 1,3,4-oxadiazole on Citrus Canker" Molecules 20, no. 8: 14103-14117. https://doi.org/10.3390/molecules200814103
APA StyleLi, P., Ma, Y., Zhou, J., Luo, H., Yan, J., Mao, Y., & Wang, Z. (2015). The Efficacy and Underlying Mechanism of Sulfone Derivatives Containing 1,3,4-oxadiazole on Citrus Canker. Molecules, 20(8), 14103-14117. https://doi.org/10.3390/molecules200814103