
Macrophage Activation by Ursolic and Oleanolic Acids during Mycobacterial Infection


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction

2. Results and Discussion
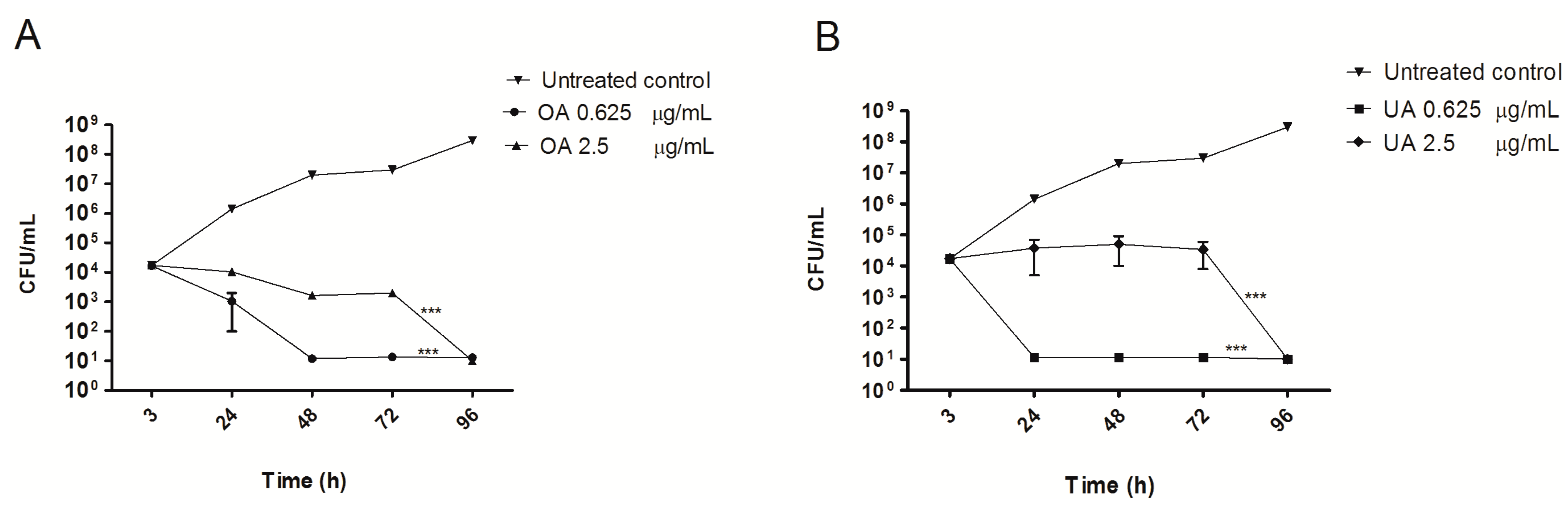
2.1. Cytotoxicity and Intracellular Survival of Mycobacteria in J774 A.1 Cells Treated with OA and UA

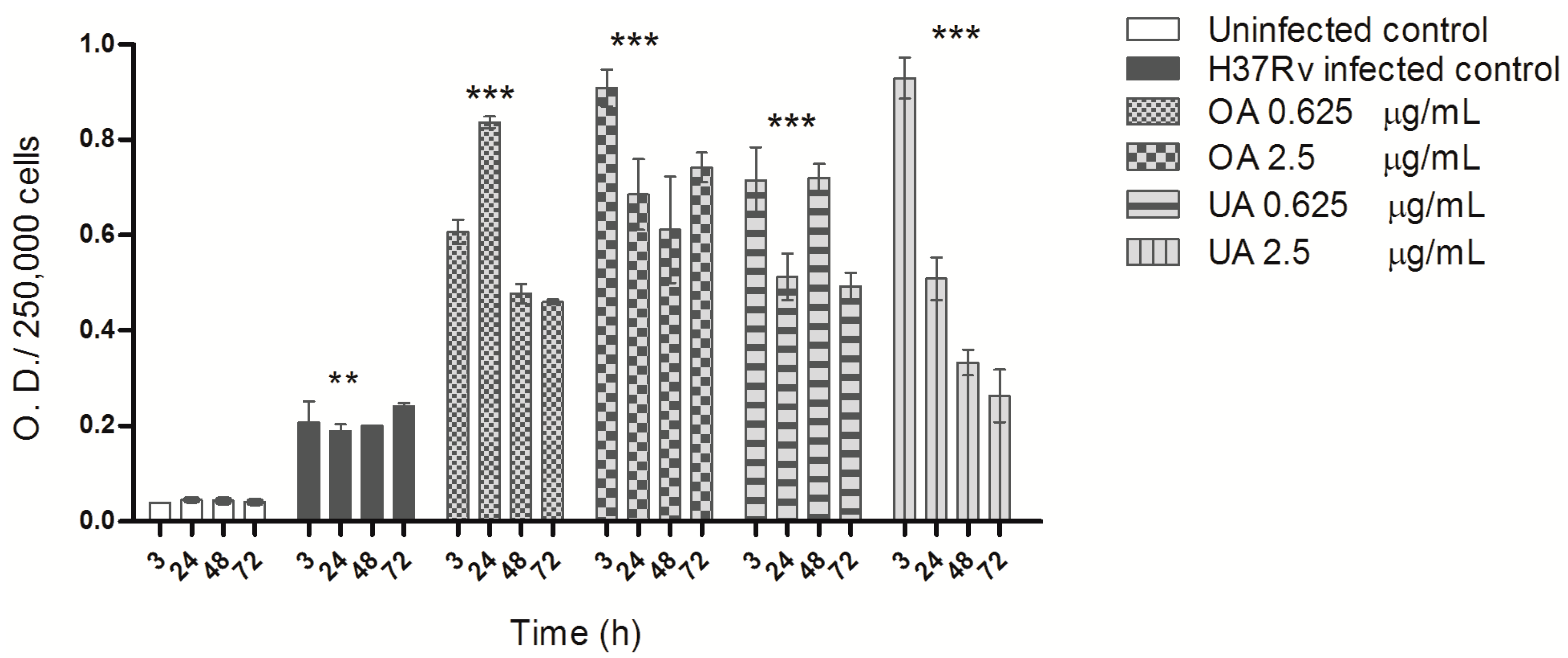
2.2. ROS Production during Mycobacterial Infection and Triterpene Treatment

2.3. NO Production during Mycobacterial Infection and Triterpene Treatment

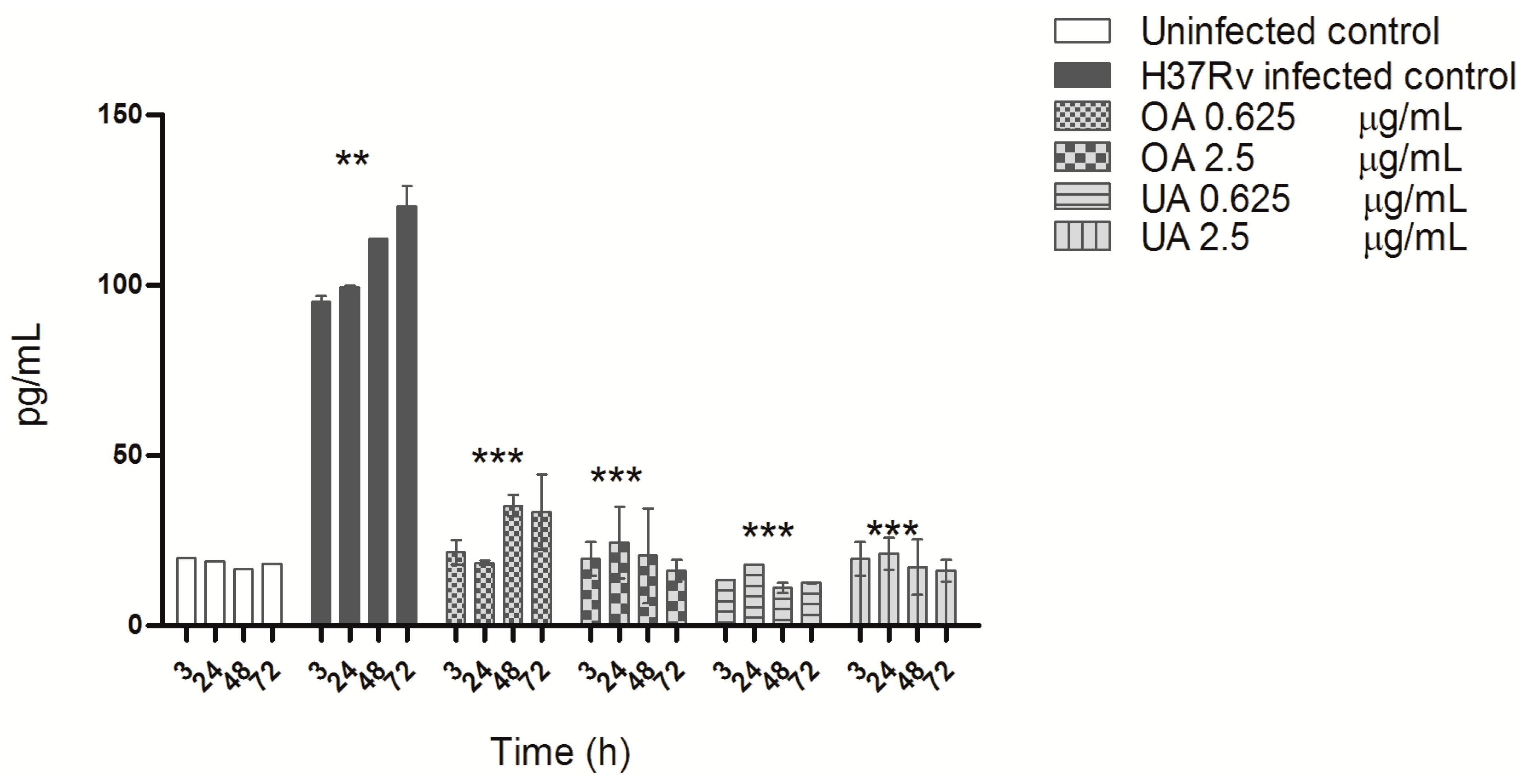
2.4. Cytokine Production during Mycobacterial Infection and Triterpene Treatment


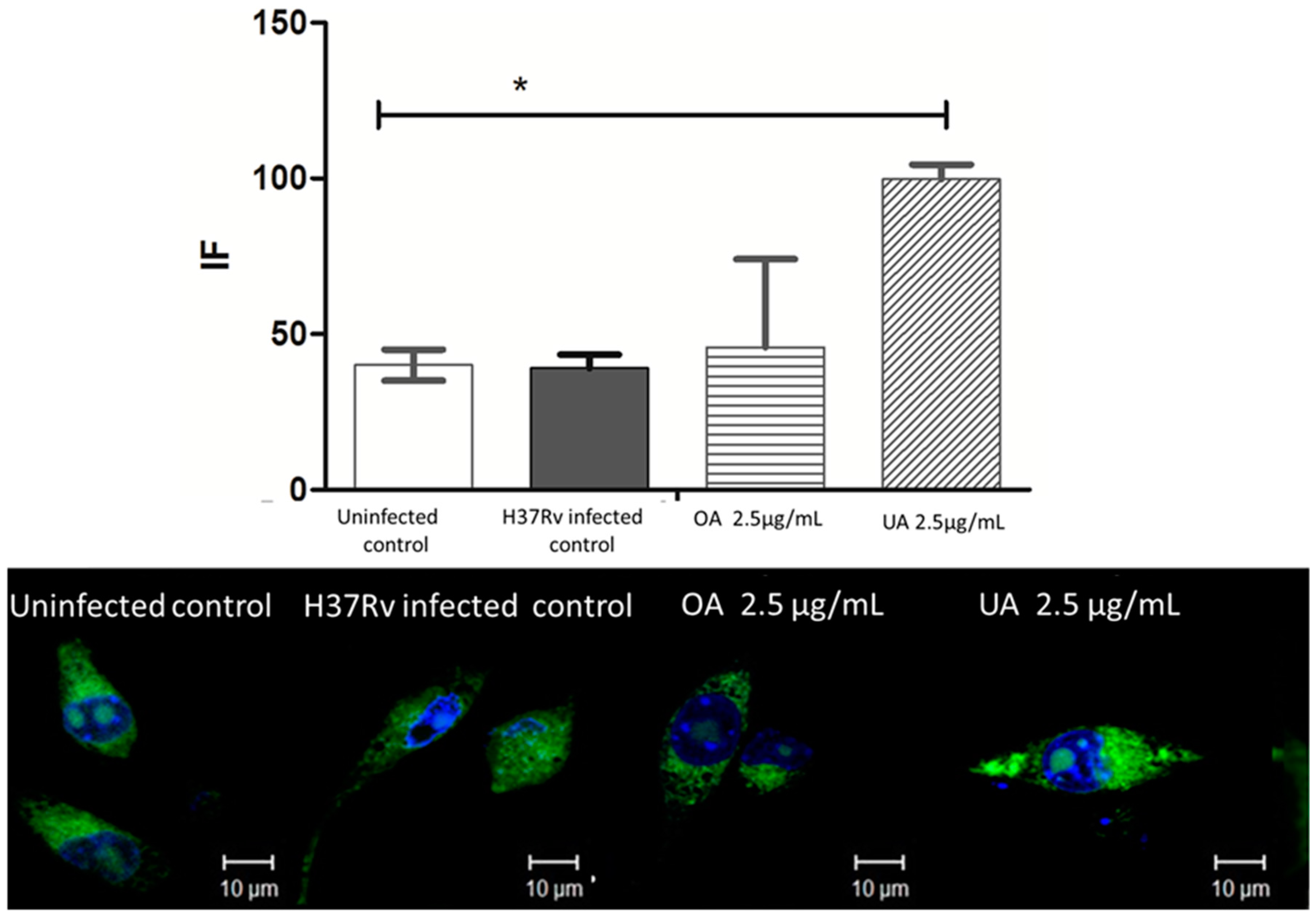
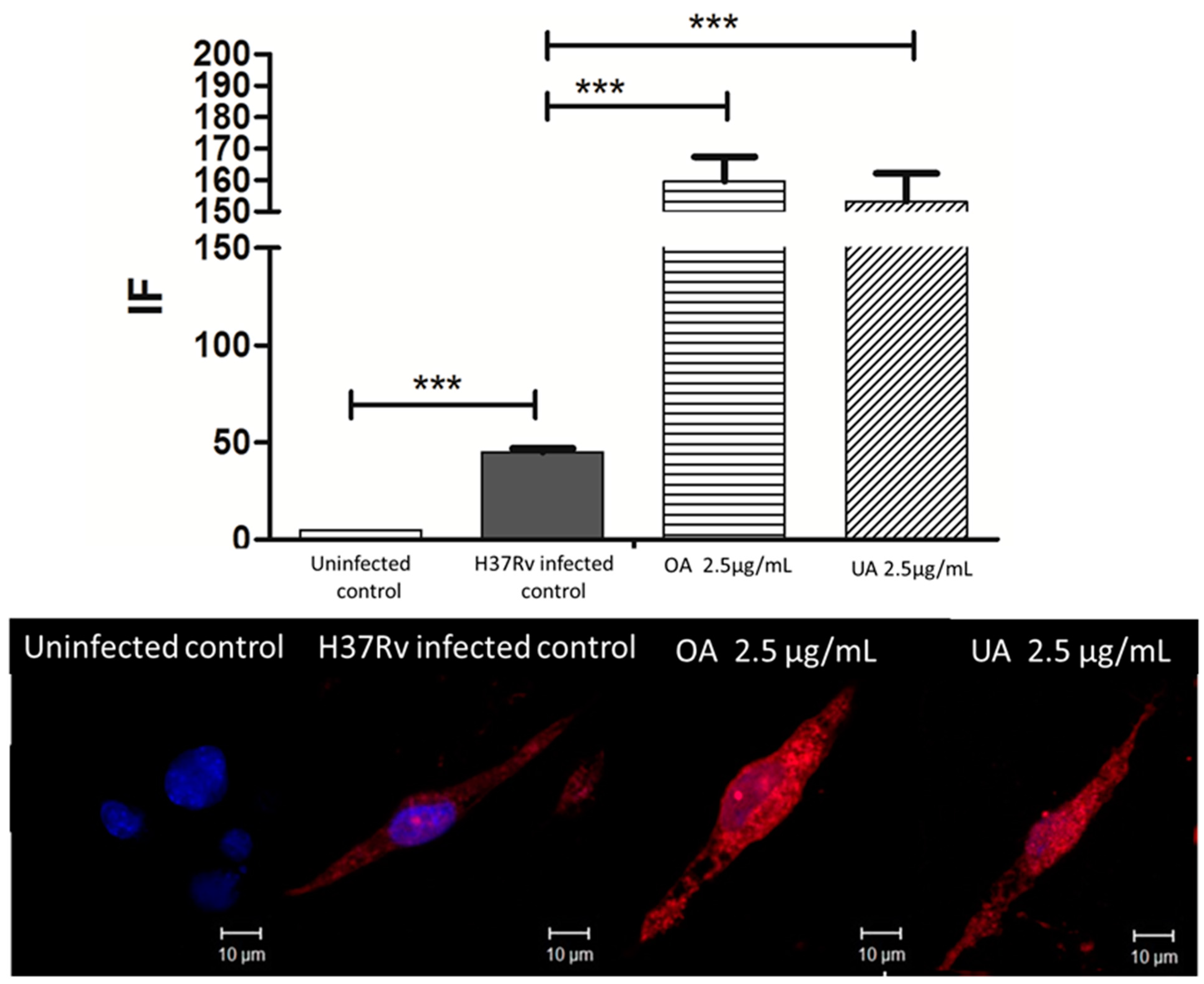
2.5. Determination of Expression of CD36 and TGR5 Receptors in Macrophages Infected by M. tuberculosis H37Rv and Treated with UA and OA

3. Experimental Section
3.1. Reagents
3.2. Cell Culture

3.3. Cytotoxicity and Intracellular Activity Assay
3.4. Intracellular Antimycobacterial Activity of OA and UA
3.5. Reactive Oxygen Species (ROS) Determination
3.6. NO Production
3.7. Quantification of Cytokines (TNF-α and TGF-β) in Supernatants by ELISA
3.8. Evaluation of CD36 and Receptor TGR5 Expression in Macrophages Infected with M. tuberculosis H37Rvand Treated with OA and UA for 24 h
3.9. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Du, H.; Wu, J.; Li, H.; Zhong, P.X.; Xu, Y.J.; Li, C.H.; Ji, K.X.; Wang, L.S. Polyphenols and triterpenes from Chaenomeles fruits: Chemical analysis and antioxidant activities assessment. Food Chem. 2013, 15, 4260–4268. [Google Scholar] [CrossRef] [PubMed]
- Muller, D.; Triebel, S.; Rudakovski, O.; Richling, E. Influence of triterpenoids present in apple peel on inflammatory gene expression associated with inflammatory bowel disease (IBD). Food Chem. 2013, 15, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Jäger, S.; Trojan, H.; Kopp, T.; Laszczyk, M.N.; Scheffler, A. Pentacyclic triterpene distribution in various plants—Rich sources for a new group of multi-potent plant extracts. Molecules 2009, 14, 2016–2031. [Google Scholar] [CrossRef] [PubMed]
- Liu, J. Oleanolic acid and ursolic acid: Research perspectives. J. Ethnopharmacol. 2005, 22, 92–94. [Google Scholar] [CrossRef] [PubMed]
- Jimenez-Arellanes, A.; Meckes, M.; Torres, J.; Luna-Herrera, J. Anti-mycobacterial triterpenoids from Lantana hispida (Verbenaceae). J. Ethnopharmacol. 2007, 111, 202–205. [Google Scholar] [CrossRef] [PubMed]
- Tanachatchairatana, T.; Bremner, J.B.; Chokchaisiri, R.; Suksamrarn, A. Antimycobacterial activity of cinnamate-based esters of the triterpenes betulinic, oleanolic and ursolic acids. Chem. Pharm. Bull. 2008, 56, 194–198. [Google Scholar] [CrossRef] [PubMed]
- Moraes, T.M.; de Araújo, M.H.; Bernardes, N.R.; de Oliveira, D.B.; Lasunskaia, E.B.; Muzitano, M.F.; da Cunha, M. Antimycobacterial activity and alkaloid prospection of Psychotria species (Rubiaceae) from the Brazilian Atlantic rainforest. Planta Medica 2011, 77, 964–970. [Google Scholar] [CrossRef] [PubMed]
- Fontanay, S.; Grare, M.; Mayer, J.; Finance, C.; Duval, R.E. Ursolic, oleanolic and betulinic acids: Antibacterial spectra and selectivity indexes. J. Ethnopharmacol. 2008, 120, 272–276. [Google Scholar] [CrossRef] [PubMed]
- Szakiel, A.; Ruszkowski, D.; Grudniak, A.; Kurek, A.; Wolska, K.; Doligalska, M.; Janiszowska, W. Antibacterial and antiparasitic activity of oleanolic acid and its glycosides isolated from marigold (Calendula officinalis). Planta Medica 2008, 74, 1709–1715. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.; Li, S.; Liao, Q.; Zhang, Y.; Sun, R.; Zhu, X.; Zhang, Q.; Wang, J.; Wu, X.; Fang, X.; et al. Oleanolic acid and ursolic acid: Novel hepatitis C virus antivirals that inhibit NS5B activity. Antiviral Res. 2013, 98, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Dufour, D.; Pichette, A.; Mshvildadze, V.; Bradette-Hébert, M.E.; Lavoie, S.; Longtin, A.; Laprise, C.; Legault, J. Antioxidant, anti-inflammatory and anticancer activities of methanolic extracts from Ledum groenlandicum Retzius. J. Ethnopharmacol. 2007, 111, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Shyamal, S.; Latha, P.G.; Suja, S.R.; Shine, V.J.; Anuja, G.I.; Sini, S.; Pradeep, S.; Shika, P.; Rajasekharan, S. Hepatoprotective effect of three herbal extracts on aflatoxin B1-intoxicated rat live. Singap. Med. J. 2010, 51, 326–331. [Google Scholar]
- Jimenez-Arellanes, A.; Meckes, M.; Alvarez, V.; Torres, J.; Parra, R. Secondary metabolites from Chamaedora tepejilote (Palmae) are active against Mycobacterium tuberculosis. Phytother. Res. 2005, 19, 320–322. [Google Scholar]
- Jung, T.Y.; Pham, T.N.; Umeyama, A.; Shoji, N.; Hashimoto, T.; Lee, J.J.; Takei, M. Ursolic acid isolated from Uncaria rhynchophylla activates human dendritic cells via TLR2 and/or TLR4 and induces the production of IFNγ by CD4+ naive T cells. Eur. J. Pharmacol. 2010, 643, 297–303. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Global Tuberculosis Report 2014. Available online: http://www.apps.who.int/iris/bitstream/10665/137094/1/9789241564809_eng.pdf (accessed on 6 April 2015).
- Ananthan, S.; Faaleolea, E.R.; Goldman, R.C.; Hobrath, J.V.; Kwong, C.D.; Laughon, B.E.; Maddry, J.A.; Mehta, A.; Rasmussen, L.; Reynolds, R.C.; et al. High-throughput screening for inhibitors of Mycobacterium tuberculosis H37Rv. Tuberculosis 2009, 89, 334–353. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Arellanes, A.; Luna-Herrera, J.; Cornejo-Garrido, J.; López-García, S.; Castro-Mussot, M.E.; Meckes-Fischer, M.; Mata-Espinosa, D.; Marquina, B.; Torres, J.; Hernández-Pando, R. Ursolic and oleanolic acids as antimicrobial and immunomodulatory compounds for tuberculosis treatment. BMC Complement. Altern. Med. 2013, 13, 258. [Google Scholar] [CrossRef] [PubMed]
- Saaby, L.M.; Nielsen, C.H. Triterpene acids from rose hip powder inhibit self-antigen- and LPS-induced cytokine production and CD4+ T-cell proliferation in human mononuclear cell cultures. Phytother. Res. 2012, 26, 1142–1147. [Google Scholar] [CrossRef] [PubMed]
- Podder, B.; Jang, W.S.; Nam, K.W.; Lee, B.E.; Song, H.Y. Ursolic acid activates intracellular killing effect of macrophages during Mycobacterium tuberculosis infection. J. Microbiol. Biotechnol. 2015, 25, 738–744. [Google Scholar] [PubMed]
- Rivero-Lezcano, O.M.; González-Cortés, C.; Reyes-Ruvalcaba, D.; Diez-Tascón, C. CCL20 is overexpressed in Mycobacterium tuberculosis-infected monocytes and inhibits the production of reactive oxygen species (ROS). Clin. Exp. Immunol 2010, 162, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Brzezinska, M.; Szulc, I.; Brzostek, A.; Klink, M.; Kielbik, M.; Sulowska, Z.; Pawelczyk, J.; Dziadek, J. The role of 3-ketosteroid 1(2)-dehydrogenase in the pathogenicity of Mycobacterium tuberculosis. BMC Microbiol. 2013, 13. [Google Scholar] [CrossRef] [PubMed]
- Chakravarti, B.; Maurya, R.; Siddiqui, J.A.; Bid, H.K.; Rajendran, S.M.; Yadav, P.P.; Konwar, R. In vitro anti-breast cancer activity of ethanolic extract of Wrightiatomentosa: Role of pro-apoptotic effects of oleanolic acid and ursolic acid. J. Ethnopharmacol. 2012, 142, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Fu, J.; Li, T.; Cui, R.; Ling, J.; Yu, X.; Ji, H.; Zhang, Y. A novel PABA/NO-based oleanolic acid derivative induces human hepatoma cell apoptosis via a ROS/MAPK-dependent mitochondrial pathway. Eur. J. Pharmacol. 2012, 15, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, B.M.; Kannan, N.; Vemu, L.; Raghunand, T.R. The Mycobacterium tuberculosis PE proteins Rv0285 and Rv1386 modulate innate immunity and mediate bacillary survival in macrophages. PLoS ONE 2012, 7, e51686. [Google Scholar] [CrossRef] [PubMed]
- Stewart, C.R.; Stuart, L.M.; Wilkinson, K.; van Gils, J.M.; Deng, J.; Halle, A.; Rayner, K.J.; Boyer, L.; Zhong, R.; Frazier, W.A.; et al. CD36 ligands promote sterile inflammation through assembly of a Toll-like receptor 4 and 6 heterodimer. Nat. Immunol. 2010, 11, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Zhang, Y.; Zhao, L.; Jing, Y.; Lai, Y.; Zhang, L.; Guo, Q.; Yuan, S.; Zhang, J.; Chen, L.; et al. Synthesis and anti-human hepatocellular carcinoma activity of new nitric oxide-releasing glycosyl derivatives of oleanolic acid. Org. Biomol. Chem. 2010, 8, 632–639. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.; Gowthaman, U.; Pahari, S.; Agrewala, J.N. Manipulation of coestimulatory U molecules by intracellular pathogens: Veni, vidi, vici!! PLoS Pathogen. 2012, 8, e1002676. [Google Scholar] [CrossRef] [PubMed]
- Tomioka, H.; Tatano, Y.; Maw, W.W.; Sano, C.; Kanehiro, Y.; Shimizu, T. Characteristics of suppressor macrophages induced by mycobacterial and protozoa infections in relation to alternatively activated M2 macrophages. Clin. Dev. Immunol. 2012, 2012. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, Y.; Murakami, A.; Ohigashi, H. Ursolic acid: An anti- and pro-inflammatory triterpenoid. Mol. Nutr. Food Res. 2008, 52, 26–42. [Google Scholar] [CrossRef] [PubMed]
- Choi, C.Y.; You, H.J.; Jeong, H.G. Nitric oxide and tumor necrosis factor-alfa production by oleanolic acid via nuclear factor kappa b activation in macrophages. Biochem. Biophys. Res. Commun. 2001, 288, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Ríos, J.L. Effects of triterpenes on the immune system. J. Ethnopharmacol. 2010, 128, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Szuster-Ciesielska, A.; Plewka, K.; Daniluk, J.; Kandefer-Szerszeń, M. Betulin and betulinic acid attenuate ethanol-induced liver stellate cell activation by inhibiting reactive oxygen species (ROS), cytokine (TNF-α, TGF-β) production and by influencing intracellular signaling. Toxicology 2011, 27, 152–163. [Google Scholar] [CrossRef] [PubMed]
- Lugo-Villarino, G.; Vérollet, C.; Maridonneau-Parini, I.; Neyrolles, O. Macrophage polarization: Convergence point targeted by Mycobacterium tuberculosis and HIV. Front. Immunol. 2011, 2, 43. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, Y.; Murakami, A.; Fujimura, Y.; Tachibana, H.; Yamada, K.; Masuda, D.; Hirano, H.; Yamashita, S.; Ohigashi, H. Aggregated ursolic acid, a natural triterpenoid, induces IL-1beta release from murine peritoneal macrophages: role of CD36. J. Immunol. 2007, 178, 4854–4864. [Google Scholar] [CrossRef] [PubMed]
- Genet, C.; Strehle, A.; Schmidt, C.; Boudjelal, G.; Lobstein, A.; Schoonjans, K.; Souchet, M.; Auwerx, J.; Saladin, R.; Wagner, A. Structure-activity relationship study of betulinic acid, a novel and selective TGR5 agonist, and its synthetic derivatives: Potential impact in diabetes. J. Med. Chem. 2010, 53, 178–190. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, K.; Boyd, J.D.; Glicksman, M.; Moore, K.J.; El-Khoury, J. A high content drug screen identifies ursolic acid as an inhibitor of amyloid β protein interactions with its receptor CD36. J. Biol. Chem. 2011, 286, 34914–34922. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.M. CD36, a scavenger receptor implicated in atherosclerosis. Exp. Mol. Med. 2014, 46, e99. [Google Scholar] [CrossRef] [PubMed]
- Mattos, K.A.; Oliveira, V.C.; Berrêdo-Pinho, M.; Amaral, J.J.; Antunes, L.C.; Melo, R.C.; Acosta, C.C.; Moura, D.F.; Olmo, R.; Han, J.; et al. Mycobacterium leprae intracelular survival relies on colesterol accumulation in infected macrophages: A potential target for new drugs for leprosy treatment. Cell Microbiol. 2014, 16, 797–815. [Google Scholar] [CrossRef] [PubMed]
- Hawkes, M.; Li, X.; Crockett, M.; Diassiti, A.; Finney, C.; Min-Oo, G.; Liles, W.C.; Liu, J.; Kain, K.C. CD36 deficiency attenuates experimental mycobacterial infection. BMC Infect. Dis. 2010, 10, 299. [Google Scholar] [CrossRef] [PubMed]
- Triantafilou, M.; Gamper, F.G.; Haston, R.M.; Mouratis, M.A.; Morath, S.; Hartung, T.; Triantafilou, K. Membrane sorting of toll-like receptor (TLR)-2/6 and TLR2/1 heterodimers at the cell surface determines heterotypic associations with CD36 and intracellular targeting. J. Biol. Chem. 2006, 281, 31002–3111. [Google Scholar] [CrossRef] [PubMed]
- Drage, M.G.; Pecora, N.D.; Hise, A.G.; Febbraio, M.; Silverstein, R.L.; Golenbock, D.T.; Boom, W.H.; Harding, C.V. TLR2 and its co-receptors determine responses of macrophages and dendritic cells to lipoproteins of Mycobacterium tuberculosis. Cell Immunol. 2009, 258, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.; Gioiello, A.; Noriega, L.; Strehle, A.; Oury, J.; Rizzo, G.; Macchiarulo, A.; Yamamoto, H.; Mataki, C.; Pruzanski, M.; et al. TGR5-mediated bile acid sensing controls glucose homeostasis. Cell Metab. 2009, 10, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Duboc, H.; Taché, Y.; Hofmann, A.F. The bile acid TGR5 membrane receptor: From basic research to clinical application. Dig. Liver Dis. 2014, 46, 302–312. [Google Scholar] [CrossRef] [PubMed]
- Keitel, V.; Reinehr, R.; Gatsios, P.; Rupprecht, C.; Görg, B.; Selbach, O.; Häussinger, D.; Kubitz, R. The G-protein coupled bile salt receptor TGR5 is expressed in liver sinusoidal endothelial cells. Hepatology 2007, 45, 695–704. [Google Scholar] [CrossRef] [PubMed]
- Eggink, H.M.; Soeters, M.R.; Pols, T.W. TGR5 ligands as potential therapeutics in inflammatory diseases. Intl. J. Interferon Cytokine Mediat. Res. 2014, 6, 27–38. [Google Scholar]
- Wang, Y.D.; Chen, W.D.; Yu, D.; Forman, B.M.; Huang, W. The G-protein coupled bile acid receptor Gpbar1 (TGR5) negatively regulates hepatic inflammatory response through antagonizing Nuclear Factor κB. Hepatology 2011, 54, 1421–1432. [Google Scholar] [CrossRef] [PubMed]
- Lou, G.; Ma, X.; Fu, X.; Meng, Z.; Zhang, W.; Wang, Y.D.; Van Ness, C.; Yu, D.; Xu, R.; Huang, W. GPBAR1/TGR5 mediates bile acid-induced cytokine expression in murine Kupffer cells. PLoS ONE 2014, 9, e93567. [Google Scholar] [CrossRef] [PubMed]
- Calmus, Y.; Poupon, R. Shaping macrophages function and innate immunity by bile acids: Mechanisms and implication in cholestatic liver diseases. Clin. Res. Hepatol. Gastroenterol. 2014, 38, 550–556. [Google Scholar] [CrossRef] [PubMed]
- Kida, T.; Tsubosaka, Y.; Hori, M.; Ozaki, H.; Murata, T. Bile acid receptor TGR5 agonism induces NO production and reduces monocyte adhesion in vascular endothelial cells. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 1663–1669. [Google Scholar] [CrossRef] [PubMed]
- Mobraten, K.; Haugbro, T.; Karlstrom, E.; Kleiveland, C.R.; Lea, T. Activation of the bile acid receptor TGR5 enhances LPS-induced inflammatory responses in a human monocytic cell line. J. Recept. Signal Transduct. Res. 2014, 24, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Forge, D.; Cappoen, D.; Laurent, J.; Stanicki, D.; Mayence, A.; Huang, T.L.; Verschaeve, L.; Huygen, K.; Van den Eynde, J.J. 1,4-diarylpiperazines and analogs as anti-tubercular agents: Synthesis and biological evaluation. Eur. J. Med. Chem. 2012, 49, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Pauli, G.F.; Case, R.J.; Inui, T.; Wang, Y.; Cho, S.; Fischer, N.H.; Franzblau, S.G. New perspectives on natural products in TB drug research. Life Sci. 2005, 78, 485–494. [Google Scholar] [CrossRef] [PubMed]
- Luna-Herrera, J.; Reddy, M.V.; Gangadharam, P.R. In-vitro and intracellular activity of rifabutin on drug-susceptible and multiple drug-resistant (MDR) tubercle bacilli. J. Antimicrob. Chemother. 1995, 36, 355–363. [Google Scholar] [CrossRef] [PubMed]
- García-Pérez, B.A.; Villagómez-Palatto, D.A.; Castañeda-Sánchez, J.I.; Coral-Vázquez, R.; Ramírez-Sánchez, I.; Ordoñez-Razo, R.M.; Luna-Herrera, J. Innate response of human endothelial cells infected with mycobacteria. Immunobiology 2011, 216, 925–935. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Not available.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
López-García, S.; Castañeda-Sanchez, J.I.; Jiménez-Arellanes, A.; Domínguez-López, L.; Castro-Mussot, M.E.; Hernández-Sanchéz, J.; Luna-Herrera, J. Macrophage Activation by Ursolic and Oleanolic Acids during Mycobacterial Infection. Molecules 2015, 20, 14348-14364. https://doi.org/10.3390/molecules200814348
López-García S, Castañeda-Sanchez JI, Jiménez-Arellanes A, Domínguez-López L, Castro-Mussot ME, Hernández-Sanchéz J, Luna-Herrera J. Macrophage Activation by Ursolic and Oleanolic Acids during Mycobacterial Infection. Molecules. 2015; 20(8):14348-14364. https://doi.org/10.3390/molecules200814348
Chicago/Turabian StyleLópez-García, Sonia, Jorge Ismael Castañeda-Sanchez, Adelina Jiménez-Arellanes, Lilia Domínguez-López, Maria Eugenia Castro-Mussot, Javier Hernández-Sanchéz, and Julieta Luna-Herrera. 2015. "Macrophage Activation by Ursolic and Oleanolic Acids during Mycobacterial Infection" Molecules 20, no. 8: 14348-14364. https://doi.org/10.3390/molecules200814348
APA StyleLópez-García, S., Castañeda-Sanchez, J. I., Jiménez-Arellanes, A., Domínguez-López, L., Castro-Mussot, M. E., Hernández-Sanchéz, J., & Luna-Herrera, J. (2015). Macrophage Activation by Ursolic and Oleanolic Acids during Mycobacterial Infection. Molecules, 20(8), 14348-14364. https://doi.org/10.3390/molecules200814348