
Antifungal Activity of Isoliquiritin and Its Inhibitory Effect against Peronophythora litchi Chen through a Membrane Damage Mechanism

Abstract
:1. Introduction
2. Results
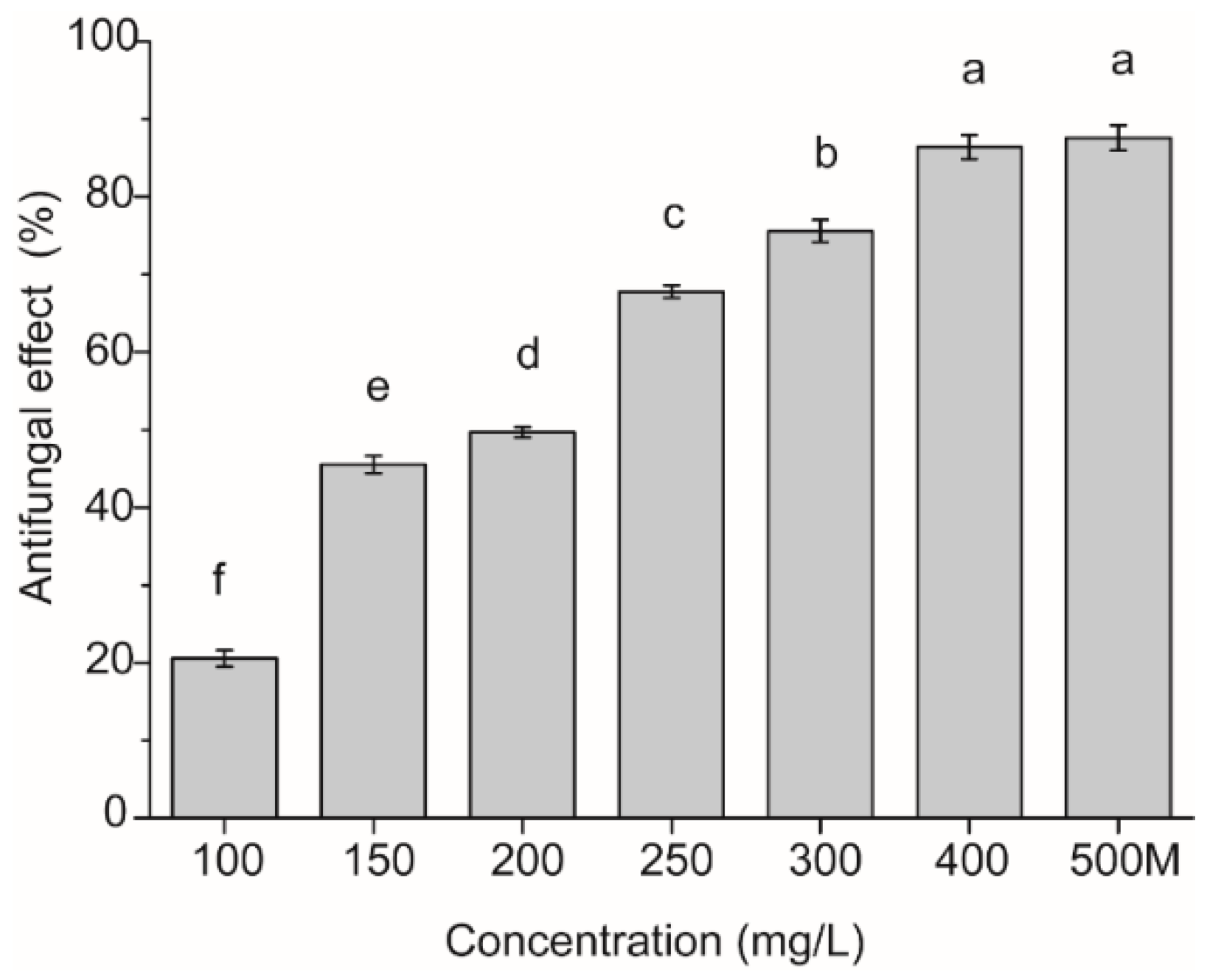
2.1. Antifungal Activity
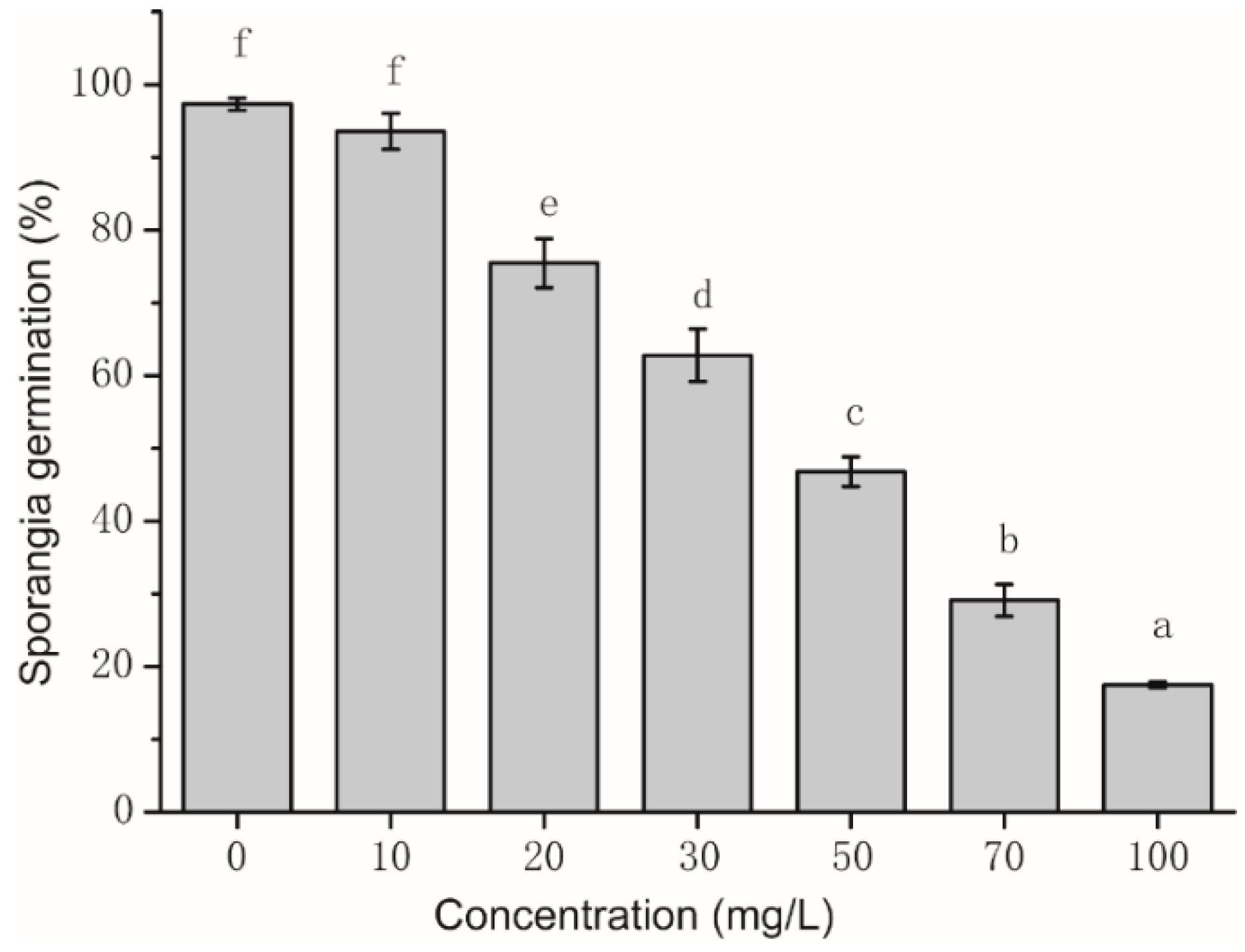
2.2. Assay of Sporangia Germination and Influence of Sporangia Morphology
2.3. Detached Leaf Test
2.4. Scanning Electron Microscopy (SEM)
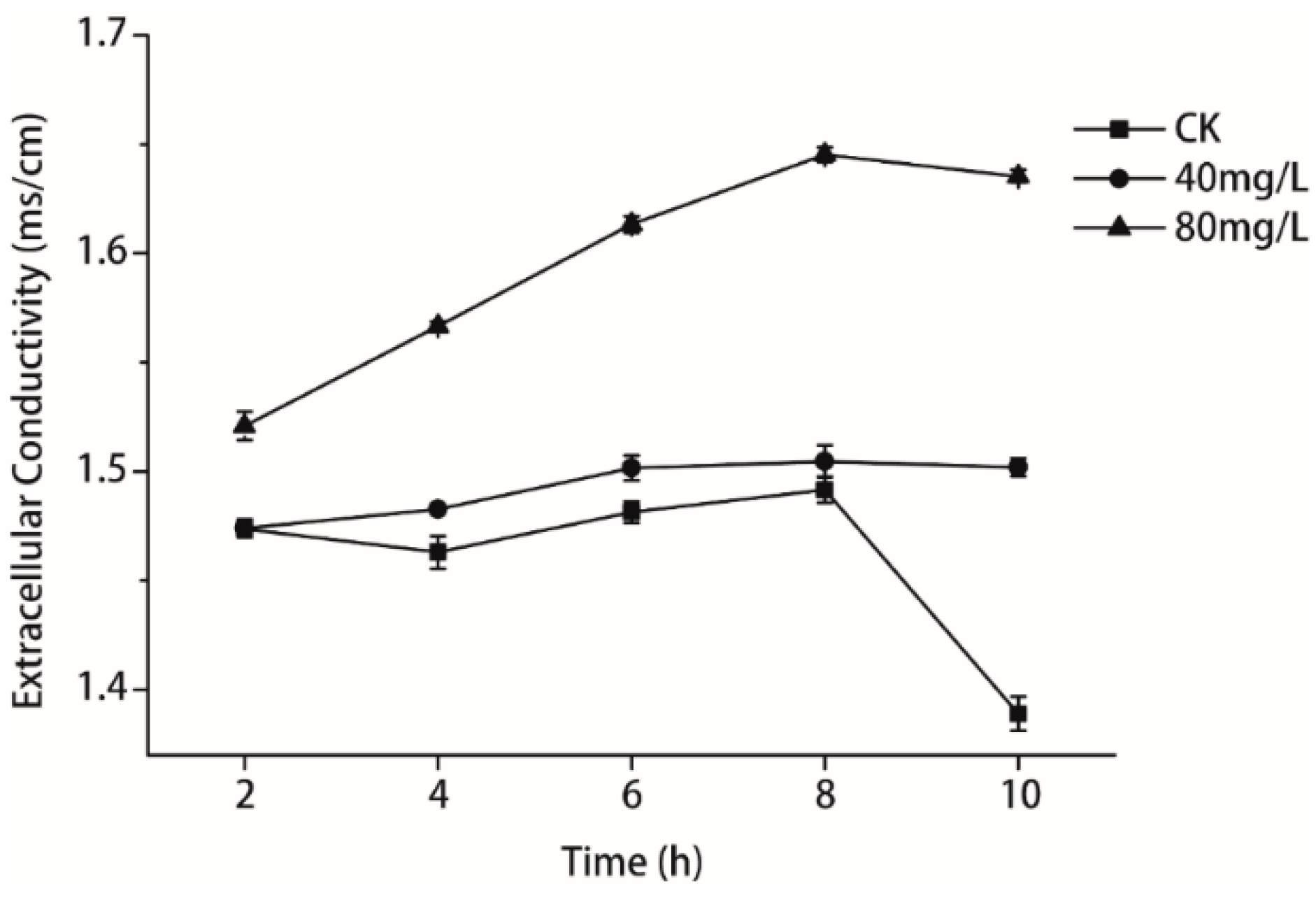
2.5. Measurement of Relative Electric Conductivity
2.6. Determination of Reducing Sugar
3. Discussion
4. Experimental Section
4.1. Fungal Species
4.2. Plant Material
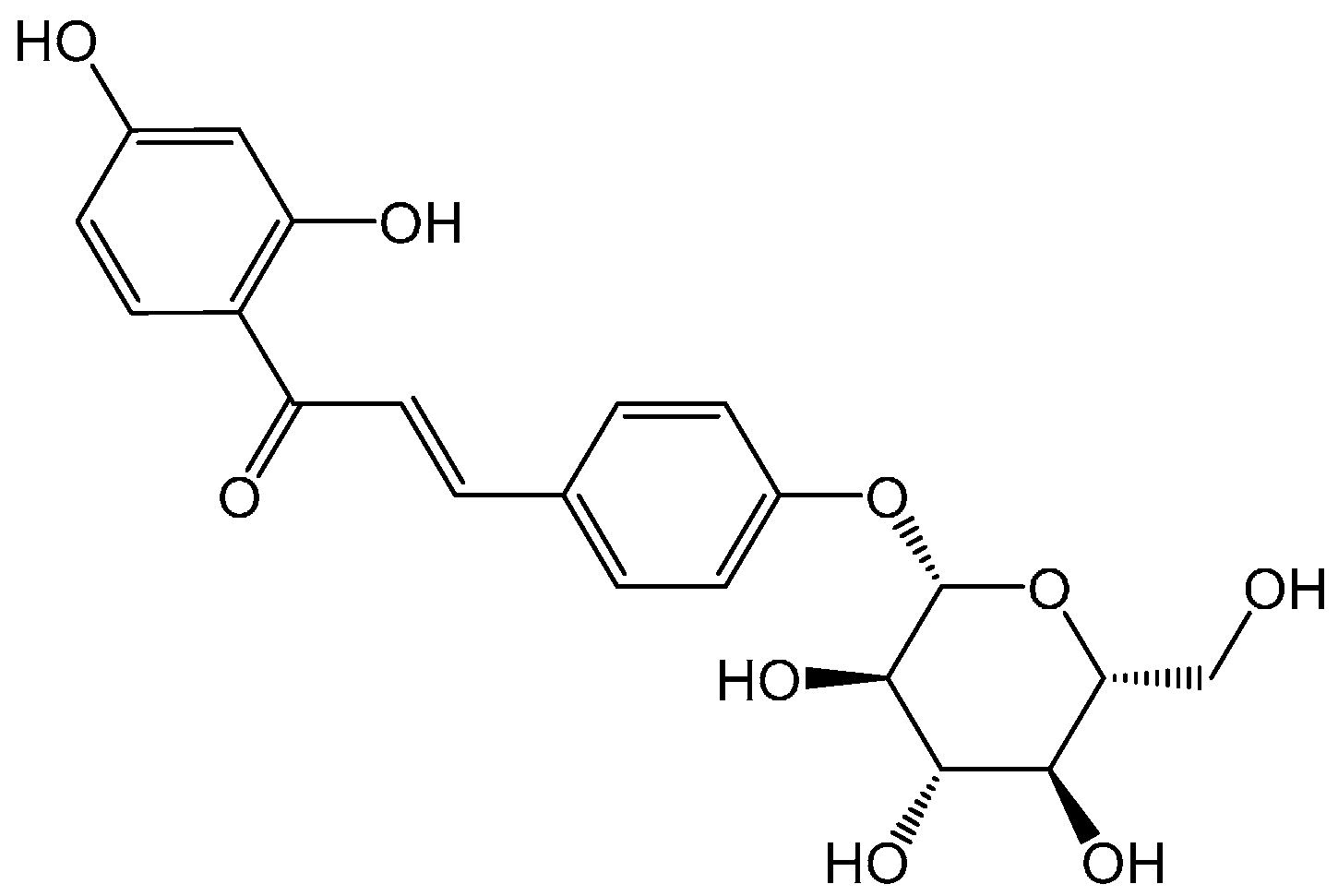
4.3. Chemicals
4.4. Antifungal Activity
4.5. Assay of Sporangia Germination and Influence of Sporangia Morphology
4.6. Detached Leaf Test
4.7. Scanning Electron Microscopy (SEM)
4.8. Measurement of Relative Electric Conductivity
4.9. Determination of Reducing Sugar
4.10. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Yang, B.; Zhao, M.M.; Liu, Y.; Li, B.Z. Characterization of litchi pericarp polysaccharide. Nat. Prod. Res. Dev. 2005, 17, 685–687. [Google Scholar]
- Chyau, C.C.; Ko, P.T.; Chang, C.H.; Mau, J.L. Free and glycosidically bound aroma compounds in lychee (Litchi chinensis Sonn.). Food Chem. 2003, 80, 387–392. [Google Scholar] [CrossRef]
- Javier, R.L.; Cesar, O.F.; Pedro, W.E. Changes in anthocyanin concentration in litchi pericarp during maturation. Food Chem. 1999, 65, 195–200. [Google Scholar]
- Hang, X.M. Lychee production in China. In Lychee Production in the Asia-Pacific Region; FAO/RAP Press: Bangkok, Thailand, 2001; pp. 41–45. [Google Scholar]
- Chi, P.K.; Pan, X.P.; Liu, R. On downy blight of Litchi chinensis Sonn. The pathogen and its infection process. Acta Phytopathol. Sin. 1984, 14, 113–119. [Google Scholar]
- Chaiwatana, K.; Manoch, T.; Vichai, R.; Kachornsak, B. Peronophythora litchii associated with root rot of litchi. In Proceedings of the National Plant Protection Conference, Chiang Mai, Thailand, 9–11 October 1995; pp. 9–11.
- Tang, Z.H.; Wang, H.C.; Hou, Y.P.; Zhang, S.P.; Wang, J.X.; Zhou, M.G. Baseline and differential sensitivity to mandipropamid among isolates of Peronophythora litchii, the causal agent of downy blight on litchi. Crop Prot. 2011, 30, 354–359. [Google Scholar] [CrossRef]
- Wang, H.C.; Sun, H.Y.; Ma, J.X.; Stammler, G.; Zhou, M.G. Fungicide effectiveness during the various developmental stages of Peronophythora litchii in vitro. J. Phytopathol. 2009, 157, 407–412. [Google Scholar] [CrossRef]
- Serrati, L.; Cestari, P.; Huggenberger, F.; Lamberth, C.; Iwanzik, W.; Knauf-Beiter, G. Mandipropamid: New fungicide against oomycete pathogens. In Proceedings of the Phytopathological Meeting, Riccione, Italy, 27–29 March 2006; Volume 2, pp. 9–16.
- Harp, T.; Cochran, A.; Tory, D.; Kuhn, P.; Payan, L.; Laird, D.; Tally, A. Development of Revus 2.09 SC (ai mandipropamid) in the US for control of downy mildews on leafy vegetables. Phytopathology 2007, 97, S45. [Google Scholar]
- Wang, H.C.; Sun, H.Y.; Stammler, G.; Ma, J.X.; Zhou, M.G. Baseline and differential sensitivity of Peronophythora litchii (lychee downy blight) to three carboxylic acid amide fungicides. Plant Pathol. 2009, 58, 571–576. [Google Scholar] [CrossRef]
- Huggenberger, F.; Lamberth, C.; Iwanzik, W.; Knauf-Beiter, G. Mandipropamid a new fungicide against oomycete pathogens. In Proceedings of the BCPC Internat Congress, Alton, Hants, UK, 31 October–2 November 2005; pp. 93–98.
- Rubin, E.; Gotlieb, D.; Gisi, U.; Cohen, Y. Mutagenesis of Phytophthora infestans for resistance against carboxylic acid amide (CAA) and phenylamide fungicides. Plant Dis. 2008, 92, 675–683. [Google Scholar] [CrossRef]
- State Pharmacopoeia Committee. Pharmacopoeia of China, Part 1; Chemical Industry Press: Beijing, China, 2010; pp. 80–81. [Google Scholar]
- Liu, Y.; Zhu, X.Q.; Li, W.D.; Wen, H.; Liu, C.S. Enhancing ergosterol production in Pichia pastoris GS115 by overexpressing squalene synthase gene from Glycyrrhiza uralensis. Chin. J. Nat. Med. 2015, 13, 338–345. [Google Scholar] [CrossRef]
- Fu, Y.; Chen, J.; Li, Y.J.; Zheng, Y.F.; Li, P. Antioxidant and antiinflammatory activities of six flavonoids separated from licorice. Food Chem. 2013, 141, 1063–1071. [Google Scholar] [CrossRef] [PubMed]
- Jatinder, P.S.; Amritpal, K.; Narpinder, S.; Lovedeep, N.; Khetan, S.; Harpreet, K.; Daljit, S.A. In vitro antioxidant and antimicrobial properties of jambolan (Syzygium cumini) fruit polyphenols. LWT Food Sci. Technol. 2016, 5, 1025–1030. [Google Scholar]
- Spencer, J.P.; Abd El Mohsen, M.M.; Minihane, A.M.; Mathers, J.C. Biomarkers of the intake of dietary polyphenols: Strengths, limitations and application in nutrition research. Br. J. Nutr. 2008, 99, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, S.; Miyamoto, T.; Kimura, I.; Kimura, M. Inhibitory effect of isoliquiritin, a compound in licorice root, on angiogenesis in vivo and tube formation in vitro. Biol. Pharm. Bull. 1995, 18, 1382–1386. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.X.; Hua, X.Y.; Zhao, Z.Y.; Liu, P.; Hu, Y.C.; Zhou, J.P.; Zhou, D.F.; Wang, Z.B.; Guo, D.; Guo, H.Z. Antidepressant-like effects of liquiritin and isoliquiritin from Glycyrrhiza uralensis in the forced swimming test and tail suspension test in mice. Prog. Neuropsychopharmacol. Biol. Psychiatry 2008, 32, 1179–1184. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.L.; Ho, W.S. Combination of liquiritin, isoliquiritin and isoliquirigenin induce apoptotic cell death through upregulating p53 and p21 in the A549 non-small cell lung cancer cells. Oncol. Rep. 2013, 31, 298–304. [Google Scholar] [PubMed]
- Wang, R.; Zhang, C.Y.; Bai, L.P.; Pan, H.D.; Shu, L.M.; Kong, A.T.; Leung, E.L.; Liu, L.; Li, T. Flavonoids derived from liquorice suppress murine macrophage activation by up-regulating heme oxygenase-1 independent of Nrf2 activation. Int. Immunopharmacol. 2015, 28, 917–924. [Google Scholar] [CrossRef] [PubMed]
- Bajpai, V.K.; AI-Reza, S.M.; Choi, U.K.; Lee, J.H.; Kang, S.C. Chemical composition, antibacterial and antioxidant activities of leaf essential oil and extracts of Metasequioa glyptostroboides Miki ex Hu. Food Chem. Toxicol. 2009, 47, 1876–1883. [Google Scholar] [CrossRef] [PubMed]
- Daglia, M. Polyphenols as antimicrobial agents. Curr. Opin. Biotechnol. 2012, 23, 174–181. [Google Scholar] [CrossRef] [PubMed]
- Renuka, J.; Umesh, S.; Kiran, R.; Satish, C.J. Bioactivities of polyphenolics from the roots of Bauhinia racemosa. Arch. Pharm. Res. 2008, 31, 1525–1529. [Google Scholar]
- Varsha, V.; Asna, U.; Ng, M. Evaluation of antioxidant and antimicrobial properties of finger millet polyphenols (Eleusine coracana). Food Chem. 2009, 114, 340–346. [Google Scholar]
- Shangmugen, S.; Thangaraj, P. In vitro antioxidant, antimicrobial and anti-diabetic properties of polyphenols of Passiflora ligularis Juss. fruit pulp. Food Sci. Hum. Wellness 2014, 3, 56–64. [Google Scholar]
- Cox, S.D.; Mann, C.M.; Markham, J.L.; Gustafson, J.E.; Warmington, J.R.; Wyllie, S.G. Determining the antimicrobial actions of tea tree oil. Molecules 2001, 6, 87–91. [Google Scholar] [CrossRef]
- Sharma, A.; Bajpai, V.K.; Baek, K.-H. Determination of antibacterial mode of action of allium sativum essential oil against foodborne pathogens using membrane permeability and surface characteristic parameters. J. Food Saf. 2013, 33, 197–208. [Google Scholar] [CrossRef]
- Li, F.D.; Hu, Z.J. Microbiology, 5th ed.; China Agriculture Press: Beijing, China, 2000; p. 3208. [Google Scholar]
- Mach, R.L.; Peterbauer, C.K.; Payer, K.; Jaksits, S.; Woo, S.L.; Zeilinger, S.; Kullnig, C.M.; Lorito, M.; Kubicek, C.P. Expression of two major chitinase genes of Trichoderma atroviride (T. harzianum P1) is triggered by different regulatory signals. Appl. Environ. Microbiol. 1999, 65, 1858–1863. [Google Scholar] [PubMed]
- Allen, P.M.; Gottlieb, D. Mechanism of action of the fungicide thiabendazole, 2-(4-thiazolyl) benzimidazole. Appl. Microbiol. 1970, 20, 919–926. [Google Scholar] [PubMed]
- Yahyazadeh, M.; Omidbaigi, R.; Zare, R.; Taheri, H. Effect of some essentialoils on mycelial growth of Penicillium digitatum Sacc. World J. Microbiol. Biotechnol. 2008, 24, 1445–1450. [Google Scholar] [CrossRef]
- Helal, G.A.; Sarhan, M.M.; Abu, S.A.N.; Abou, E.-K.E.K. Effects of Cymbopogon citratus L. essential oil on the growth, morphogenesis and afflatoxin production of Aspergillus flavus ML2-strain. J. Basic Microbiol. 2007, 47, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Paul, S.; Dubey, R.C.; Maheswari, D.K.; Kang, S.C. Trachyspermum ammi (L.) fruit essential oil influencing on membrane permeability and surface characteristics in inhibiting food-borne pathogens. Food Control. 2011, 22, 725–731. [Google Scholar] [CrossRef]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Yuan, Z.; Cong, G.; Zhang, J. Effects of exogenous salicylic acid on polysaccharides production of Dendrobium officinale. S. Afr. J. Bot. 2014, 95, 78–84. [Google Scholar] [CrossRef]
- Chung, R.S.; Chen, C.-C.; Ng, L.-T. Nitrogen fertilization affects the growth performance, betaine and polysaccharide concentrations of Lycium barbarum. Ind. Crops Prod. 2010, 32, 650–655. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the compounds are not available from the authors.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Concentration (mg/L) | Inhibition Activity (%) | |||
|---|---|---|---|---|
| PL | PC | SS | CH | |
| 1 | 3.98 ± 0.74 d | 9.09 ± 0.39 e | 20.84 ± 0.69 bc | 20.39 ± 0.15 b |
| 5 | 10.86 ± 0.39 cd | 18.17 ± 1.08 d | 24.17 ± 0.35 bc | 22.84 ± 0.36 b |
| 10 | 22.61 ± 0.89 c | 33.71 ± 0.41 c | 27.93 ± 0.61 bc | 26.59 ± 0.86 b |
| 50 | 66.29 ± 0.87 b | 63.85 ± 0.41 b | 41.46 ± 0.34 b | 34.80 ± 0.76 b |
| 100 | 83.58 ± 0.31 ab | 81.82 ± 0.37 ab | 48.12 ± 0.47 b | 40.13 ± 0.35 ab |
| 200 | 96.45 ± 0.20 a | 93.35 ± 0.04 a | 51.21 ± 0.39 ab | 43.45 ± 0.33 ab |
| 400 | 100.00 a | 97.34 ± 0.39 a | 64.31 ± 1.01 a | 53.22 ± 0.41 a |
| Fungal Pathogen | Y = a + bx | Correlation Coefficient | EC50 (mg/L) | 95% Confidence Limits (mg/L) |
|---|---|---|---|---|
| PL | Y = 1.7583 + 2.2563x | 0.9791 | 27.33 | 15.49–48.23 |
| PC | Y = 2.9663 + 1.5035x | 0.9924 | 22.52 | 19.37–44.17 |
| SS | Y = 3.0569 + 0.7884x | 0.9907 | 291.42 | 176.08–356.28 |
| CH | Y = 3.9245 + 0.4195x | 0.9836 | 367.79 | 268.24–417.65 |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, J.; Li, Z.; Wang, J.; Weng, Q.; Chen, S.; Hu, M. Antifungal Activity of Isoliquiritin and Its Inhibitory Effect against Peronophythora litchi Chen through a Membrane Damage Mechanism. Molecules 2016, 21, 237. https://doi.org/10.3390/molecules21020237
Luo J, Li Z, Wang J, Weng Q, Chen S, Hu M. Antifungal Activity of Isoliquiritin and Its Inhibitory Effect against Peronophythora litchi Chen through a Membrane Damage Mechanism. Molecules. 2016; 21(2):237. https://doi.org/10.3390/molecules21020237
Chicago/Turabian StyleLuo, Jianjun, Zhibin Li, Jingjing Wang, Qunfang Weng, Shaohua Chen, and Meiying Hu. 2016. "Antifungal Activity of Isoliquiritin and Its Inhibitory Effect against Peronophythora litchi Chen through a Membrane Damage Mechanism" Molecules 21, no. 2: 237. https://doi.org/10.3390/molecules21020237
APA StyleLuo, J., Li, Z., Wang, J., Weng, Q., Chen, S., & Hu, M. (2016). Antifungal Activity of Isoliquiritin and Its Inhibitory Effect against Peronophythora litchi Chen through a Membrane Damage Mechanism. Molecules, 21(2), 237. https://doi.org/10.3390/molecules21020237