
Natural Products from Chinese Medicines with Potential Benefits to Bone Health


Abstract
:1. Introduction
2. Indicators of Bone Health
2.1. Bone Formation Markers
2.2. Bone Resorption Markers
3. Natural Products with Osteoprotective and Related Activities
3.1. Carthami Flos (Flower of Carthamus tinctorius L.)
3.2. Cimicifugae Rhizoma (Rhizome of Actaea heracleifolia (Kom.) J. Compton [syn. Cimicifuga heracleifolia Kom.], A. dahurica (Turcz. ex Fisch. and C.A. Mey.) Franch. [syn. C. dahurica (Turcz.) Maxim.], or A. foetida L. [syn. C. foetida L.])
3.3. Cistanches Herba [Stem of Cistanche deserticola Y.C. Ma or C. tubulosa (Forssk.) Beck]
3.4. Cordyceps [Cordyceps sinensis (Berk.) Sacc.]
3.5. Dipsaci Radix (Root of Dipsacus asper Wall. ex C.B. Clarke [syn. D. asperoides C.Y. Cheng and T.M. Ai] or D. japonicas Miq.)
3.6. Drynariae Rhizoma [Rhizome of Drynaria fortunei (Kunze ex Mett.) J. Sm. (syn. D. roosii Nakaike)]
3.7. Ecliptae Herba (Above-Ground Parts of Eclipta prostrata L.)
3.8. Epimedii Folium [Leaf of Epimedium brevicornum Maxim., E. sagittatum (Siebold and Zucc.) Maxim., E. pubescens Maxim. or E. koreanum Nakai]
3.9. Erythrina variegata L.
3.10. Eucommiae Cortex (Stem bark of Eucommia ulmoides Oliv.)
3.11. Ligustri Lucidi Frustus (Fruit of Ligustrum lucidum W.T. Aiton)
3.12. Morindae Officinalis Radix (Root of Morinda officinalis F.C. How)
3.13. Podocarpium podocarpum (DC.) Yang et Huang [syn. Desmodium podocarpum DC.]
3.14. Psoraleae Fructus (Fruit of Psoralea corylifolia L. [syn. Cullen corylifolium (L.) Medik.])
3.15. Puerariae lobatae Radix [Root of Pueraria lobate (Willd.) Ohwi]
3.16. Rehmanniae Radix [Root of Rehmannia glutinosa (Gaertn.) DC.]
3.17. Salviae miltiorrhizae Radix et Rhizoma (Root and rhizome of Salvia miltiorrhiza Bunge)
3.18. Sambuci Caulis (Stem of Sambucus williamsii Hance)
3.19. Sophorae Fructus (Fruit of Sophora japonica L. [syn. Styphnolobium japonicum (L.) Schott.])
3.20. Visci Herba [Twig of Viscum coloratum (Kom.) Nakai]
3.21. Yams (Dioscorea spp.)
3.22. Miscellaneous Compounds
4. Concluding Remarks
Acknowledgments
Author Contributions
Conflicts of Interest
References
- International Osteoporosis Foundation: Facts and Statistics about Osteoporosis and Its Impact. Available online: http://www.iofbonehealth.org/facts-and-statistics.html (accessed on 14 December 2015).
- Korpi-Steiner, N.; Milhorn, D.; Hammett-Stabler, C. Osteoporosis in men. Clin. Biochem. 2014, 47, 950–959. [Google Scholar] [CrossRef] [PubMed]
- National Osteoporosis Foundation Prevalence Report. Available online: http://www.nof.org/advocacy/resources/prevalencereport (accessed on 4 December 2015).
- Management of osteoporosis in postmenopausal women: 2010 position statement of the North American menopause society. Menopause 2010, 17, 25–54.
- Tella, S.H.; Gallagher, J.C. Prevention and treatment of postmenopausal osteoporosis. J. Steroid Biochem. Mol. Biol. 2014, 142, 155–170. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Xiong, D.; Peng, Y.Q.; Sheng, Z.F.; Wu, X.Y.; Wu, X.P.; Wu, F.; Yuan, L.Q.; Liao, E.Y. Epidemiology and management of osteoporosis in the People’s Republic of China: Current perspectives. Clin. Interv. Aging 2015, 10, 1017–1033. [Google Scholar] [PubMed]
- Sandhu, S.K.; Hampson, G. The pathogenesis, diagnosis, investigation and management of osteoporosis. J. Clin. Pathol. 2011, 64, 1042–1050. [Google Scholar] [CrossRef] [PubMed]
- Sweet, M.G.; Sweet, J.M.; Jeremiah, M.P.; Salazka, S.S. Diagnosis and treatment of osteoporosis. Am. Fam. Phys. 2009, 79, 193–200. [Google Scholar]
- Rachner, T.D.; Khosla, S.; Hofbauer, L.C. Osteoporosis: Now and the future. Lancet 2011, 377, 1276–1287. [Google Scholar] [CrossRef]
- Hollick, R.J.; Reid, D.M. Role of bisphosphonates in the management of postmenopausal osteoporosis: An update on recent safety anxieties. Menopause Int. 2011, 17, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Park-Wyllie, L.Y.; Mamdani, M.M.; Juurlink, D.N.; Hawker, G.A.; Gunraj, N.; Austin, P.C.; Whelan, D.B.; Weiler, P.J.; Laupacis, A. Bisphosphonate use and the risk of subtrochanteric or femoral shaft fracture in older women. JAMA 2011, 305, 783–789. [Google Scholar] [CrossRef] [PubMed]
- Lewiecki, E.M. Safety of long-term bisphosphonate therapy of the management of osteoporosis. Drugs 2011, 71, 791–814. [Google Scholar] [CrossRef] [PubMed]
- Watts, N.B.; Diab, D.L. Long-term use of bisphosphonates in osteoporosis. J. Clin. Endocrinol. Metab. 2010, 95, 1555–1565. [Google Scholar] [CrossRef] [PubMed]
- Kos, M. Bisphosphonates promote jaw osteonecrosis through facilitating bacterial colonization. Med. Hypotheses 2011, 77, 214–215. [Google Scholar] [CrossRef] [PubMed]
- Knopp-Sihota, J.A.; Cummings, G.G.; Homik, J.; Voaklander, D. The association between serious upper gastrointestinal bleeding and incident bisphosphonate use: A population-based nested cohort study. BMC Geriatr. 2013, 13, 36. [Google Scholar] [CrossRef] [PubMed]
- Honig, S. Osteoporosis, new treatment and updates. Bull. NYU Hosp. Jt. Dis. 2010, 68, 166–170. [Google Scholar] [PubMed]
- Silva, B.C.; Bilezikian, J.P. New approaches to the treatment of osteoporosis. Annu. Rev. Med. 2011, 62, 307–322. [Google Scholar] [CrossRef] [PubMed]
- Bhutani, G.; Gupta, M.C. Emerging therapies for the treatment of osteoporosis. J. Midlife Health 2013, 4, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Chapurlat, R.D. Odanacatib for the treatment of postmenopausal osteoporosis. Expert Opin. Pharmacother. 2014, 15, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Ng, K.W.; Martin, T.J. New therapeutics for osteoporosis. Curr. Opin. Pharmacol. 2014, 16, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Bowring, C.E.; Francis, R.M. National osteoporosis society’s position statement on hormone replacement therapy in the prevention and treatment of osteoporosis. Menopause Int. 2011, 17, 63–65. [Google Scholar] [CrossRef] [PubMed]
- Spangler, M.; Phillips, B.B.; Ross, M.B.; Moores, K.G. Calcium supplementation in postmenopausal women to reduce the risk of osteoporotic fractures. Am. J. Health Syst. Pharm. 2011, 68, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Quesada-Gomez, J.M.; Blanch-Rubio, J.; Diaz-Curiel, M.; Diez-Perez, A. Calcium citrate and vitamin D in the treatment of osteoporosis. Clin. Drug Investig. 2011, 31, 285–298. [Google Scholar] [CrossRef] [PubMed]
- Al-Anazi, A.F.; Qureshi, V.F.; Javaid, K.; Qureshi, S. Preventive effects of phytoestrogens against postmenopausal osteoporosis as compared to the available therapeutic choices: An overview. J. Nat. Sci. Biol. Med. 2011, 2, 154–163. [Google Scholar] [PubMed]
- Taku, K.; Melby, M.K.; Nishi, N.; Omori, T.; Kurzer, M.S. Soy isoflavones for osteoporosis: An evidence-based approach. Maturitas 2011, 70, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Wei, P.; Liu, M.; Chen, Y.; Chen, D.C. Systematic review of soy isoflavone supplements on osteoporosis in women. Asian Pac. J. Trop. Med. 2012, 243–248. [Google Scholar] [CrossRef]
- Lagari, V.S.; Levis, S. Phytoestrogens in the prevention of postmenopausal bone loss. J. Clin. Densitom. 2013, 16, 445–449. [Google Scholar] [CrossRef] [PubMed]
- Lagari, V.S.; Levis, S. Phytoestrogens for menopausal bone loss and climacteric symptoms. J. Steroid Biochem. Mol. Biol. 2014, 139, 294–301. [Google Scholar] [CrossRef] [PubMed]
- Bedell, S.; Nachtigall, M.; Naftolin, F. The pros and cons of plant estrogens for menopause. J. Steroid Biochem. Mol. Biol. 2014, 139, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Poluzzi, E.; Piccinni, C.; Raschi, E.; Rampa, A.; Recanatini, M.; DePonti, F. Phytoestrogens in postmenopause: The state of the art from a chemical, pharmacological and regulatory perspective. Curr. Med. Chem. 2014, 21, 417–436. [Google Scholar] [CrossRef] [PubMed]
- Putnam, S.E.; Scutt, A.M.; Bicknell, K.; Priestley, C.M.; Williamson, E.M. Natural products as alternative treatments for metabolic bone disorders and for maintenance of bone health. Phytother. Res. 2007, 21, 99–112. [Google Scholar] [CrossRef] [PubMed]
- Shirwaikar, A.; Khan, S.; Kamariya, Y.H.; Patel, B.D.; Gajera, F.P. Medicinal plants for the management of post menopausal osteoporosis: A review. Open Bone J. 2010, 2, 1–13. [Google Scholar] [CrossRef]
- Wang, Z.Q.; Li, J.L.; Sun, Y.L.; Yao, M.; Gao, J.; Yang, Z.; Shi, Q.; Cui, X.J.; Wang, Y.J. Chinese herbal medicine for osteoporosis: A systematic review of randomized controlled trials. Evid. Based Complement. Altern. Med. 2013, 356260. [Google Scholar] [CrossRef] [PubMed]
- Leung, P.C.; Siu, W.S. Herbal treatment for osteoporosis: A current reivew. J. Tradit. Complement. Med. 2013, 3, 82–87. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Li, Q.; Liu, R.; Niu, Y.; Pan, Y.; Zhai, Y.; Mei, Q. Medicinal herbs in the prevention and treatment of osteoporosis. Am. J. Chin. Med. 2014, 42, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Mukwaya, E.; Xu, F.; Wong, M.S.; Zhang, Y. Chinese herbal medicine for bone health. Pharm. Biol. 2014, 52, 1223–1228. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, J.P.; Xia, Y. Chinese herbal medicines for treating osteoporosis (review). Cochrane Database Syst. Rev. 2014, CD005467. [Google Scholar]
- Rufus, P.; Mohamed, N.; Shuid, A.N. Beneficial effects of traditional Chinese medicine on the treatment of osteoporosis on ovariectomised rat models. Curr. Drug Targets 2015, 16, 1689–1693. [Google Scholar] [CrossRef]
- Zhao, X.; Feng, Y.; Peng, Y. Prevention and treatment of osteoporosis with Chinese herbal medicines. Chin. Herb. Med. 2010, 4, 265–270. [Google Scholar]
- Mukudai, Y.; Kondo, S.; Koyama, T.; Li, C.; Banka, S.; Kogure, A.; Yazawa, K.; Shintani, S. Potential antiosteoporotic effects of herbal extracts on osteoclasts, osteoblasts and chondrocytes in vitro. BMC Complement. Altern. Med. 2014, 14, 29. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Chin, A.; Zhang, L.; Lu, J.; Wong, R.W. The role of traditional Chinese medicines in osteogenesis and angiogenisis. Phytother. Res. 2014, 28, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Jia, M.; Nie, Y.; Cao, D.P.; Xue, Y.Y.; Wang, J.S.; Zhao, L.; Rahman, K.; Zhang, Q.Y.; Qin, L.P. Potential antiosteoporotic agents from plants: A comprehensive review. Evid. Based Complement. Altern. Med. 2012, 364604. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Vasikaran, S. Current recommendations for laboratory testing and use of bone turnover markers in management of osteoporosis. Ann. Lab. Med. 2012, 32, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Seeman, E.; Delmas, P.D. Bone quality—The material and structural basis of bone strength and fragility. N. Engl. J. Med. 2006, 354, 2250–2261. [Google Scholar] [CrossRef] [PubMed]
- Kanis, J.A.; McCloskey, E.V.; Johansson, H.; Oden, A.; Melton, L.J.; Khaltaev, N. A reference standard for the description of osteoporosis. Bone 2008, 42, 467–475. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Zhou, X.; Fujita, H.; Onozuka, M.; Kubo, K. Age-related changes in trabecular and cortical bone microstructure. Int. J. Endocrinol. 2013, ID213234. [Google Scholar] [CrossRef] [PubMed]
- Wheater, G.; Elshahaly, M.; Tuck, S.P.; Datta, H.K.; van Laar, J.M. The clinical utility of bone marker measurements in osteoporosis. J. Transl. Med. 2013, 11, 201. [Google Scholar] [CrossRef] [PubMed]
- Hlaing, T.T.; Compston, J.E. Biochemical markers of bone turnover-uses and limitations. Ann. Clin. Biochem. 2014, 51 Pt 2, 189–202. [Google Scholar] [CrossRef] [PubMed]
- Vasikaran, S.; Eastell, R.; Bruyere, O.; Foldes, A.J.; Garnero, P.; Griesmacher, A.; McClung, M.; Morris, H.A.; Silverman, S.; Trenti, T.; et al. Markers of bone turnover for the prediction of fracture risk and monitoring of osteoporosis treatment: A need for international reference standards. Ostoporos. Int. 2011, 22, 391–420. [Google Scholar] [CrossRef] [PubMed]
- Khashayar, P.; Meybodi, H.A.; Amoabediny, G.; Larijani, B. Biochemical markers of bone turnover and their role in osteoporosis diagnosis: A narrative review. Recent Pat. Endocr. Metab. Immune Drug Discov. 2015, 9, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Garnero, P. New development in biological markers of bone metabolism in osteoporosis. Bone 2014, 66, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Golub, E.E.; Boesze-Battaglia, K. The role of alkaline phosphatase in mineralization. Curr. Opin. Orthop. 2007, 18, 444–448. [Google Scholar] [CrossRef]
- Kress, B.C. Bone alkaline phosphatase: Methods of quantitation and clinical utility. J. Clin. Ligand Assay 1998, 21, 139–148. [Google Scholar]
- Halleen, J.; Tiitinen, S.; Ylipahkala, H.; Fagerlund, K.M.; Vaananen, K. Tartrate-resistant acid phosphatase 5b (TRACP 5b) as a marker of bone resorption. Clin. Lab. 2006, 52, 499–509. [Google Scholar] [PubMed]
- Munoz-Torres, M.; Reyes-Garcia, R.; Mezquita-Raya, P.; Fernandez-Garcia, D.; Alonso, G.; de Dios Luna, J.; Ruiz-Requena, M.E.; Escobar-Jimenez, F. Serum cathepsin K as a marker of bone metabolism in postmenopausal women treated with alendronate. Maturitas 2009, 64, 188–192. [Google Scholar] [CrossRef] [PubMed]
- Karsenty, G.; Ferron, M. The contribution of bone to whole-organism physiology. Nature 2012, 481, 314–320. [Google Scholar] [CrossRef] [PubMed]
- Jharna, S.; Nupur, S.; Purnima, D.S.; Angoorbala, B. Assessment of bone loss in postmenopausal women by evaluation of urinary hydroxyproline and serum status of osteocalcin. Int. Res. J. Biol. Sci. 2013, 2, 11–14. [Google Scholar]
- Delmas, P.D.; Schlemmer, A.; Gineyts, E.; Riis, B.; Christiansen, C. Urinary excretion of pyridinoline crosslinks correlates with bone turnover measured on iliac crest biopsy in patients with vertebral osteoporosis. J. Bone Miner. Res. 1991, 6, 639–644. [Google Scholar] [CrossRef] [PubMed]
- Vasikaran, S.; Cooper, C.; Eastell, R.; Griesmacher, A.; Morris, H.A.; Trenti, T.; Kanis, J.A. International Osteoporosis Foundation and International Federation of Clinical Chemistry and Laboratory Medicine position on bone marker standards in osteoporosis. Clin. Chem. Lab Med. 2011, 49, 1271–1274. [Google Scholar] [CrossRef] [PubMed]
- Fodor, D.; Bondor, C.; Albu, A.; Simon, S.P.; Craciun, A.; Muntean, L. The value of osteopontin in the assessment of bone mineral density status in postmenopausal women. J. Invertig. Med. 2013, 61, 15–21. [Google Scholar]
- Jabbar, S.; Drury, J.; Fordham, J.N.; Datta, H.K.; Francis, R.M.; Tuck, S.P. Ostoprotegerin, RANKL and bone turnover in postmenopausal osteoporosis. J. Clin. Pathol. 2011, 64, 354–357. [Google Scholar] [CrossRef] [PubMed]
- Khosla, S. Minireview: The OPG/RANKL/RANK system. Endocrinology 2001, 142, 5050–5055. [Google Scholar] [CrossRef] [PubMed]
- Bauer, D.; Krege, J.; Lane, N.; Leary, E.; Libanati, C.; Miller, P.; Myers, G.; Silveman, S.; Vesper, H.W.; Lee, D.; et al. National bone health alliance bone turnover marker project: Current practices and the need for us harmonization, standardization, and common reference ranges. Osteoporos. Int. 2012, 23, 2425–2433. [Google Scholar] [CrossRef] [PubMed]
- Kostenuik, P.J. Osteoprotegerin and RANKL regulate bone resorption, density, geometry and strength. Curr. Opin. Pharmacol. 2005, 5, 618–625. [Google Scholar] [CrossRef] [PubMed]
- Boyce, B.F.; Xing, L. Biology of RANK, RANKL, and osteoprotegerin. Arthritis Res. Ther. 2007, 9 (Suupl. 1), S1. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Bae, Y.C.; Park, R.W.; Choi, S.W.; Cho, S.H.; Choi, Y.S.; Lee, W.J. Bone-protecting effect of safflower seeds in ovariectomized rats. Calcif. Tissue Int. 2002, 71, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.R.; Kim, S.M.; Lee, J.I.; Chon, S.K.; Choi, S.J.; Choi, I.H.; Kim, N.S. Effects of safflower seed oil in osteoporosis induced-ovariectomized rats. Am. J. Chin. Med. 2006, 34, 601–612. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Choi, C.W.; Kim, J.J.; Ganapathi, A.; Udayakumar, R.; Kim, S.C. Determination of mineral content in methanolic safflower (Carthamus tinctorius L.) seed extract and its effect on osteoblast markers. Int. J. Mol. Sci. 2009, 10, 292–305. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.O.; Park, Y.S.; Lee, J.H.; Seo, J.B.; Koo, K.I.; Jeong, S.C.; Jin, S.D.; Lee, Y.H.; Eom, H.S.; Yun, I. Effect of extracts from safflower seeds on osteoblast differentiation and intracellular calcium ion concentration in MC3T3-E1 cells. Nat. Prod. Res. 2007, 21, 787–797. [Google Scholar] [CrossRef] [PubMed]
- Yuk, T.H.; Kang, J.H.; Lee, S.R.; Yuk, S.W.; Lee, K.G.; song, B.Y.; Kim, C.H.; Kim, D.W.; Dong, I.K.; Lee, T.K.; et al. Inhibitory effects of Carthamus tinctorius L. seed extracts on bone resoprtion medicated by tyrosine kinase, COX-2 (clyclooxygenase) and PG (prostaglandin) E2. Am. J. Chin. Med. 2002, 30, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Moon, H.J.; Kim, E.K.; Nam, Y.S.; Kim, J.H.; Ko, W.K.; Lee, J.M.; Lee, C.H.; Jang, J.B.; Lee, K.S.; Kwon, I.K. Safflower seed extract inhibits osteoclast differentiation by suppression of the p38 mitogen-activated protein kinase and IκB kinase activity. Phytother. Res. 2012, 26, 1648–1655. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.W.; Lee, W.J.; Park, R.W. Use of polyphenol compounds obtained from the seeds of Carthamus tinctorius L. Repub. Korean Kongkae Taeho Kongbo KR 1020010034963 A 2020060809, 7 May 2001. [Google Scholar]
- Li, J.X.; Kadota, S.; Li, H.Y.; Miyahara, T.; Wu, Y.W.; Seto, H.; Namba, T. Effects of Cimicifugae Rhizoma on serum calcium and phosphate levels in low calcium dietary rats and on bone mineral density in ovariectomized rats. Phytomedicine 1996, 3, 379–385. [Google Scholar] [CrossRef]
- Li, J.X.; Liu, J.; He, C.C.; Yu, Z.Y.; Du, Y.; Kadota, S.; Seto, H. Triterpenoids from Cimicifugae Rhizoma, a novel class of inhibitors on bone resorption and ovariectomy-induced bone loss. Maturitas 2007, 58, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Ahn, B.S.; Yang, M.; Jang, H.; Lee, H.J.; Moon, C.; Kim, J.C.; Jung, U.; Jo, S.K.; Jang, J.S. Evaulation of the antiosteoporotic potential of Cimicifuga heracleifolia in female mice. Phytother. Res. 2012, 26, 663–668. [Google Scholar] [CrossRef] [PubMed]
- Viereck, V.; Grundker, C.; Friess, S.C.; Frosch, K.H.; Raddatz, D.; Schoppet, M.; Nisslein, T.; Emons, G.; Hofbauer, L.C. Isopropanolic extract of black cohosh stimulates osteoprotegerin production by human osteoblasts. J. Bone Miner. Res. 2005, 20, 2036–2043. [Google Scholar] [CrossRef] [PubMed]
- Cui, G.; Leng, H.; Wang, K.; Wang, J.; Zhu, S.; Jia, J.; Chen, X.; Zhang, W.; Qin, L.; Bai, W. Effects of remifemin treatment on bone integrity and remodeling in rats with ovariectomy-induced osteoporosis. PLoS ONE 2013, 8, e82815. [Google Scholar] [CrossRef] [PubMed]
- Seidlova-Wuttke, D.; Stecher, G.; Kammann, M.; Haunschild, J.; Eder, N.; Stahnke, V.; Wessels, J.; Wuttke, W. Osteoprotective effects of Cimicifuga racemosa and its triterpene saponins are responsible for reduction of bone marrow fat. Phytomedicine 2012, 19, 855–860. [Google Scholar] [CrossRef] [PubMed]
- Qiu, S.X.; Dan, C.; Ding, L.S.; Peng, S.; Chen, S.N.; Farnsworth, N.R.; Nolta, J.; Gross, M.L.; Zhou, P. A triterpene glycoside from black cohosh that inhibits osteoclastogeneisis by modulating RANKL and TNFα signaling pathways. Chem. Biol. 2007, 14, 860–869. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.M. Deoxyactein stimulates osteoblast function and inhibits bone-resorbing mediators in MC3T3-E1 cells. J. Appl. Toxicol. 2013, 33, 190–195. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, K.; Shinohara, C.; Kojima, S.; Sodeoka, M.; Tsuji, T. (2E,6R)-8-Hydroxy-2,6-dimethyl-2-octenoic acid, a novel anti-osteoporotic monoterpene, isolated from Cistanche salsa. Biosci. Biotechnol. Biochem. 1999, 63, 731–735. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.; Yu, F.; Tong, Z.; Huang, Z. Effect of Cistanches Herba aqueous extract on bone loss in ovariectomized rat. Int. J. Mol. Sci. 2011, 12, 5060–5069. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.D.; Yu, F.; Tong, Z.H.; Zhang, H.Q.; Liang, W. Cistanches Herba aqueous extract affecting serum BGP and TRAP and bone marrow Smad1 mRNA, Smad5 mRNA, TGF-β1 mRNA and TIEG1 mRNA expression levels in osteoporosis disease. Mol. Biol. Rep. 2013, 40, 757–763. [Google Scholar] [CrossRef] [PubMed]
- Li, T.M.; Huang, H.C.; Su, C.M.; Ho, T.Y.; Wu, C.M.; Chen, W.C.; Fong, Y.C.; Tang, C.H. Cistanche deserticola extract increases bone formation in osteoblasts. J. Pharm. Pharmacol. 2012, 64, 897–907. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Yang, Y.; Zhu, P.; Chen, W.; Qi, D.; Shi, X.; Zhang, C.; Yang, Z.; Li, P. Echinacoside promotes bone regeneration by increasing OPG/RANKL ratio in MC3T3-E1 cells. Fitoterapia 2012, 83, 1443–1450. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Yang, X.; Yang, Y.; Guo, C.; Zhang, C.; Yang, Z.; Li, P. Antiosteoporotic activity of echinacoside in ovariectomized rats. Phytomedicine 2013, 20, 549–557. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Li, F.; Yang, Y.; Shen, J.; Zou, R.; Zhu, P.; Zhang, C.; Yang, Z.; Li, P. Efficacy and safety of echinacoside in a rat osteopenia model. Evid. Based Complement. Altern. Med. 2013, 926928. [Google Scholar] [CrossRef] [PubMed]
- Mizuha, Y.; Yamamoto, H.; Sato, T.; Tsuji, M.; Masuda, M.; Uchida, M.; Sakai, K.; Taketani, Y.; Yasutomo, K.; Sasaki, H.; et al. Water extract of Cordyceps sinensis (Wecs) inhibits the RANKL-induced osteoclast differentiation. Biofactors 2007, 30, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Qi, W.; Yan, Y.B.; Wang, P.J.; Lei, W. The co-effect of Cordyceps sinensis and strontium on osteoporosis in ovariectomized rats. Biol. Trace Elem. Res. 2011, 141, 216–223. [Google Scholar] [CrossRef] [PubMed]
- Qi, W.; Wang, P.J.; Guo, W.J.; Yan, Y.B.; Zhang, Y.; Lei, W. The mechanism of Cordyceps sinensis and strontium in prevention of osteoporosis in rats. Biol. Trace Elem. Res. 2011, 143, 302–309. [Google Scholar] [CrossRef] [PubMed]
- Qi, W.; Yan, Y.B.; Lei, W.; Wu, Z.X.; Zhang, Y.; Liu, D.; Shi, L.; Cao, P.C.; Liu, N. Prevention of disuse osteoporosis in rats by Cordyceps sinensis extract. Ostoporos. Int. 2012, 23, 2347–2357. [Google Scholar] [CrossRef] [PubMed]
- Qi, W.; Zhang, Y.; Yan, Y.B.; Lei, W.; Wu, Z.X.; Liu, N.; Liu, S.; Shi, L.; Fan, Y. The protective effect of cordymin, a peptide purified from the medicinal mushroom Cordyceps sinensis, on diabetic osteopenia in alloxan-induced diabetic rats. Evid. Based Complement. Altern. Med. 2013, 985636. [Google Scholar] [CrossRef]
- Zhang, D.W.; Wang, Z.L.; Qi, W.; Zhao, G.Y. The effects of Cordyceps sinensis phytoestrogen on estrogen deficiency-induced osteoporosis in ovariectomized rats. BMC Complement. Altern. Med. 2014, 14, 484. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Lee, H.; Kang, K.S.; Chun, K.H.; Hwang, G.S. Cordyceps militaris mushroom and cordycepin inhibit rnakl-induced osteoclast differentiation. J. Med. Food 2015, 18, 446–452. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.W.; Wang, Z.L.; Qi, W.; Lei, W.; Zhao, G.Y. Cordycepin (3′-deoxyadenosine) down-regulates the proinflammatory cytokines in inflammation-induced osteoporosis model. Inflammation 2014, 37, 1044–1049. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Cao, Y.; Lv, Z.; Jiang, T.; Wang, L.; Li, Z. Cordycepin protected against the TNF-α-induced inhibition of osteogenic differentiation of human adipose-derived mesenchymal stem cells. Int. J. Immunopathol. Pharmacol. 2015, 28, 296–307. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.W.; Deng, H.; Qi, W.; Zhao, G.Y.; Cao, X.R. Osteoprotective effect of cordycepin on estrogen deficiency-induced osteoporosis in vitro and in vivo. Biomed. Res. Int. 2015, 423869. [Google Scholar] [CrossRef]
- Wong, R.W.K.; Rabie, A.B.M.; Hagg, E.U.O. The effect of crude extract from Radix Dipsaci on bone in mice. Phytother. Res. 2007, 21, 596–598. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.G.; Zhang, R.; Li, C.; Ma, X.; Liu, L.; Wang, J.P.; Mei, Q.B. The osteoprotective effect of Radix Dipsaci extract in ovariectomized rats. J. Ethnopharmacol. 2009, 123, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; Li, C.; Pan, Y.; Li, Y.; Kong, X.; Wang, S.; Zhai, Y.; Wu, X.; Mei, Q. Treatment of Radix Dipsaci extract prevents long bone loss induced by modeled microgravity in hindlimb unloading rats. Pharm. Biol. 2015, 53, 110–116. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.B.; Li, Y.H.; Kong, X.H.; Zhang, R.; Sun, Y.; Li, Q.; Li, C.; Liu, L.; Wang, J.P.; Mei, Q.B. The beneficial effect of Radix Dipsaci total saponins on bone metabolism in vitro and in vivo and the possible mechanisms of action. Osteoporos. Int. 2012, 23, 2649–2660. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.B.; Kong, X.H.; Li, Y.H.; Fan, L.; Pan, Y.L.; Li, C.R.; Wu, X.L.; Lu, T.L.; Mei, Q.B. Radix Dipsaci total saponins stimulate MC3T3-E1 cell differentiation via the bone morphogenetic protein-2/MAPK/Smad-dependent RUNX2 pathway. Mol. Med. Rep. 2015, 11, 4468–4472. [Google Scholar] [PubMed]
- Niu, Y.; Li, Y.; Huang, H.; Kong, X.; Zhang, R.; Liu, L.; Sun, Y.; Wang, T.; Mei, Q. Asperosaponin VI, a saponin component from Dipsacus asper Wall, induces osteoblastic differentiation through bone morphogenetic protein-2/p38 and extracellular signal-regulated kinase 1/2 pathway. Phytother. Res. 2011, 25, 1700–1706. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.S.; Kim, Y.C.; Zadeh, H.; Park, Y.J.; Pi, S.H.; Shin, H.S.; You, H.K. Effects of the dichloromethane fraction of Dipsaci Radix on the osteoblastic differentiation of human alveolar bone marrow-derived mesenchymal stem cells. Biosci. Biotechnol. Biochem. 2011, 75, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Jing, L.J.; Wang, C.M.; Wei, Y.J.; Jia, X.B.; Xie, L. Screening on antiosteoporotic active parts of Dipsacus Radix based on zebrafish model. Chin. Herb. Med. (Zhongyaocai) 2014, 37, 635–640. [Google Scholar]
- Wang, C.; Jing, L.; Wei, Y.; Jia, X. Evaluation of the antiosteoporotic activity of micro-amount asperosaponins V and VI in zebrafish osteoporosis model. J. China Pharm. Univ. (Zhongguo Yaoke Daxue Xuebao) 2014, 45, 88–91. [Google Scholar]
- Lin, C.Y.; Sun, J.S.; Sheu, S.Y.; Lin, F.H.; Wang, Y.J.; Chen, L.T. The effect of Chinese medicine on bone cell activities. Am. J. Chin. Med. 2002, 30, 271–285. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.S.; Lin, C.Y.; Dong, G.C.; Sheu, S.Y.; Lin, F.H.; Chen, L.T.; Wang, Y.L. The effect of Gu-Sui-Bu (Drynaria fortunei J. Sm) on bone cell activities. Biomaterials 2002, 23, 3377–3385. [Google Scholar] [CrossRef]
- Jeong, J.C.; Kang, S.K.; Youn, C.H.; Jeong, C.W.; Kim, H.M.; Lee, Y.C.; Chang, Y.C.; Kim, C.H. Inhibition of Drynariae Rhizoma extracts on bone resorption mediated by processing of cathepsin K in cultured mouse osteoclasts. Int. Immunopharmacol. 2003, 3, 1685–1697. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.L.; Liu, K.; Wu, L.G. Interventional value of total flavonoids from Rhizoma Drynariae on cathepsin K, a potential target of osteoporosis. Chin. J. Integr. Med. 2011, 17, 556–560. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.L.; Li, C.W.; Wan, Q.Z.; Li, A.Q.; Wang, H.; Liu, K. Drynaria total flavonoids decrease cathepsin K expression in ovariectomized rats. Genet. Mol. Res. 2014, 13, 4311–4319. [Google Scholar] [CrossRef] [PubMed]
- Wong, R.W.; Rabie, A.B. Systemic effect of crude extract from rhizome of Drynaria foutunei on bone formation in mice. Phytother. Res. 2006, 20, 313–315. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Chen, H.; Zhu, M. Effects of Drynaria fortunei on bone ultramicrostructure and proline hydroxylation level of ovariectomized rats. Zhongguo Gushang 2011, 24, 48–50. [Google Scholar]
- Lee, Y.E.; Liu, H.C.; Lin, Y.L.; Liu, S.H.; Yang, R.S.; Chen, R.M. Drynaria fortunei J. Sm. improves the bone mass of ovariectomized rats through osteocalcin-involved endochondral ossification. J. Ethnopharmacol. 2014, 158 Pt A, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.L.; Wang, N.L.; Zhang, Y.; Gao, H.; Pang, W.Y.; Wong, M.S.; Zhang, G.; Qin, L.; Yao, X.S. Effects of eleven flavonoids from the ostoprotective fraction of Drynaria fortunei (Kunze) J. Sm. on osteoblastic proliferation using an osteoblast-like cell line. Chem. Pharm. Bull. 2008, 56, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhen, L.; Zhang, G.; Wong, M.S.; Qin, L.; Yao, X.S. Osteogenic effects of flavonoid aglycones from an osteoprotective fraction of Drynaria fortunei—An in vitro efficacy study. Phytomedicine 2011, 18, 868–872. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.C.; Pang, W.Y.; Wang, X.L.; Mok, S.K.; Lai, W.P.; Chow, H.K.; Leung, P.C.; Yao, X.S.; Wong, M.S. Drynaria fortunei-derived flavonoid fraction and isolated compounds exert oestrogen-like protective effects in bone. Br. J. Nutr. 2013, 110, 475–485. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Dai, K.R.; Yan, S.G.; Yan, W.Q.; Zhang, C.; Chen, D.Q.; Xu, B.; Xu, Z.W. Effects of naringin on the proliferation and osteogenic differentiation of human bone messenchymal stem cell. Eur. J. Pharmacol. 2009, 607, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Shu, X.; Liu, J.; Zhu, D.; Pang, T. Effect of Rhizoma Drynariae total flavonoids on osteogenesis in cultured bone mesenchymal stem cells. Chin. J. Pathol. Physiol. (Zhongguo Bingli Shengli Zashi) 2010, 26, 1261–1264. [Google Scholar]
- Chen, L.L.; Lei, L.H.; Ding, P.H.; Tang, Q.; Wu, Y.M. Osteogenic effect of Drynariae rhizoma extracts and naringin on MC3T3-E1 cells and an induced rat alveolar bone resoption model. Arch. Oral. Biol. 2011, 56, 1655–1662. [Google Scholar] [CrossRef] [PubMed]
- Pang, W.Y.; Wang, X.L.; Wong, K.C.; Leung, P.C.; Yao, X.S.; Wong, M.S. Total flavonoid fraction of Rhizoma Drynaria improves bone properties in ovariectomized mice and exerts estrogen-like activities in rat osteoblast-like (UMR-106) cells. J. Food Drug Anal. 2012, 20 (Suppl. 1), 265–269. [Google Scholar]
- Guo, Y.; Li, P.F.; Shu, X.C.; Deng, H.; Ma, H.L.; Sun, L. Involvement of Wnt/β-catenin signaling in the osteogenesis of bone marrow mesenchymal stem cells induced by Drynaria total flavonoids. Chin. J. Med. (Zhonghua Yixue Zazhi) 2012, 92, 2288–2291. [Google Scholar]
- Hung, T.Y.; Chen, T.L.; Liao, M.H.; Ho, W.P.; Liu, D.Z.; Chuang, W.C.; Chen, R.M. Drynaria fortunei J. Sm. promotes osteoblast maturation by inducing differentiation-related gene expression and protecting against oxidative stress-induced apoptotic insults. J. Ethnopharmacol. 2010, 131, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Yin, F.M.; Xiao, L.B.; Zhang, Y. Research progress on Drynaria fortunei naringin on inflammation and bone activity. Chin. Orthop. (Zhongguo Gushang) 2015, 28, 182–186. [Google Scholar]
- Wong, R.W.; Rabie, A.B. Effect of naringin on bone cells. J. Orthop. Res. 2006, 24, 2045–2050. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.B.; Fong, Y.C.; Tsai, H.Y.; Chen, Y.F.; Tsuzuki, M.; Tang, C.H. Naringin-induced bone morphogenetic protein-2 expression via PI3K, AKT, c-Fos/c-Jun and AP-1 pathway in osteoblasts. Eur. J. Pharmacol. 2008, 588, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Ding, P.; Tang, Q.; Chen, L. Effects of naringin on proliferation, differentiation and matrix mineralization of MC3T3-E1 cells. Chin. J. Chin. Med. (Zhongguo Zhongyao Zazhi) 2009, 34, 1712–1716. [Google Scholar]
- Pang, W.Y.; Wang, X.L.; Mok, S.K.; Lai, W.P.; Chow, H.K.; Leung, P.C.; Yao, X.S.; Wong, M.S. Naringin improves bone properties in ovariectomized mice and exerts oestrogen-like activities in rat osteoblast-like (UMR-106) cells. Br. J. Pharmacol. 2010, 159, 1693–1703. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Jiang, Y.; Wooley, P.H.; Xu, Z.; Yang, S.Y. Naringin promotes osteoblast differentiation and effectively reverses ovariectomy-associated osteoporosis. J. Orthop. Res. 2013, 18, 478–485. [Google Scholar]
- Zhai, Y.K.; Niu, Y.B.; Pan, Y.L.; Li, C.R.; Wu, X.L.; Mei, Q.B. Effects of naringin on proliferation, differentiation and maturation of rat calvarial osteoblasts in vitro. Chin. J. Chin. Med. (Zhongguo Zhongyao Zazhi) 2013, 38, 105–111. [Google Scholar]
- Liu, M.; Li, Y.; Yang, S.T. Effects of naringin on the proliferation and osteogenic differentiation of human amniotic fluid-derived stem cells. J. Tissue Eng. Regen. Med. 2014. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Cheng, W.; Qin, Z.; Yu, H.; Yu, Z.; Zhong, M.; Sun, K.; Zhang, W. Effects of naringin on proliferation and osteogenic differentiation of human periodontal ligament stem cells in vitro and in vivo. Stem Cell Int. 2015, 758706. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Li, J.; Fan, Q. Naringin promotes differentiation of bone marrow stem cells into osteoblasts by upregulating the expression levels of microrRNA-20a and downregulating the expression levels of PPARγ. Mol. Med. Rep. 2015, 12, 4759–4765. [Google Scholar] [PubMed]
- Cao, X.; Lin, W.; Liang, C.; Zhang, D.; Yang, F.; Zhang, Y.; Zhang, X.; Feng, J.Q.; Chen, C. Naringin rescued the TNF-α-induced inhibition of osteogenesis of bone marrow-derived mesenchymal stem cells by depressing the activation of NF-κB signaling pathway. Immunol. Res. 2015, 62, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Zhao, X.; Wu, T.; Zhou, Z.; Gao, Y.; Wang, X.; Zhang, C.Q. Inhibiting wear particles-induced osteolysis with naringin. Int. Orthop. 2013, 37, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Li, F.B.; Sun, X.L.; Ma, J.X.; Zhang, Y.; Zhao, B.; Li, Y.L.; Ma, X.L. Effect of naringin on osteoclast differentiation. Chin. J. Chin. Med. (Zhongguo Zhongyao Zazhi) 2015, 40, 308–312. [Google Scholar]
- Ang, E.S.; Yang, X.; Chen, H.; Liu, Q.; Zheng, M.H.; Xu, J. Naringin abrogates osteoclastogenesis and bone resorption via the inhibition of RANKL-induced NF-κB and ERK activation. FEBS Lett. 2011, 585, 2755–2762. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Sun, X.; Ma, J.; Ma, X.; Zhao, B.; Zhang, Y.; Tian, P.; Li, Y.; Han, Z. Naringin prevents ovariectomy-induced osteoporosis and promotes osteoclasts apoptosis through the mitochondria-medicated apoptosis pathway. Biochem. Biophys. Res. Commun. 2014, 452, 629–635. [Google Scholar] [CrossRef] [PubMed]
- Wei, M.; Yang, Z.; Li, P.; Zhang, Y.; See, W.C. Anti-osteoporosis activity of naringin in the retinoic acid-induced osteoporosis model. Am. J. Chin. Med. 2007, 35, 663–667. [Google Scholar] [CrossRef] [PubMed]
- Mandadi, K.; Ramirez, M.; Jayaprakasha, G.K.; Faraji, B.; Lihono, M.; Deyhim, F.; Patil, B.S. Citrus bioactive compounds improve bone qualtiy and plasma antioxidant activity in orchidectomized rats. Phytomedicine 2009, 16, 513–520. [Google Scholar] [CrossRef] [PubMed]
- Habauzit, V.; Sacco, S.M.; Gil-Izquierdo, A.; Trzeciakiewicz, A.; Morand, C.; Barron, D.; Pinaud, S.; Offord, E.; Horcajada, M.N. Differential effects of two citrus flavanones on bone quality in senescent male rats in relation to their bioavailability and metabolism. Bone 2011, 49, 1108–1116. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Zhang, P.; Zhang, C.; Zhu, Z. Promotion of bone formation by naringin in a titanium particle-induced diabetic murine calvarial osteolysis model. J. Orthop. Res. 2010, 28, 451–456. [Google Scholar] [CrossRef] [PubMed]
- Wong, R.W.; Rabie, A.B. Effect of naringin collagen graft on bone formation. Biomaterials 2006, 27, 1824–1831. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.Y.; Lin, K.C.; Chen, Y.S.; Yao, C.H. A novel porous gelatin composite containing naringin for bone repair. Evid. Based Complement. Altern. Med. 2013, 283941. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.; Wang, L.; Watts, D.C.; Qiu, H.; You, T.; Deng, F.; Wu, X. Controlled-release naringin nanoscaffold for osteoporotic bone healing. Dent. Mater. 2014, 30, 1263–1273. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Xu, Z.; Wooley, P.H.; Zhang, J.; Yang, S.Y. Therapeutic potentials of naringin on polymethylmethacrylate induced osteoclastogenesis and osteolysis, in vitro and in vivo assessments. Drug Des. Devel. Ther. 2014, 8, 1–11. [Google Scholar] [PubMed]
- Li, N.H.; Xu, Z.W. Naringin reduced polymethylmethacrylate-induced osteolysis in the mouse air sacs model. Chin. Orthop. (Zhongguo Gushang) 2015, 28, 345–349. [Google Scholar]
- Kang, S.N.; Lee, J.S.; Park, J.H.; Cho, J.H.; Park, J.H.; Cho, K.K.; Lee, O.H.; Kim, I.S. In vitro anti-osteoporosis properties of diverse Korean Drynariae Rhizoma phenolic extracts. Nutrients 2014, 6, 1737–1751. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.Y.; Lin, Y.H.; Wu, J.C.; Chen, Y.C.; Yang, S.H.; Chen, J.L.; Chen, T.J. Prescription patterns of Chinese herbal products for menopausal syndrome: Analysis of a nationwide preseciption database. J. Ethnopharmacol. 2011, 137, 1261–1266. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.K.; Ha, N.R.; Yang, H.; Sung, S.H.; Kim, Y.C. Simulatory constituents of Eclipta prostrata on mouse osteoblast differentiation. Phytother. Res. 2009, 23, 129–131. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.H.; Wu, Y.B.; Lin, S.; Zeng, J.W.; Zeng, P.Y.; Wu, J.Z. Effects of volatile components and ethanolic extract from Eclipta prostrata on proliferation and differentiation of primary osteoblasts. Molecules 2010, 15, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.G.; Bai, D.; Liu, M.J.; Li, Y.; Pan, J.H.; Liu, H.; Wang, W.L.; Xiang, L.H.; Xiao, G.G.; Ju, D.H. Therapeutic effect of aqueous extract from Ecliptae Herba on bone metabolism of ovariectomized rats. Menopause 2013, 20, 232–240. [Google Scholar] [CrossRef] [PubMed]
- Joh, E.H.; Gu, W.; Kim, D.H. Echinocystic acid ameliorates lung inflammation in mice and alveolar macrophages by inhibiting the binding of LPS to TLR4 in NF-κB and MAPK pathways. Biochem. Pharmacol. 2012, 84, 331–240. [Google Scholar] [CrossRef] [PubMed]
- Ryu, S.; Shin, J.S.; Jung, J.Y.; Cho, Y.W.; Kim, S.J.; Jang, D.S.; Lee, K.T. Echinocystic acid isolated from Eclipta prostrata suppresses lipopolysaccharide-induced iNOS, TNF-α, and IL-6 expressions via NF-κB inactivation in RAW 264.7 macrophages. Planta Med. 2013, 79, 1031–1037. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wang, Q.; Xiao, S.L.; Yu, F.; Ye, M.; Zheng, Y.X.; Zhao, C.K.; Sun, D.A.; Zhang, L.H.; Zhou, D.M. Elucidation of the pharmacophore of echinocystic acid, a new lead for blocking HCV entry. Eur. J. Med. Chem. 2013, 64, 160–168. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.T.; Kang, W.B.; Zhao, J.N.; Liu, G.; Zhao, M.G. Osteoprotective effect of echinocystic acid, a triterpone component from Eclipta prostrata, in ovariectomy-induced osteoporotic rats. PLoS ONE 2015, 10, e0136572. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.Q.; Zhan, L.B.; Liu, T.; Cheng, M.C.; Liu, X.Y.; Xiao, H.B. Inhibitory effect of Ecliptae Herba extract and its component wedelolactone on pre-osteoclastic proliferation and differentiation. J. Ethnopharmacol. 2014, 157, 206–211. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Y.K.; Guo, X.; Pan, Y.L.; Niu, Y.B.; Li, C.R.; Wu, X.L.; Mei, Q.B. A systematic review of the efficacy and pharmacological profile of Herba Epimedii in osteoporosis therapy. Pharmazie 2013, 68, 713–722. [Google Scholar] [PubMed]
- Wu, H.; Lien, E.J.; Lien, L.L. Chemical and pharmacological investigations of Epimedium species: A survey. Prog. Drug Res. 2003, 60, 1–57. [Google Scholar] [PubMed]
- Zhang, G.; Qin, L.; Hung, W.Y.; Shi, Y.Y.; Leung, P.C.; Yeung, H.Y.; Leung, K.S. Flavonoids derived from herbal Epimedium brevicornum Maxim. Prevent OVX-induced osteoporosis in rats independent of its enhancement in intestinal calcium absorption. Bone 2006, 38, 818–825. [Google Scholar] [CrossRef] [PubMed]
- Songlin, P.; Ge, Z.; Yixin, H.; Xinluan, W.; Pingchung, L.; Kwoksui, L.; Ling, Q. Epimedium-derived flavonoids promote osteoblastogenesis and suppress adipogenesis in bone marrow stromal cells while exerting an anabolic effect on osteoporotic bone. Bone 2009, 45, 534–544. [Google Scholar] [PubMed]
- Xu, Y.X.; Wu, C.L.; Wu, Y.; Tong, P.J.; Jin, H.T.; Yu, N.Z.; Xiao, L.W. Epimedium-derived flavonoids modulate the balance between osteogenic differentiation and adipogenic differentiation in bone marrow stromal cells of ovariectomized rats via Wnt/β-catenin signal pathway activation. Chin. J. Integr. Med. 2012, 18, 909–917. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Qin, L.; Shi, Y. Epimedium-derived phytoestrogen flavonoids exert beneficial effect on preventing bone loss in late postmenopausal women: A 24-month randomized, double-blind and placebo-controlled trial. J. Bone Miner. Res. 2007, 22, 1072–1079. [Google Scholar] [CrossRef] [PubMed]
- Ming, L.G.; Chen, K.M.; Xian, C.J. Functions and action mechanisms of flavonoids genistein and icariin in regulating bone remodeling. J. Cell. Physiol. 2013, 228, 513–521. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Li, Q.; Mei, Q.; Lu, T. Pharmacological effects and pharmacokinetic properties of icariin, the major bioactive component in Herba Epimedii. Life Sci. 2015, 126, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Yu, Z.; Qu, H.; Li, M. Comparative effects of hispidulin, genistein, and icariin with estrogen on bone tissue in ovariectomized rats. Cell. Biochem. Biophys. 2014, 70, 485–490. [Google Scholar] [CrossRef] [PubMed]
- Nian, H.; Ma, M.H.; Nian, S.S.; Xu, L.L. Antiosteoporotic activity of icariin in ovariectomized rats. Phytomedicine 2009, 16, 320–326. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Zhong, C.; He, R.X.; Chen, L.F. Icariin associated with exercise therapy is an effective treatment for postmenopausal osteoporosis. Chin. Med. J. (Engl. Ed.) 2012, 125, 1784–1789. [Google Scholar]
- Mok, S.K.; Chen, W.F.; Lai, W.P.; Leung, P.C.; Wang, X.L.; Yao, X.S.; Wong, M.S. Icariin protects against bone loss induced by oestrogen deficiency and activates oestrogen receptor-dependent osteoblastic functions in UMR 106 cells. Br. J. Pharmacol. 2010, 159, 939–949. [Google Scholar] [CrossRef] [PubMed]
- Bian, Q.; Huang, J.H.; Liu, S.F.; Ning, Y.; Yang, A.; Zhao, Y.J.; Shen, Z.Y.; Wang, Y.J. Different molecular targets of icariin on bMSCs in CORT and OVX-rats. Front. Biosci. (Elite Ed.) 2012, 4, 1224–1236. [Google Scholar] [CrossRef] [PubMed]
- Li, G.W.; Xu, Z.; Chang, S.X.; Nian, H.; Wang, X.Y.; Qin, L.D. Icariin prevents ovariectomy-induced bone loss and lowers marrow adipogenesis. Menopause 2014, 21, 1007–1016. [Google Scholar] [CrossRef] [PubMed]
- Feng, R.; Feng, L.; Yuan, Z.; Wang, D.; Wang, F.; Tan, B.; Han, S.; Li, T.; Li, D.; Han, Y. Icariin protects against glucocorticoid-induced osteoporosis in vivo and prevents glucocorticoid-induced osteocyte apoptosis in vitro. Cell. Biochem. Biophys. 2013, 67, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Song, J.; Shao, J. Icariin attenuates glucocorticoid-induced bone deteriorations, hypocalcemia and hypercalciuria in mice. Int. J. Clin. Exp. Med. 2015, 8, 7306–7314. [Google Scholar] [PubMed]
- Zheng, D.; Peng, S.; Yang, S.H.; Shao, Z.W.; Yang, C.; Feng, Y.; Wu, W.; Zhen, W.X. The beneficial effect of icariin on bone is diminished in osteoprotegerin-deficient mice. Bone 2012, 51, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Li, X.F.; Xu, H.; Zhao, Y.J.; Tang, D.Z.; Xu, G.H.; Holz, J.; Wang, J.; Cheng, S.D.; Shi, Q.; Wang, Y.J. Icariin augments bone formation and reverses the phenotypes of osteoprotegerin-deficient mice through the activation of Wnt/β-catenin-BMP signaling. Evid. Based Complement. Altern. Med. 2013, 652317. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Y.K.; Guo, X.Y.; Ge, B.F.; Zhen, P.; Ma, X.N.; Zhou, J.; Ma, H.P.; Xian, C.J.; Chen, K.M. Icariin stimulates the osteogenic differentiation of rat bone marrow stromal cells via activating the PI3K-AKT-eNOS-NO-cGMP-PKG. Bone 2014, 66, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.; Liu, M.; Sun, L.; Rui, F. Icariin recovers the osteogenic differentiation and bone formation of bone marrow stromal cells from a rat model of estrogen deficiency-induced osteoporosis. Mol. Med. Rep. 2015, 12, 382–388. [Google Scholar] [CrossRef] [PubMed]
- Meng, F.H.; Li, Y.B.; Xiong, Z.L.; Jiang, Z.M.; Li, F.M. Osteoblastic proliferative activity of Epimedium brevicornuim Maxim. Phytomedicine 2005, 12, 189–193. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.J.; Cao, L.G.; Wu, T.; Wang, D.X.; Jin, D.; Jiang, S.; Zhang, Z.Y.; Bi, L.; Pei, G.X. The dose-effect of icariin on the proliferation and osteogenic differentiation of human bone mesenchymal stem cells. Molecules 2011, 16, 10123–10133. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.X.; Chen, Z.Q.; Liu, Z.L.; Ma, Q.J.; Dang, G.T. Icariin stimulates proliferation and differentiation of human osteoblasts by increasing production of bone morphogenetic protein 2. Chin. Med. J. (Engl. Ed.) 2007, 120, 204–210. [Google Scholar]
- Hsieh, T.P.; Sheu, S.Y.; Sun, J.S.; Chen, M.H.; Liu, M.H. Icariin isolated from Epimedium pubescens regulates osteoblasts anabolism through BMP-2, SMAD4, and Cbfa1 expression. Phytomedicine 2010, 17, 414–423. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Ohba, S.; Komiyama, Y.; Shinkai, M.; Chung, U.I.; Nagamune, T. Icariin: A potential osteoinductive compound for bone tissue engineering. Tissue Eng. (Part A) 2010, 16, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.P.; Ming, L.G.; Ge, B.F.; Zhai, Y.K.; Song, P.; Xian, C.J.; Chen, K.M. Icariin is more potent than genistein in promoting osteoblast differentiation and mineralization in vitro. J. Cell Biochem. 2011, 112, 916–923. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Ke, Y.; Zhang, Y.; Zhang, C.J.; Qian, W.; Zhang, G.L. Icariin stimulates MC3T3-E1 cell proliferation and differentiation through up-regulation of bone morphogenetic protein-2. Int. J. Mol. Med. 2012, 29, 435–439. [Google Scholar] [PubMed]
- Liang, W.; Lin, M.; Li, X.; Li, C.; Gao, B.; Gan, H.; Yang, Z.; Lin, X.; Liao, L.; Yang, M. Icariin promotes bone formation via the BMP-2/Smad4 signal transduction pathway in the hFOB 1.19 human osteoblastic cell line. Int. J. Mol. Med. 2012, 30, 889–895. [Google Scholar] [PubMed]
- Zhao, J.; Ohba, S.; Shinkai, M.; Chung, U.I.; Nagamune, T. Icariin induces osteogenic differentiation in vitro in a BMP- and RUNX2-dependent manner. Biochem. Biophys. Res. Commun. 2008, 369, 444–448. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.N.; Ge, B.F.; Chen, K.M.; Zhou, J.; Shi, W.G.; Xie, Y.F.; Guo, X.Y.; Lv, X.; Cheng, K.; Gao, Y.H. Mechanisms of icariin in regulatng bone formation of osteoblasts and bone resorption of osteoclasts. J. Chin. Acad. Med. Sci. (Zhongguo Yixue Kexue Yuan Xuebao) 2013, 35, 432–438. [Google Scholar]
- Ma, X.N.; Zhou, J.; Ge, B.F.; Zhen, P.; Ma, H.P.; Shi, W.G.; Cheng, K.; Xian, C.J.; Chen, K.M. Icariin induces osteoblast differentiation and mineralization without dexamethasone in vitro. Planta Med. 2013, 79, 1501–1508. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.H.; Fung, C.Y.; Mok, S.K.; Wong, K.C.; Ho, M.X.; Wang, X.L.; Yao, X.S.; Wong, M.S. Flavonoids from Herba Epimedii selectively activate estrogen receptor α and stimulate ER-dependent osteoblastic functions in UMR-106 cells. J. Steroid Biochem. Mol. Biol. 2014, 143, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Zhao, J.; Zhang, X.; Li, H.; Zhou, Y. Icariin induces osteoblast proliferation, differentiation and mineralization through estrogen receptor-mediated ERK and JNK signal activation. Eur. J. Pharmacol. 2013, 714, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Ding, L.; Zhang, S.; Jiang, T.; Yang, Y.; Li, R. Effects of icariin on the regulation of the OPG-RANKL-RANK system are mediated through the MAPK pathways in IL-1β-stimulated human SW1353 chondrosarcoma cells. Int. J. Mol. Med. 2014, 34, 1720–1726. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Xia, L.; Zhou, Y.; Xu, Y.; Jiang, X. Icariin induces osteogenic differentiation of bone mesenchymal stem cells in a MAPK-dependent manner. Cell Prolif. 2015, 48, 375–384. [Google Scholar] [CrossRef] [PubMed]
- Bian, Q.; Huang, J.H.; Yang, Z.; Ning, Y.; Zhao, Y.J.; Wang, Y.J.; Shen, Z.Y. Effects of active ingredients in three kidney-tonifying chinese herbal drugs on gene expression profile of bone marrow stromal cells from a rat model of corticosterone-induced osteoporosis. Integr. Chin. West. Med. (Zhong Xi Yi Jie He Xuebao) 2011, 9, 179–185. [Google Scholar] [CrossRef]
- Ma, H.P.; Ma, X.N.; Ge, B.F.; Zhen, P.; Zhou, J.; Gao, Y.H.; Xian, C.J.; Chen, K.M. Icariin attenuates hypoxia-induced oxidative stress and apoptosis in osteoblasts and preserves their osteogenic differentiation potential in vitro. Cell Prolif. 2014, 47, 527–539. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.B.; Wang, J.W.; Xiao, H.; Zhang, Q.S.; Kan, W.S.; Mo, F.B.; Hu, S.; Ye, S.N. Icariin may benefit the mesenchymal stem cells of patients with steroid-associated osteonecrosis by ABCB1-promoter demethylation: A preliminary study. Osteoporos. Int. 2015, 26, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Liu, T.; Huang, Y.; Wismeijer, D.; Liu, Y. Icariin: Does it have an osteoinductive potential for bone tissue engineering? Phytother. Res. 2014, 28, 498–509. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Guo, Y.; Li, D.X.; Wang, R.; Fan, H.S.; Xiao, Y.M.; Zhang, L.; Zhang, X.D. The effect of loading icariin on biocompatibiltiy and bioactivity of porous β-TCP ceramic. J. Mater. Sci.: Matr. Med. 2011, 22, 371–379. [Google Scholar]
- Xie, X.; Pei, F.; Wang, H.; Tan, Z.; Yang, Z.; Kang, P. Icariin: A promising osteoinductive compound for repairing bone defect and osteonecrosis. J. Biomater. Appl. 2015, 30, 290–299. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Li, Y.; Zhou, Z.; Dai, Y.; Liu, H.; Liu, H. Icariin delivery porous phbv scaffolds for pormoting osteoblast expansion in vitro. Mater. Sci. Eng. 2013, 33, 3545–3552. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Zhu, M.; Zhu, S.; Wang, G.; Xu, Y.; Geng, D. Inhibitory effect of icariin on Ti-induced inflammatory osteoclastogenesis. J. Surg. Res. 2014, 192, 447–453. [Google Scholar] [CrossRef] [PubMed]
- Shao, H.; Shen, J.; Wang, M.; Cui, J.; Wang, Y.; Zhu, S.; Zhang, W.; Yang, H.; Xu, Y.; Geng, D. Icariin protects against titanium particle-induced osteolysis and inflammatory response in a mouse calvarial model. Biomaterials 2015, 60, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.C.; Sun, H.J.; Li, K.H.; Fu, C.; Liu, M.Z. Icariin promotes directed chondrogenic differentiation of bone marrow messenchymal stem cells but not hypertrophy in vitro. Exp. Ther. Med. 2014, 8, 1528–1534. [Google Scholar] [PubMed]
- Li, D.; Yuan, T.; Zhang, X.; Xiao, Y.; Wang, R.; Fan, Y.; Zhang, X. Icariin: A potential promoting compound for cartilage tissue engineering. Osteoarthr. Cartil. 2012, 20, 1647–1656. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.G.; Pang, L.; Chen, Z.R.; Tan, X.P. Dual-delivery of vanacomycin and icariin from an injectable calcium phosphate cement-release system for controlling infection and improving bone healing. Mol. Med. Rep. 2013, 8, 1221–1227. [Google Scholar] [PubMed]
- Wang, Q.S.; Zhang, X.C.; Li, R.X.; Sun, J.G.; Su, W.H.; Guo, Y.; Li, H.; Zhang, X.Z. A comparative study of mechanical strain, icariin and combination stimulations on improving osteoinductive potential via NF-κB activation in osteoblast-like cells. Biomed. Eng. Online 2015, 14, 46. [Google Scholar] [CrossRef] [PubMed]
- Xue, L.; Jiao, L.; Wang, Y.; Nie, Y.; Han, T.; Jiang, Y.; Rahman, K.; Zhang, Q.; Qin, L. Effects and interaction of icariin, curculigoside, and berberine in Er-Xian decoction, a traditional chinese medicinal formula, on osteoclastic bone resorption. Evid. Based Complement. Altern. Med. 2012, 490843. [Google Scholar] [CrossRef] [PubMed]
- Xue, L.; Wang, Y.; Jiang, Y.; Han, T.; Nie, Y.; Zhao, L.; Zhang, Q.; Qin, L. Comparative effects of Er-Xian decoction, Epimedium herbs, and icariin with estrogen on bone and reproductive tissue in ovariectomized rats. Evid. Based Complement. Altern. Med. 2012, 241416. [Google Scholar] [CrossRef]
- Chen, K.M.; Ge, B.F.; Liu, X.Y.; Ma, P.H.; Lu, M.B.; Bai, M.H.; Wang, Y. Icariin inhibits the osteoclast formation induced by RANKL and macrophage-colony stimulating factor in mouse bone marrow culture. Pharmazie 2007, 62, 388–391. [Google Scholar] [PubMed]
- Huang, J.; Yuan, L.; Wang, X.; Zhang, T.L.; Wang, K. Icaritin and its glycosides enhance osteoblastic, but suppress osteoclastic, differentiation and activity in vitro. Life Sci. 2007, 81, 832–840. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, T.P.; Sheu, S.Y.; Sun, J.S.; Chen, M.H. Icariin inhibits osteoblast differentiation and bone resoprtion by suppression of MAPKs/NF-κB regulated HIF-1α and PGE(2) synthesis. Phytomedicine 2011, 18, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Luo, J.G.; Kong, L.Y. An MC3T3-E1 cell line biomembrane extraction and HPLC-ESI-MSn method for simultaneous analysis of potential anti-osteoporosis components of Eipmedium koreanum. Chromatographia 2012, 75, 607–615. [Google Scholar] [CrossRef]
- Zhan, Y.; Wei, Y.J.; Sun, E.; Xu, F.J.; Jia, X.B. Two-dimensional zebrafish model combined with hyphenated chromatographic techniques for evaluation of antiosteoporosis activity of epimedin A and its metabolite baohuoside. Acta Pharm. Sin. (Yaoxue Xuebao) 2014, 49, 932–937. [Google Scholar]
- Zhang, D.W.; Zhang, J.; Fong, C.; Yao, X.S.; Yang, M. Herba Epimedii flavonoids suppress osteoclastic differentiation and bone resoption by inducing G2/M arrest and apoptosis. Biochimie 2012, 94, 2514–2522. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Wang, N.L.; Cai, G.P. Maohuoside a promotes osteogenesis of rat mesenchymal stem cells via BMP and MAPK signaling pathways. Mol. Cell. Biochem. 2011, 358, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Cai, M.; Li, G.; Tao, K.; Yang, Y.; Lou, L.; Cai, Z.; Yu, Y. Maohuoside a acts in a BMP-dependent manner during osteogenesis. Phytother. Res. 2013, 27, 1179–1184. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.J.; Park, Y.R.; Nepal, M.; Choi, B.Y.; Cho, N.P.; Choi, S.H.; Heo, S.R.; Kim, H.S.; Yang, M.S.; Soh, Y. Inhibition of osteoclastogenic differentiation by ikarisoside A in RAW 264.7 cells via JNK and NF-κB signaling pathways. Eur. J. Pharmacol. 2010, 636, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Zhang, Y.; Yang, M.; Shen, Z.; Zhang, X.; Zhang, W.; Li, H. LC-MS/MS method for the simultaneous determination of icariin and its major metabolites in rat plasma. J. Pharm. Biomed. Anal. 2007, 45, 667–672. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Fan, M.; Fan, L.; Sun, J.G.; Guo, D. Liquid chormatography-tandem mass spectrometry analysis of metabolites in rats after administration of prenylflavonoids from Epimedium. J. Chromatogr. B 2010, 878, 1113–1124. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Y.K.; Ge, B.F.; Chen, K.M.; Ma, H.P.; Ming, L.G.; Li, Z.F. Comparative study on the osteogenic differentiation of rat bone marrow stromal cells effected by icariin and icariside II. Zhongyaocai 2010, 33, 1896–1900. [Google Scholar] [PubMed]
- Yao, D.; Xie, X.H.; Wang, X.L.; Wan, C.; Lee, Y.W.; Chen, S.H.; Pei, D.Q.; Wang, Y.X.; Li, G.; Qin, L. Icaritin, an exogenous phytomolecule, enhances osteogenesis but not antiogenesis - an in vitro efficacy study. PLoS ONE 2012, 7, e41264. [Google Scholar]
- Sheng, H.; Rui, X.F.; Sheng, C.J.; Li, W.J.; Cheng, X.Y.; Jhummon, N.P.; Yu, Y.C.; Qu, S.; Zhang, G.; Qin, L. A novel semisynthetic molecule icaritin stimulates osteogenic defferentiation and inhibits adipogenesis of mesenchymal stem cells. Int. J. Med. Sci. 2013, 10, 782–789. [Google Scholar] [CrossRef] [PubMed]
- Peng, S.; Zhang, G.; Zhang, B.T.; Guo, B.; He, Y.; Bakker, A.J.; Pan, X.; Zhen, W.; Hung, L.; Qin, L.; et al. The beneficial effect of icaritin on osteoporotic bone is dependent on the treatment initiation timing in adult ovariectomized rats. Bone 2013, 55, 230–240. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Qin, L.; Shang, H.; Wang, X.L.; Wang, Y.X.; Yeung, D.K.; Griffith, J.F.; Yao, X.S.; Xie, X.H.; Li, Z.R.; et al. A novel semisynthesized small molecule icaritin reduces incidence of steroid-associated osteonecrosis with inhibition of both thrombosis and lipid-deposition in a dose-dependent manner. Bone 2009, 44, 345–356. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.Q.; Yang, Q.X.; Cheng, M.C.; Xiao, H.B. Synergistic inhibitory effect of icariside II with icaritin from Herba Epimedii on pre-osteoclastic RAW264.7 cell growth. Phytomedicine 2014, 21, 1633–1637. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.H.; Wang, X.L.; Zhang, G.; He, Y.X.; Wang, X.H.; Liu, Z.; He, K.; Peng, J.; Leng, Y.; Qin, L. Structural and degradation characteristics of an innovative porous PLGA/TCP scaffold incorporated with bioactive molecular icaritin. Biomed. Mater. 2010, 5, 054109. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.H.; Wang, X.L.; Xie, X.H.; Zheng, L.Z.; Yao, D.; Wang, D.P.; Leng, Y.; Zhang, G.; Qin, L. Comparative study of osteogenic potential of a composite scaffold incorporating either endogenous bone morphogenetic protein-2 or exogenous phytomolecule icaritin: An in vitro efficacy study. Acta Biomater. 2012, 8, 3128–3137. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.H.; Wang, X.L.; Zhang, G.; He, Y.X.; Leng, Y.; Tang, T.T.; Pan, X.; Qin, L. Biofabrication of a PLGA-TCP-based porous bioactive bone substitute with sustained release of icaritin. J. Tissue Eng. Regen. Med. 2015, 9, 961–972. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.H.; Lei, M.; Xie, X.H.; Zhang, L.Z.; Yao, D.; Wang, X.L.; Li, W.; Zhao, Z.; Kong, A.; Xiao, D.M.; et al. PLGA/TCP composite scaffold incorporating bioactive phytomolecule icaritin for enhancement of bone defect repair in rabbits. Acta Biomater. 2013, 9, 6711–6722. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.L.; Xie, X.H.; Zhang, G.; Chen, S.H.; Yao, D.; He, K.; Wang, X.H.; Yao, X.S.; Leng, Y.; Fung, K.P.; et al. Exogenous phytoestrogenic molecule icaritin incorporated into a porous scaffold for enhancing bone defect repair. J. Orthop. Res. 2013, 31, 164–172. [Google Scholar] [CrossRef] [PubMed]
- Qin, L.; Yao, D.; Zheng, L.; Liu, W.C.; Liu, Z.; Lei, M.; Huang, L.; Xie, X.; Wang, X.; Chen, Y.; et al. Phytomolecule icaritin incorporated PLGA/TCP scaffold for steroid-associated osteonecrosis: Proof-of-concept for prevention of hip joint collapse in bipedal emus and mechanistic study in quadrupedal rabbits. Biomaterials 2015, 59, 125–143. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, X.L.; Lai, W.P.; Chen, B.; Chow, H.K.; Wu, C.F.; Wang, N.L.; Yao, X.S.; Wong, M.S. Anti-osteoporotic effect of Erythrina variegata L. in ovariectomized rats. J. Ethnopharmacol. 2007, 109, 165–169. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, Q.; Li, X.L.; Wan, H.Y.; Wong, M.S. Erythrina variegata extract exerts osteoprotective effects by suppression of the process of bone resorption. Br. J. Nutr. 2010, 104, 965–971. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, X.L.; Yao, X.S.; Wong, M.S. Osteogenic activities of genistein derivatives were influenced by the presence of prenyl group at ring A. Arch. Pharm. Res. 2008, 31, 1534–1539. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Liu, Z.G.; Li, C.; Hu, S.J.; Liu, L.; Wang, J.P.; Mei, Q.B. Du-zhong (Eucommia ulmoides Oliv.) cortex extract prevents OVX-induced osteoporosis in rats. Bone 2009, 45, 553–559. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Lee, J.I.; Song, M.; Lee, D.; Song, J.; Kim, S.Y.; Park, J.; Choi, H.Y.; Kim, H. Effects of Eucommia ulmoides extract on longitudinal bone growth rate in adolescent female rats. Phytother. Res. 2015, 29, 148–153. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Niu, Y.; Li, C.; Zhai, Y.; Zhang, R.; Guo, X.; Mei, Q. Du-zhong (Eucommia ulmoides) prevents disuse-induced osteoporosis in hind limb suspension rats. Am. J. Chin. Med. 2014, 42, 143–155. [Google Scholar] [CrossRef] [PubMed]
- Ha, H.; Ho, J.; Shin, S.; Kim, H.; Kook, S.; Kim, I.H.; Kim, C. Effects of Eucommiae Cortex on osteoblast-like cell proliferation and osteoclast inhibition. Arch. Pharm. Res. 2003, 26, 929–936. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Pan, Y.L.; Hu, S.J.; Kong, X.H.; Juan, W.; Mei, Q.B. Effects of total lignans from Eucommia ulmoides barks prevent bone loss in vivo and in vitro. J. Ethnopharmacol. 2014, 155, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Tan, X.L.; Zhang, Y.H.; Cai, J.P.; Zhu, L.H.; Ge, W.J.; Zhang, X. 5-(Hydroxymethyl)-2-furaldehyde inhibits adipogenic and enhances osteogenic differentiation of rat bone mesenchymal stem cells. Nat. Prod. Commun. 2014, 9, 529–532. [Google Scholar] [PubMed]
- Lin, J.; Fan, Y.J.; Mehl, C.; Zhu, J.J.; Chen, H.; Jin, L.Y.; Xu, J.H.; Wang, H.M. Eucommia ulmoides Oliv. antagonizes H2O2-induced rat osteoblastic MC3T3-E1 apoptosis by inhibiting expressions of capases 3, 6, 7 and 9. J. Zhejiang Univ. Sci. B 2011, 12, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Fujikawa, T.; Mizuno, K.; Ishida, T.; Ooi, K.; Hirata, T.; Wada, A. Eucommia leaf extract (ELE) prevents OVX-induced osteoporosis and obesity in rats. Am. J. Chin. Med. 2012, 40, 735–752. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, M.J.; Li, S.; Zhang, Y.M.; Zhao, Y.; Xie, R.M.; Sun, W.J. Effect of total glycosides from Eucommia ulmoides seed on bone microarchitecture in rats. Phytother. Res. 2011, 25, 1895–1897. [Google Scholar] [CrossRef] [PubMed]
- Che, C.T.; Wong, M.S. Ligustrum lucidum and its constituents: A mini-review on the anti-osteoporosis potential. Nat. Prod. Commun. 2015, 10, 2189–2194. [Google Scholar] [PubMed]
- Li, J.X.; Hareyama, T.; Tezuka, Y.; Zhang, Y.; Miyahara, T.; Kadota, S. Five new oleanolic acid glycosides from Achyranthes bidentata with inhibitory activity on osteoclast formation. Plant Med. 2005, 71, 673–679. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, J.X.; Zhao, J.; Wang, S.Z.; Pan, Y.; Tanaka, K.; Kadota, S. Synthesis and activity of oleanolic acid derivatives, a novel class of inhibitors of osteoclast formation. Bioorg. Med. Chem. Lett. 2005, 15, 1629–1632. [Google Scholar] [CrossRef] [PubMed]
- Li, J.F.; Zhao, Y.J.; Cai, M.M.; Li, X.F.; Li, J.X. Synthesis and evaluation of a novel series of heterocyclic oleanolic acid derivatives with anti-osteoclast formation activity. Eur. J. Med. Chem. 2009, 44, 2796–2806. [Google Scholar] [CrossRef] [PubMed]
- Li, J.F.; Chen, S.J.; Zhao, Y.; Li, J.X. Glycoside modification of oleanolic acid derivatives as a novel class of anti-osteoclast formation agents. Carbohydr. Res. 2009, 344, 599–605. [Google Scholar] [CrossRef] [PubMed]
- Bian, Q.; Liu, S.F.; Huang, J.H.; Yang, Z.; Tang, D.Z.; Zhou, Q.; Ning, Y.; Zhao, Y.J.; Lu, S.; Shen, Z.Y. Oleanolic acid exerts an osteoprotective effect in ovariectomy-induced osteoporotic rats and stimulates the osteoblasic differentiation of bone mesenchymal stem cells in vitro. Menopause 2012, 19, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Cheon, Y.H.; Oh, H.M.; Rho, M.C.; Erkhembaatar, M.; Kim, M.S.; Lee, C.H.; Kim, J.J.; Choi, M.K.; Yoon, K.H.; et al. Oleanolic acid acetate inhibits osteoclast differentiation by downregulating PLCγ2-Ca(2+)-NFATC1 signaling and suppresses bone loss in mice. Bone 2014, 60, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.U.; Park, S.J.; Kwak, H.B.; Oh, J.; Min, Y.K.; Kim, S.H. Anabolic activity of ursolic acid in bone: Stimulating osteoblast differentiation in vitro and inducing new bone formation in vivo. Pharmacol. Res. 2008, 58, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Tan, H.; Furuta, S.; Nagata, T.; Ohnuki, K.; Akasaka, a.; Shirouchi, B.; Sato, M.; Kondo, R.; Shimizu, K. Inhibitory effects of the leaves of ioquat (Eriobotrya japonica) on bone mineral density loss in ovariectomized mice and osteoclast differentiation. J. Agric. Food Chem. 2014, 62, 836–841. [Google Scholar] [CrossRef] [PubMed]
- Tan, H.; Ashour, A.; Katakura, Y.; Shimizu, K. A structure-activity relationship study on anti-osteoclastogenesis effect of triterpenoids from the leaves of ioquat (Eriobotrya japonica). Phytomedicine 2015, 22, 498–503. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Xiao, F.; Gu, X.; Zhai, Z.; Liu, X.; Wang, W.; Tang, T.; Wang, Y.; Zhu, Z.; Dai, K.; et al. Inhibitory effects of ursolic acid on osteoclastogenesis and titanium particle-induced osteolysis are mediated primarily via suppression of NF-κB signaling. Biochimie 2015, 111, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Yang, L.; Zhang, G.; Wang, F. Bioactivity-guided isolation of antiosteoporotic compounds from Ligustrum lucidum. Phytother. Res. 2013, 27, 973–979. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Wu, Y.; Wu, J.; Yi, J.; Zhang, Q.; Chen, T.; Wu, J. Chemical constituents from Ligustrum lucidum differentially promote bone formation and prevent oxidative damage in osteoblastic UMR-106 cells. Lat. Am. J. Pharmacy 2014, 33, 258–265. [Google Scholar]
- Garcia-Martinez, O.; Rivas, A.; Ramos-Torrecillas, J.; de Luna-Bertos, E.; Ruiz, C. The effect of olive oil on osteoporosis prevention. Int. J. Food Sci. Nutr. 2014, 65, 834–840. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.C.; Wang, S.Q.; Zhao, S. The effect of salidroside on leptin expression of diabetic rats with osteoporosis. Chin. J. Pathophysiol. 2009, 25, 787–788. [Google Scholar]
- Seo, B.I.; Ku, S.K.; Cha, E.M.; Park, J.H.; Kim, J.D.; Choi, H.Y.; Lee, H.S. Effect of Morindae Radix extracts on experimental osteoporosis in sciatic neurectomized mice. Phytother. Res. 2005, 19, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.Y.; Wang, C.J.; Zhang, H.S.; Pei, X.W.; Fen, J.M. Protective effect of polysaccharides from Morinda officinalis on bone loss in ovariectomzied rats. Int. J. Biol. Macromol. 2008, 43, 276–278. [Google Scholar]
- Li, N.; Qin, L.P.; Han, T.; QWu, Y.B.; Zhang, Q.Y.; Zhang, H. Inhibitory effects of Morinda officinalis extract on bone loss in ovariectomized rats. Molecules 2009, 14, 2049–2061. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.B.; Zheng, C.J.; Qin, L.P.; Sun, L.N.; Han, T.; Jiao, L.; Zhang, Q.Y.; Wu, J.Z. Antiosteoporotic activity of anthraquinones from Morinda officinalis on osteoblasts and osteoclasts. Molecules 2009, 14, 573–583. [Google Scholar] [CrossRef] [PubMed]
- Bao, L.; Qin, L.; Liu, L.; Wu, Y.; Han, T.; Xue, L.; Zhang, Q. Anthraquinone compounds from Morinda officinalis inhibit osteoclastic bone resoprtion in vitro. Chem. Biol. Interations 2011, 194, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Liang, J.; Zheng, C.; Hu, C.; Zhao, X.; Rahman, K.; Qin, L. Phenylpropanoids from Podocarpium podocarpum. Pharm. Biol. 2013, 51, 1021–1025. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.Q.; Zheng, C.J.; Zhang, Y.; Hu, C.L.; Lin, B.; Fu, X.Y.; Han, L.Y.; Xu, L.S.; Rahman, K.; Qin, L.P. Antiosteoporotic flavonoids from Podcarpium podocarpum. Phytochem. Lett. 2013, 6, 118–122. [Google Scholar] [CrossRef]
- Ye, Q.; Ma, X.Q.; Hu, C.L.; Lin, B.; Xu, L.S.; Zheng, C.J.; Qin, L.P. Antiosteoporotic activity and consituents of Podocarpium podocarpum. Phytomedicine 2015, 22, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Bhargavan, B.; Gautam, A.K.; Singh, D.; Kumar, A.; Chaurasia, S.; Tyagi, A.M.; Yadav, D.K.; Mishra, J.S.; Singh, A.B.; Sanyal, S.; et al. Methoxylated isoflavones, cajanin and isoformononetin, have non-estrogenic bone forming effect via differential mitogen activated protein kinase (MAPK) signaling. J. Cell. Biochem. 2009, 108, 388–399. [Google Scholar] [CrossRef] [PubMed]
- Tsai, M.H.; Huang, G.S.; Hung, Y.C.; Bin, L.; Liao, L.T.; Lin, L.W. Psoralea corylifolia extract ameliorates experimental osteoporosis in ovariectomized rats. Am. J. Chin. Med. 2007, 35, 669–680. [Google Scholar] [CrossRef] [PubMed]
- Wong, R.W.; Rabie, A.B. Systemic effect of Fructus Psoraleae extract on bone in mice. Phytother. Res. 2010, 24, 1578–1580. [Google Scholar] [CrossRef] [PubMed]
- Khushboo, P.S.; Jadhav, V.M.; Kadam, V.J.; Sathe, N.S. Psoralea corylifolia Linn.—“Kushtanashini”. Phyarmacogn. Rev. 2010, 4, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Chopra, B.; Dhingra, A.K.; Dhar, K.L. Psoralea corylifolia L. (Buguchi)—folklore to modern evidence: Review. Fitoterapia 2013, 90, 44–56. [Google Scholar] [CrossRef] [PubMed]
- Xin, D.; Wang, H.; Yang, J.; Su, Y.F.; Fan, G.W.; Wang, Y.F.; Zhu, Y.; Gao, X.M. Phytoestrogens from Psoralea corylifolia reveal estrogen receptor-subtype selectivity. Phytomedicine 2010, 17, 126–131. [Google Scholar] [CrossRef] [PubMed]
- Wong, R.W.; Rabie, A.B. Effect of psoralen on bone formation. J. Orthop. Res. 2011, 29, 158–164. [Google Scholar] [CrossRef] [PubMed]
- Wong, R.W.; Rabie, A.B. Effect of Buguzhi (Psoralea corylifolia fruit) extract on bone formation. Phytother. Res. 2010, 24 (Suppl. 2), S155–S160. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Huang, J.H.; Liu, S.F.; Zhao, Y.J.; Shen, Z.Y.; Wang, Y.J.; Bian, Q. The osteoprotective effect of psoralen in ovariectomy-induced osteoporotic rats via stimulaitng the osteoblastic differentiation from bone messenchymal stem cells. Menopause 2012, 19, 1156–1164. [Google Scholar] [CrossRef] [PubMed]
- Tang, D.Z.; Yang, F.; Yang, Z.; Huang, J.; Shi, Q.; Chen, D.; Wang, Y.J. Psoralen stimulates osteoblast differentiation through activation of BMP signaling. Biochem. Biophys. Res. Commun. 2011, 405, 256–261. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Pan, X.; Sun, Y.; Xu, W.; Njunge, L.; Yang, L. Psoralen activates cartilaginous cellular functions of rat chondrocytes in vitro. Pharm. Biol. 2015, 53, 1010–1015. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Sun, X.; Geng, X. Effects of psoralen on chondrocyte degeneration in lumbar intervertebral disc of rats. Pak. J. Pharm. Sci. 2015, 28 (Suppl. 2), 667–670. [Google Scholar] [PubMed]
- Ming, L.; Ge, B.; Chen, K.; Ma, H.; Zhai, Y. Effects of isopsoralen on bone marrow stromal stem cells differentiation and proliferation in vitro. China J. Chin. Med. (Zhongguo Zhongyao Zazhi) 2011, 36, 2124–2128. [Google Scholar]
- Ming, L.G.; Cheng, K.M.; Ge, B.F.; Ma, H.P.; Zai, Y.K. Effect of isopsoralen on the proliferation and differentiation of osteoblasts in vitro. Chin. Med. Mater. (Zhongyaocai) 2011, 34, 404–408. [Google Scholar]
- Li, J.P.; Wang, X.J.; Zeng, Y.; Lin, Q.; Mo, X.M.; Liu, S.J.; Yang, J. Study on the effect of psoralidin on anti-experimental postmenopausal osteoporosis and its mechanism. China J. Chin. Med. (Zhongguo Zhongyao Zazhi) 2013, 38, 1816–1819. [Google Scholar]
- Wang, D.; Li, F.; Jiang, Z. Osteoblastic proliferation stimulating activity of Psoralea corylifolia extracts and two of its flavonoids. Planta Med. 2001, 67, 748–749. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Z.; Wang, D.; Xu, Y.; Li, F. Osteoblastic differentiation bioassay and its application to investigating the activity of fractions and compounds from Psoralea corylifolia L. Pharmazie 2003, 58, 925–928. [Google Scholar] [PubMed]
- Li, W.D.; Yan, C.P.; Wu, Y.; Weng, Z.B.; Yin, F.Z.; Yang, G.M.; Cai, B.C.; Chen, Z.P. Osteoblasts proliferation and differentiation stimulting activities of the main components of Fructus Psoraleae corylifoliae. Phytomedicine 2014, 21, 400–405. [Google Scholar] [CrossRef] [PubMed]
- Don, M.J.; Lin, L.C.; Chiou, W.F. Neobavaisoflavone stimulates osteogenesis via p38-mediated up-regulation of transcription factors and osteoid genes expression in MC3T3-E1 cells. Phytomedicine 2012, 19, 551–561. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.H.; Ha, T.Y.; Kim, S.R.; Ahn, J.; Park, H.J.; Kim, S. Ethanol extract of Psoralea corylifolia L. and its main constituent, bakuchiol, reduce bone loss in ovariectomized Sprague-Dawley rats. Br. J. Nutr. 2009, 101, 1031–1039. [Google Scholar] [CrossRef] [PubMed]
- Park, C.K.; Lee, Y.C.; Chang, E.J.; Lee, M.H.; Yoon, J.H.; Ryu, J.H.; Kim, H.H. Bavachalcone inhibits osteoclast differentiation through suppression of NFATc1 induction by RANKL. Biochem. Pharmacol. 2008, 75, 2175–2182. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.X.; Zhang, H.; Peng, C. Puerarin: A review of pharmacological effects. Phytother. Res. 2014, 28, 961–975. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.Y.; Chen, Y.; Xu, X.Y. Progress on the pharmacological research of puerarin: A review. Chin. J. Nat. Med. 2014, 12, 407–414. [Google Scholar] [CrossRef]
- Wang, X.; Wu, J.; Chiba, H.; Umegaki, K.; Yamada, K.; Ishimi, Y. Puerariae Radix prevents bone loss in ovariectomized mice. J. Bone Miner. Metab. 2003, 21, 268–275. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Jin, B.Q.; Sun, G.J.; Du, X.F.; Wan, X.C. Effects of puerarin on bone metabolism in ovariectomized rats. Chin. J. Geratol. (Zhongguo Laonianxue Zazhi) 2009, 29, 2482–2484. [Google Scholar]
- Wang, P.P.; Zhu, X.F.; Yang, L.; Liang, H.; Feng, S.W.; Zhang, R.H. Puerarin stimulates osteoblasts differentiation and bone formation through estrogen receptor, p38 MAPK, and Wnt/β-catenin pathways. J. Asian Nat. Prod. Res. 2012, 14, 897–905. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Li, B.B. Effect of puerarin on osteoporosis resulted from ovariectomy in rats. Chin. J. Comp. Med. (Zhongguo Bijiao Yixue Zazhi) 2012, 22, 16–20. [Google Scholar]
- Li, B.; Liu, H.; Jia, S. Puerarin enhances bone mass by promoting osteoblastogenesis and slightly lowering bone marrow adiposity in ovariectomized rats. Biol. Pharm. Bull. 2014, 37, 1919–1925. [Google Scholar] [CrossRef] [PubMed]
- Wong, R.W.; Rabie, B. Effect of puerarin on bone formation. Osteoarthr. Cartil. 2007, 15, 894–899. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.Y.; Qiang, H.; Yang, H.Q.; Dang, X.Q.; Wang, K.Z. In vitro and in vivo effects of puerarin on promotion of osteoblast bone formation. Chin. J. Integr. Med. 2012, 18, 276–282. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Chen, H.; Pan, W.; Xu, C. Puerarin inhibits caspase-3 expression in osteoblasts of diabetic rats. Mol. Med. Rep. 2012, 5, 1419–1422. [Google Scholar] [PubMed]
- Wang, Y.; Yin, L.; Li, Y.; Liu, P.; Cui, Q. Preventive effects of puerarin on alcohol-induced osteonecrosis. Clin. Orthop. Relat. Res. 2008, 466, 1059–1067. [Google Scholar] [CrossRef] [PubMed]
- Michihara, S.; Tanaka, T.; Uzawa, Y.; Moriyama, T.; Kawamura, Y. Puerarin exerted anti-osteoporotic action independent of estrogen receptor-mediated pathway. J. Nutr. Sci. Vitaminol. 2012, 58, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Li, B.B. Inhibitory effect of puerarin on osteoclastic bone resorption induced by 1,25-dihydroxyvitamin D3 in vitro: The optimal concentration of puerarin. Chin. Res. Tissue Eng. (Zhongguo Zuzhi Gongcheng Yanjiu) 2012, 28, 5141–5144. [Google Scholar]
- Zhang, Y.; Zeng, X.; Zhang, L.; Zheng, X. Stimulatory effect of puerarin on bone formation through activation of PI3K/AKT pathway in rat calvaria osteoblasts. Planta Med. 2007, 73, 341–347. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Meng, M.X.; Tang, X.L.; Chen, K.M.; Zhang, L.; Liu, W.N.; Zhao, Y.Y. The proliferation, differentiation, and mineralization effects of puerarin on osteoblasts in vitro. Chin. J. Nat. Med. 2014, 12, 436–442. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, W.L.; Xie, W.L.; Li, L.Z.; Sun, J.; Sun, W.J.; Gong, H.Y. Puerarin stimulates proliferation and differentiation and protects against cell dealth in human osteoblastic MG-63 cells via ER-dependent MEK/ERK and PI3K/AKT activation. Phytomedicine 2013, 20, 787–796. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.J.; Liu, L.Q.; Bo, T.; Li, S.J.; Zhu, Z.; Cui, R.R.; Mao, D.A. Puerarin suppresses apoptosis of human osteoblasts via ERK signaling pathway. Int. J. Endocrinol. 2013, 786574. [Google Scholar] [CrossRef]
- Sheu, S.Y.; Tsai, C.C.; Sun, J.S.; Chen, M.H.; Liu, M.H.; Sun, M.G. Stimulatory effect of puerarin on bone formation through co-activation of nitric oxide and bone morphogenetic protein-2/mitogen-activated protein kinases pathways in mice. Chin. Med. J. (Engl. Ed.) 2012, 125, 3646–3653. [Google Scholar]
- Wang, Y.; Yang, C.; Xie, W.L.; Zhao, Y.W.; Li, Z.M.; Sun, W.J.; Li, L.Z. Puerarin concurrently stimulates osteoprotegerin and inhibits receptor activator of NF-κB ligand (RANKL) and interleukin-6 production in human osteoblastic MG-63 cells. Phytomedicine 2014, 21, 1032–1036. [Google Scholar] [CrossRef] [PubMed]
- Lv, H.; Che, T.; Tang, X.L.; Liu, L.; Cheng, J. Puerarin enhances proliferation and osteoblastic differentiation of human bone marrow stromal cells via a nitric oxide/cyclic guanosine monophosphate signaling pathway. Mol. Med. Rep. 2015, 12, 2283–2290. [Google Scholar] [CrossRef] [PubMed]
- Zhan, X.Q.; Qian, K.Q.; Sun, Y.M. Effect of puerarin on TGF-β1/SMAD pathway in MC3T3-E1 cells. Chin. Patent Drugs (Zhongchenyao) 2013, 35, 1121–1124. [Google Scholar]
- Tiyasatkulkovit, W.; Charoenphandhu, N.; Wongdee, K.; Thongbunchoo, J.; Krishnamra, N.; Malaivijitnond, S. Upregulation of osteoblastic differentiation marker mRNA expression in osteoblast-like UMR 106 cells by puerarin and phytoestrogens from Pueraria mirifica. Phytomedicine 2012, 19, 1147–1155. [Google Scholar] [CrossRef] [PubMed]
- Tiyasatkulkovit, W.; Malaivijitnond, S.; Charoenphandhu, N.; Havill, L.M.; Ford, A.L.; VandeBerg, J.L. Pueraria mirifica extract and puerarin enhance proliferation and expression of alkaline phosphatase and type 1 collagen in primary baboon osteoblasts. Phytomedicine 2014, 21, 1498–1503. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Zhang, H.; Wang, J.; Zhang, Z.; Li, C. Puerarin decreases bone loss and collagen destruction in rats with ligature-induced periodontitis. J. Periodontal Res. 2015, 50, 748–757. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Chen, B.; Pang, G.; Chen, J.; Xie, J.; Huang, H. Anti-osteoporotic activity of puerarin 6′′-O-xyloside on ovariectomized mice and its potential mechanisms. Pharm. Biol. 2016, 54, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Oh, K.O.; Kim, S.W.; Kim, J.Y.; Ko, S.Y.; Kim, H.M.; Baek, J.H.; Ryoo, H.M.; Kim, J.K. Effect of Rehmannia glutinosa Libosch extracts on bone metabolism. Clin. Chim. Acta 2003, 334, 185–195. [Google Scholar] [CrossRef]
- Lim, D.W.; Kim, Y.T. Dried root of Rehmannia glutinosa prevents bone loss in ovariectomized rats. Molecules 2013, 18, 5804–5813. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y.; Lee, K.S.; Yi, S.H.; Kook, S.H.; Lee, J.C. Acteoside suppresses RANKL-mediated osteoclastogenesis by inhibiting c-Fos induction and NF-κB pathway and attenuating ROS production. PLoS ONE 2013, 8, e80873. [Google Scholar] [CrossRef] [PubMed]
- Kiem, P.V.; Quang, T.H.; Huong, T.T.; Nhung, L.T.H.; Nguyen, X.C.; Minh, C.V.; Choi, E.M.; Kim, Y.H. Chemical constituents of Acanthus ilicifolius L. And effect on osteoblastic MC3T3-E1 cells. Arch. Pharm. Res. 2008, 31, 823–829. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.A.; Lee, S.A.; Kim, K.H.; Lee, K.S.; Lee, J.C. Acteoside inhibits irradiation-mediated decreases in the viability and DNA synthesis of MC3T3-E1 cells. Food Sci. Biotech. 2013, 22, 845–851. [Google Scholar] [CrossRef]
- Chae, H.J.; Chae, S.W.; Yun, D.H.; Keum, K.S.; Yoo, S.K.; Kim, H.R. Prevention of bone loss in ovariectomized rats: The effect of Salvia miltiorrhiza extracts. Immunopharmacol. Immunotoxicol. 2004, 26, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.; Zou, L.Y.; Liu, Y.Y.; Ai, C.M.; Wu, T.; Wang, Y. Preventing cancellous bone loss in steroid-treated rats and stimulating bone formation by water extract of Salvia miltiorrhiza and danshensu. Chin. Pharmacol. Bull. 2004, 20, 286–291. [Google Scholar]
- Miao, B.; Wang, J.; Zhu, Y.; Yue, C.; Chen, M. Experimental study on the effect of Salvia miltiorrhiza on alveolar bone metabolism and variation in bone mass in diabetic rats. Zhongguo Zhongyao Zazhi 2012, 37, 1659–1662. [Google Scholar] [PubMed]
- Wong, R.W.; Rabie, A.B. Effect of danshen extract on bone formation. Ann. R. Aust. Coll. Dent. Surg. 2008, 19, 171–174. [Google Scholar]
- Wu, Y.B.; Ni, Z.Y.; Shi, Q.W.; Dong, M.; Kiyota, H.; Gu, Y.C.; Cong, B. Constituents from Salvia species and their biological activities. Chem. Rev. 2012, 112, 5967–6026. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.; Liu, Y.Y.; Wu, T.; Ai, C.M.; Chen, H.Q. Osteogenic effects of d-β-3,4-dihydroxyphenyl lactic acid (salvianic acid A, SAA) on osteoblasts and bone marrow stromal cells of intact and prednisone-treated rats. Acta Pharmacol. Sin. 2009, 30, 321–332. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.; Li, T.; Liu, Y.; Zhou, L.; Li, P.; Xu, B.; Huang, L.; Chen, Y.; Liu, Y.; Tian, X.; et al. Salvianolic acid B prevents bone loss in prednisone-treated rats through stimulation of osteogenesis and bone marrow angiogenesis. PLoS ONE 2012, 7, e34647. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.; Wu, T.; Liu, Y.Y.; Deng, Y.F.; Al, C.M.; Chen, H.Q. Tanshinone prevents cancellous bone loss induced by ovariectomy in rats. Acta Pharmacol. Sin. (Yaoli Xuebao) 2004, 25, 678–684. [Google Scholar]
- Lee, S.Y.; Choi, D.Y.; Woo, E.R. Inhibition of osteoclast differentiation by tanshinones from the root of Salvia miltiorrihiza Bunge. Arch. Pharm. Res. 2005, 28, 909–913. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.K.; Woo, E.R.; Lee, H.W.; Park, H.R.; Kim, H.N.; Jung, Y.K.; Choi, J.Y.; Chae, S.W.; Kim, H.R.; Chae, H.J. The correlation of Salvia miltiorrhiza extract-induced regulation of osteoclastogenesis with the amount of components tanshinone I, tanshinone IIa, cryptotanshinone, and dihydrotanshinone. Immunopharmacol. Immunotoxicol. 2008, 30, 347–364. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.H.; Kim, J.H.; Kwak, H.B.; Huang, H.; Han, S.H.; Ha, H.; Lee, S.W.; Woo, E.R.; Lee, Z.H. Inihbition of osteoclast differentiation and bone resorption by tanshinone IIa isolated from Salvia miltiorrhiza Bunge. Biochem. Pharmacol. 2004, 67, 1647–1656. [Google Scholar] [CrossRef] [PubMed]
- Kwak, H.B.; Yang, D.; Ha, H.; Lee, J.H.; Kim, H.N.; Woo, E.R.; Lee, S.; Kim, H.H.; Lee, Z.H. Tanshinone IIa inhibits osteoclast differentiation through down-regulation of c-Fos and NFATc1. Exp. Mol. Med. 2006, 38, 256–264. [Google Scholar] [CrossRef] [PubMed]
- Kwak, H.B.; Sun, H.M.; Ha, H.; Kim, H.N.; Lee, J.H.; Kim, H.H.; Shin, H.I.; Lee, Z.H. Tanshinone IIa suppresses inflammatory bone loss by inhibiting the sysnthesis of prostaglandin E2 in osteoblasts. Eur. J. Pharmacol. 2008, 601, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Nicolin, V.; Dal-Piaz, F.; Nori, S.L.; Narducci, P.; De Tommasi, N. Inhibition of bone resorption by tanshinone VI isolated from Salvia miltiorrhiza Bunge. Eur. J. Histochem. 2010, 54, e21. [Google Scholar] [CrossRef] [PubMed]
- Shama, V.; Panwar, P.; O’Donoghue, A.J.; Cui, H.; Guido, R.V.; Craik, C.S.; Bromme, D. Structural requirements for the collagenase and elastase activity of cathepsin K and its selective inhibition by an exosite inhibitor. Biochem. J. 2015, 465, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Chin, A.; Yang, Y.; Chai, L.; Wong, R.W.; Rabie, A.B. Effects of medicinal herb Salvia miltiorrhiza on osteoblastic cells in vitro. J. Orthop. Res. 2011, 29, 1059–1063. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Xu, L.; Zhou, C.; Lee, W.Y.; Wu, T.; Cui, L.; Li, G. Salvianolic acid B promotes osteogenesis of human mesenchymal stem cells through activating ERK signaling pathway. Int. J. Biochem. Cell Biol. 2014, 51, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Xie, F.; Wu, C.F.; Zhang, Y.; Yao, X.S.; Cheung, P.Y.; Chan, A.S.; Wong, M.S. Increase in bone mass and bone strength by Sambucus williamsii Hance in ovariectomized rats. Biol. Pharm. Bull. 2005, 28, 1879–1885. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, Q.; Wan, H.Y.; Xiao, H.H.; Lai, W.P.; Yao, X.S.; Wong, M.S. Study of the mechanisms by which Sambucus williamsii Hance extract exert protective effects against ovariectomy-induced osteoporosis in vivo. Osteoporos. Int. 2011, 22, 703–709. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.H.; Dai, Y.; Wan, H.Y.; Wong, M.S.; Yao, X.S. Bone-protective effects of bioactive fractions and ingredients in Sambucus williamsii Hance. Br. J. Nutr. 2011, 106, 1802–1809. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.X.; Yao, X.S.; Xiao, H.H.; Dai, Y. Extraction method of active fraction from Chinese medicine Sambucus and its application for reducing risk of menopausal bone-related diseases. Chin. Patent CN101773490, 14 July 2010. [Google Scholar]
- Xiao, H.H.; Dai, Y.; Wong, M.S.; Yao, X.S. New lignans from the bioactive fraction of Sambucus williamsii Hance and proliferation actvities on osteoblast-like UMR 106 cells. Fitoterapia 2014, 94, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.H.; Gao, Q.G.; Ho, M.X.; Zhang, Y.; Wong, K.C.; Dai, Y.; Yao, X.S.; Wong, M.S. An 8-O-4′ norlignan exerts oestrogen-like actions in osteoblastic cells via rapid nongenomic ER signaling pathway. J. Ethnopharmacol. 2015, 170, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.H.; Gao, Q.G.; Zhang, Y.; Wong, K.C.; Dai, Y.; Yao, X.S.; Wong, M.S. Vanillic acid exerts oestrogen-like activities in osteoblast-like UMR 106 cells through MAP kinase (MEK/ERK)-mediated ER signaling pathway. J. Steroid Biochem. Mol. Biol. 2014, 144(Part B), 382–391. [Google Scholar] [CrossRef] [PubMed]
- Joo, S.S.; Won, T.J.; Kang, H.C.; Lee, D.I. Isoflavones extracted from Sophorae fructus upregulate IGF-1 and TGF-β and inihbit osteoclastogenesis in rat bone marrow cells. Arch. Pharm. Res. 2004, 27, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.L.; Sun, J.Y.; Wang, D.N.; Xie, Y.H.; Wang, S.W.; Zhao, W.M. Pharmacological studies of the large-scale purified genistein from Huaijiao (Sophoara japonica–Leguminosae) on anti-osteoporosis. Phytomedicine 2006, 13, 718–723. [Google Scholar] [CrossRef] [PubMed]
- Yoon, H.J.; Seo, C.R.; Kim, M.; Kim, Y.J.; Song, N.J.; Jang, W.S.; Kim, B.J.; Lee, J.; Hong, J.W.; Nho, C.W.; et al. Dichloromethane extracts of Sophora japonica L. stimulate osteoblast differentiation in mesenchymal stem cells. Nutr. Res. 2013, 33, 1053–1062. [Google Scholar] [CrossRef] [PubMed]
- Chiou, W.F.; Lee, C.H.; Liao, J.F.; Chen, C.C. 8-Prenylkaempferol accelerates osteoblast maturation through bone morphogenetic protein-2/p38 pathway to activate RUNX2 transcription. Life Sci. 2011, 88, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, H.M.; Al-Abd, A.M.; Asaad, G.F.; Abdel-Naim, A.B.; El-halawany, A.M. Isolation of antiosteoporotic compounds from seeds of Sophora japonica. PLoS ONE 2014, 9, e98559. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Kim, J.Y.; Kim, J.J.; Oh, J.; Kim, Y.C.; Lee, M.S. (2S)-2′-Methoxykurarinone inhibits osteoclastogenesis and bone resorption through down-regulation of RANKL signaling. Biol. Pharm. Bull. 2014, 37, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Han, N.; Xu, X.; Liu, Z.; Zhang, B.; Kadota, S. Inhibitory activity of the ethyl acetate fraction from Viscum coloratum on bone resoprtion. Planta Med. 2008, 74, 120–125. [Google Scholar] [CrossRef] [PubMed]
- Han, N.; Huang, T.; Wang, Y.C.; Yin, Y.; Kadota, S. Flavanone glycosides from Viscum coloratum and their inhibitory effects on osteoclast formation. Chem. Biodiv. 2011, 8, 1682–1688. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Tezuka, Y.; Kouda, K.; Tran, Q.L.; Miyahara, T.; Chen, Y.; Kadota, S. Antiosteoporotic activity of the water extract of Dioscorea spongiosa. Biol. Pharm. Bull. 2004, 27, 583–586. [Google Scholar] [CrossRef] [PubMed]
- Peng, K.Y.; Horng, L.Y.; Sung, H.C.; Huang, H.C.; Wu, R.T. Antiosteoporotic activity of Dioscorea alata L. cv. Phyto through driving mesenchymal stem cell differentiation for bone formation. Evid. Based Complement. Altern. Med. 2011, 712892. [Google Scholar] [CrossRef]
- Kim, S.; Shin, M.Y.; Son, K.H.; Sohn, H.Y.; Lim, J.H.; Lee, J.H.; Kwun, I.S. Yam (Dioscorea batatas) root and bark extracts stimulate osteoblast mineralization by increasing Ca and P accumulation and alkaline phosphatase activity. Prev. Nutr. Food Sci. 2014, 19, 194–203. [Google Scholar] [CrossRef] [PubMed]
- Alcantara, E.H.; Shin, M.Y.; Sohn, H.Y.; Park, Y.M.; Kim, T.; J.H., L.; Jeong, H.J.; Kwon, S.T.; Kwun, I.S. Diosgenin stimulates osteogenic activity by increasing bone matrix protein synthesis and bone-specific transcription factor Runx2 in osteoblastic MC3T3-E1 cells. J. Nutr. Biochem. 2011, 22, 1055–1063. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Kouda, K.; Tezuka, Y.; LeTran, Q.; Miyahara, T.; Chen, Y.; Kadota, S. New diarylheptanoids from the rhizomes of Dioscorea spongiosa and their antiosteoporotic activity. Planta Med. 2004, 70, 54–58. [Google Scholar] [PubMed]
- Yin, J.; Han, N.; Liu, Z.; Song, S.; Kadota, S. The in vitro antiosteoporotic activity of some glycosides in Dioscorea spongiosa. Biol. Pharm. Bull. 2010, 33, 316–320. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Yang, J.; He, X.; Yang, Z.; Ding, Y.; Zhao, P.; Liu, Z.; Shao, H.; Li, Z.; Zhang, Y.; et al. Identification of upregulators of BMP-2 expression via high-throughput screening of a synthetic and natural compound library. J. Biomol. Screen. 2009, 14, 1251–1256. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.H.; Li, Q.; Wu, M.Y.; Guo, D.J.; Chen, H.L.; Chen, S.L.; Seto, S.W.; Au, A.L.S.; Poon, C.C.W.; Leung, G.P.H.; et al. Formononetin, an isoflavone, relaxes rat isolated aorta through endothelium-dependent and endothelium-indepenent pathways. J. Nutr. Biochem. 2010, 21, 613–620. [Google Scholar] [CrossRef] [PubMed]
- Ha, H.; Lee, H.Y.; Lee, J.H.; Jung, D.; Choi, J.; Song, K.Y.; Jung, H.J.; Choi, J.S.; Chang, S.I.; Kim, C. Formononetin prevents ovariectomy-induced bone loss in rats. Arch. Pharm. Res. 2010, 33, 625–632. [Google Scholar] [CrossRef] [PubMed]
- Gautam, A.K.; Bhargavan, B.; Tyagi, A.M.; Srivastava, K.; Yadav, D.K.; Kumar, A.; Singh, A.; Mishra, J.S.; Singh, A.B.; Sanyal, S.; et al. Differential effects of formononetin and cladrin on osteoblast function, peak bone mass achievement and bioavailability in rats. J. Nutr. Biochem. 2011, 22, 318–327. [Google Scholar] [CrossRef] [PubMed]
- Kaczmarczyk-Sedlak, I.; Wojnar, W.; Zych, M.; Ozimina-Kaminska, E.; Taranowicz, J.; Siwek, A. Effect of formononetin on mechanical properties and chemical composition of bones in rats with ovariectomy-induced osteoporosis. Evid. Based Complement. Altern. Med. 2013, 457052. [Google Scholar] [CrossRef] [PubMed]
- Rong, H.S.; Jan, F.; Deinzer, M.L.; Cooman, L.D.; Keukeleire, D.D. Identification of isoflavones in the roots of Pueraria lobata. Planta Med. 1998, 64, 620–627. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.; Hao, J.; Xu, Z.; Chen, T.; Qiu, S.X. Polyprenylated isoflavone and isoflavonoids from Ormosia henryi and their cytotoxicity and anti-oxidation activity. Fitoterapia 2012, 83, 161–165. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.H.; Wang, R.; Shi, Y.P. Flavonoids in the poisonous plant Oxytropis falcata. J. Nat. Prod. 2010, 73, 1398–1403. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, K.; Tyagi, A.M.; Khan, K.; Dixit, M.; Lahiri, S.; Kumar, A.; Changkija, B.; Khan, M.P.; Nagar, G.K.; Yadav, D.K.; et al. Isoformononetin, a methoxydaidzein present in medicinal plants, reverses bone loss in osteopenic rats and exerts bone anabolic action by preventing osteoblast apoptosis. Phytomedicine 2013, 20, 470–480. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhao, L.; Liu, X.; Qi, J. Effects of IBP on calcium absorption and biochemical indicators in glucocorticoid-induced osteoporosis rats. Chin. J. Prev. Med. (Zhongguo Yufang Yixue Zazhi) 2011, 12, 1004–1007. [Google Scholar]
- Park, E.; Jin, H.S.; Cho, D.Y.; Kim, J.; Kim, M.C.; Choi, C.W.; Jin, Y.; Lee, J.W.; Park, J.H.; Chung, Y.S.; et al. The effect of lycii Radicis Cortex extract on bone formation in vitro and in vivo. Molecules 2014, 19, 19594–19609. [Google Scholar] [CrossRef] [PubMed]
- Yen, P.H.; Kiem, P.V.; Nhiem, N.X.; Tung, N.H.; Quang, T.H.; Minh, C.V.; Kim, J.W.; Choi, E.M.; Kim, Y.H. A new monoterpene glycoside from the roots of Paeonia lactiflora increases the differentiation of osteoblastic MC3T3-E1 cells. Arch. Pharm. Res. 2007, 30, 1179–1185. [Google Scholar] [CrossRef] [PubMed]
- Appendino, G.; Spagliardi, P.; Carvotto, G.; Pocock, V.; Milligan, S. Daucane phytoestrogens: A structure-activity study. J. Nat. Prod. 2002, 65, 1612–1615. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, K.; arao, Y.; Otsuka, H.; Nomoto, S.; Horiguchi, H.; Kato, S.; Kayama, F. Terpenoids found in the Umbelliferae family act as agonists/antagonists for ERα and ERβ: Differential transcription activity between ferutinine-liganded ERα and ERβ. Biochem. Biophys. Res. Commun. 2002, 291, 354–360. [Google Scholar] [CrossRef] [PubMed]
- Appendino, G.; Spagliardi, P.; Sterner, O.; Milligan, S. Structure-activity relationships of the estrogenic sesquiterpene ester ferutinin. Modification of the terpenoid core. J. Nat. Prod. 2004, 67, 1557–1564. [Google Scholar] [CrossRef] [PubMed]
- Ferretti, M.; Bertoni, L.; Cavani, F.; Zavatti, M.; Resca, E.; Carnevale, G.; Benelli, A.; Zanoli, P.; Palumbo, C. Influence of ferutinin on bone metabolism in ovariectomized rats. II: Role in recovering osteoporosis. J. Anat. 2010, 217, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Palumbo, C.; Ferretti, M.; Bertoni, L.; Cavani, F.; Resca, E.; Casolari, B.; Carnevale, G.; Zavatti, M.; Montanari, C.; Benelli, A.; et al. Influence of ferutinin on bone metabolism in ovariectomized rats. I: Role in preventing osteoporosis. J. Bone Miner. MeTable 2009, 27, 538–545. [Google Scholar] [CrossRef] [PubMed]
- Cavani, F.; Ferretti, M.; Carnevale, G.; Bertoni, L.; Zavatti, M.; Palumbo, C. Effects of different doses of ferutinin on bone formation/resorption in ovariectomized rats. J. Bone Miner. Metab. 2012, 30, 619–629. [Google Scholar] [CrossRef] [PubMed]
- Ferretti, M.; Bertoni, L.; Cavani, F.; Benincasa, M.; Sena, P.; Carnevale, G.; Zavatti, M.; Viesti, V.D.; Zanoli, P.; Palumbo, C. Structural and histomorphometric evaluations of ferutinin effects on the uterus of ovariectomized rats during osteoporosis treatment. Life Sci. 2012, 90, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Ferretti, M.; Cavani, F.; Manni, P.; Carnevale, G.; Bertoni, L.; Zavatti, M.; Palumbo, C. Ferutinin dose-dependent effects on uterus and mammary gland in ovariectomized rats. Histol. Histopathol. 2014, 29, 1027–1037. [Google Scholar] [PubMed]
- Zavatti, M.; Resca, E.; Bertoni, L.; Maraldi, t.; Guida, M.; Carnevale, G.; Ferrari, A.; DePol, A. Ferutinin promotes proliferation and osteoblastic differentiation in human amniotic fluid and dental pulp stem cells. Life Sci. 2013, 92, 993–1003. [Google Scholar] [CrossRef] [PubMed]
- Zavatti, M.; Bertoni, L.; Maraldi, T.; Resca, E.; Beretti, F.; Guida, M.; LaSala, G.B.; DePol, A. Critical-size bone defect repair using amniotic fluid stem cell/collagen constructs: Effect of oral ferutinin treatment in rats. Life Sci. 2015, 121, 174–183. [Google Scholar] [CrossRef] [PubMed]
- Park, E.J.; Pezzuto, J.M. The pharmacology of resveratrol in animals and humans. Biochim. Biophys. Acta 2015, 1852, 1071–1113. [Google Scholar] [CrossRef] [PubMed]
- Mobasheri, A.; Shakibaei, M. Osteogenic effects of resveratrol in vitro: Potential for the prevention and treatment of osteoporosis. Ann. N. Y. Acad. Sci. 2013, 1290, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Tou, J.C. Evaluating resveratrol as a therapeutic bone agent: Preclinical evidence from rat models of osteoporosis. Ann. N. Y. Acad. Sci. 2015, 1348, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Mizutani, K.; Ikeda, K.; Kawai, Y.; Yamori, Y. Resveratrol stimulates the proliferation and differentiation of osteoblastic MC3T3-E1 cells. Biochem. Biophys. Res. Commun. 1998, 253, 859–863. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.; Li, Y.; Quarles, L.D.; Song, T.; Pan, W.; Zhou, H.; Xiao, Z. Resveratrol enhances proliferation and osteoblastic differentiation in human mesenchymal stem cells via ER-dependent ERK1/2 activation. Phytomedicine 2007, 14, 806–814. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Shang, L.; Li, X.; Zhang, X.; Gao, G.; Guo, C.; Chen, B.; Liu, Q.; Gong, Y.; Shao, C. Resveratrol augments the canonical Wnt signaling pathway in promoting osteoblastic differentiation of multipotent mesenchymal cells. Exp. Cell Res. 2009, 315, 2953–2962. [Google Scholar] [CrossRef] [PubMed]
- Tseng, P.C.; Hou, S.M.; Chen, R.J.; Peng, H.W.; Hsieh, C.F.; Kuo, M.L.; Yen, M.L. Resveratrol promotes osteogenesis of human mesenchymal stem cells by upregulating RUNX2 gene expression via the SIRT1/FOXO3A axis. J. Bone Miner. Res. 2011, 26, 2552–2563. [Google Scholar] [CrossRef] [PubMed]
- Shakibaei, M.; Shayan, P.; Busch, F.; Aldinger, C.; Buhrmann, C.; Lueders, C.; Mobasheri, A. Resveratrol mediated modulation of SIRT-1/RUNX2 promotes osteogenic differentiation of mesenchymal stem cells: Potential role of RUNX2 deacetylation. PLoS ONE 2012, 7, e35712. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Andersson, G.; Lindgren, U.; Li, Y. Resveratrol prevents RANKL-induced osteoclast differentiation of murine osteoclast progenitor RAW264.7 cells through inhibition of ROS production. Biochem. Biophys. Res. Commun. 2010, 401, 356–362. [Google Scholar] [CrossRef] [PubMed]
- Shakibaei, M.; Buhrmann, C.; Mobasheri, A. Resveratrol-mediated SIRT-1 interactions with p300 modulate receptor activator of NF-κB ligand (RANKL) activation of NF-κB signaling and inhibit osteoclastogenesis in bone-derived cells. J. Biol. Chem. 2011, 286, 11492–11505. [Google Scholar] [CrossRef] [PubMed]
- Tou, J.C. Resveratrol supplementation affects bone acquisition and osteoporosis: Pre-clinical evidence toward translational diet therapy. Biochim. Biophys. Acta 2015, 1852, 1186–1194. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.P.; Li, W.X.; Yu, B.; Huang, J.; Sun, J.; Huo, J.S.; Liu, C.X. Effects of trans-resveratrol from Polygonum cuspidatum on bone loss using the ovariectomized rat model. J. Med. Food 2005, 8, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Li, X.; Li, N.; Liu, T.; Liu, J.; Li, Z.; Xiao, H.; Li, J. Long-term resveratrol treatment prevents ovariectomy-induced osteopenia in rats without hyperplastic effects on the uterus. Br. J. Nutr. 2014, 111, 836–846. [Google Scholar] [CrossRef] [PubMed]
- Habold, C.; Momken, I.; Ouadi, A.; Bekaert, V.; Brasse, D. Effect of prior treatment with resveratrol on density and structure of rat long bones under tail-suspension. J. Bone Miner. Metab. 2011, 29, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Durbin, S.M.; Jackson, J.R.; Tyan, M.J.; Gigliotti, J.C.; Alway, S.E.; Tou, J.C. Resveratrol supplementation influences bone properties in the tibia of hindlimb-suspended mature Fisher 344 x Brown Norway male rats. Appl. Physiol. Nutr. MeTable 2012, 37, 1179–1188. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.D.; Shi, Y.M.; Li, L.; Guo, J.D.; Zhang, Y.P.; Hou, S.X. Treatment with resveratrol attenuates sublesional bone loss in spinal cord-injured rats. Br. J. Pharmacol. 2013, 170, 796–806. [Google Scholar] [CrossRef] [PubMed]
- Casarin, R.C.; Casati, M.Z.; Pimentel, S.P.; Cirano, F.R.; Algayer, M.; Pires, P.R.; Ghiraldini, B.; Duarte, P.M.; Ribeiro, F.V. Resveratrol improves bone repair by moduation of bone morphogenetic proteins and osteopontin gene expression in rats. Int. J. Oral Maxillofac. Surg. 2014, 43, 900–906. [Google Scholar] [CrossRef] [PubMed]
- Tresguerres, I.F.; Tamimi, F.; Eimar, H.; Barralet, J.; Torres, J.; Blanco, L.; Tresguerres, J.A.F. Resveratrol as anti-aging therapy for age-related bone loss. Rejuvenation Res. 2014, 17, 439–445. [Google Scholar] [CrossRef] [PubMed]
- Durbin, S.M.; Jackson, J.R.; Ryan, M.J.; Gigliotti, J.C.; Alway, S.E.; Tou, J.C. Resveratrol supplementation preserves long bone mass, microstructure, and strength in hindlimb-suspended old male rats. J. Bone Miner. Metab. 2014, 32, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Danmark, S.; Edlund, U.; Finne-Wistrand, A.; He, X.; Norgard, M.; Blomen, E.; Hutenby, K.; Andersson, G.; Lindgren, U. Resveratrol-conjugated poly-ε-caprolactone facilitates in vitro mineralization and in vivo bone regeneration. Acta Biomater. 2011, 7, 751–758. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.; Wu, T.; Li, Q.N.; Lin, L.S.; Liang, N.C. Preventive effects of ginsenosides on osteopenia of rats induced by ovariectomy. Acta Pharmacol. Sin. 2001, 22, 428–434. [Google Scholar] [PubMed]
- Gong, Y.S.; Chen, J.; Zhang, Q.Z.; Zhang, J.T. Effects of 17-β-oestradiol and ginsenoside on osteoporosis in ovariectomized rats. J. Asian Nat. Prod. Res. 2006, 8, 649–656. [Google Scholar] [CrossRef] [PubMed]
- Cheng, B.; Li, J.; Du, J.; Lv, X.; Weng, L.; Ling, C. Ginsenoside Rb1 inhibits osteoclastogenesis by modulating NF-κB and MAPKS pathways. Food Chem. Toxicol. 2012, 50, 1610–1615. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Gao, B.; Jie, Q.; Wei, B.Y.; Fan, J.; Zhang, H.Y.; Zhang, J.K.; Li, X.J.; Shi, J.; Luo, Z.J.; et al. Ginsenoside-Rb2 displays anti-osteoporosis effects through reducing oxidative damage and bone-resorbing cytokines during osteogenesis. Bone 2014, 66, 306–314. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Huang, Q.; Jie, Q.; Zhang, H.Y.; Wang, L.; Guo, Y.S.; Sun, Z.; Wei, B.Y.; Han, Y.H.; Liu, J.; et al. Ginsenoside-Rb2 inhibits dexamethasone-induced apoptosis through promotion of GPR120 induction in bone marrow-derived mesenchymal stem cells. Stem Cells Dev. 2015, 24, 781–790. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.Y.; Park, Y.G.; Quan, H.Y.; Kim, S.J.; Jung, M.S.; Chung, S.H. Ginsenoside Rd stimulates the differentiation and mineralization of osteoblastic MC3T3-E1 cells by activating AMP-activated protein kinase via the BMP-2 signaling pathway. Fitoterapia 2012, 83, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.H.; Cheng, W.X.; Qin, Z.S.; Sun, K.M.; Zhong, M.; Wang, J.K.; Gao, W.Y.; Yu, Z.H. Effects of ginsenoside Rg1 on the proliferation and osteogenic differentiation of human periodontal ligament stem cells. Chin. J. Integr. Med. 2015, 21, 676–681. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Wei, X.; Zhang, F.; Yang, K.; Qu, C.; Luo, H.; He, L. Ginsenoside Rg1 of Panax ginseng stimulates the proliferation, odontogenic/osteogenic differentiation and gene expression profiles of human dental pulp stem cells. Phytomedicine 2014, 21, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.F.; Gao, Q.G.; Dai, Z.J.; Kung, A.W.; Guo, D.A.; Wong, M.S. Estrogenic effects of ginsenoside Rg1 in endometrial cells in vitro were not observed in immature CD-1 mice or ovariectomized mice model. Menopause 2012, 19, 1052–1061. [Google Scholar] [CrossRef] [PubMed]
- Siddiqi, M.H.; Siddiqi, M.Z.; Kang, S.; Noh, H.Y.; Ahn, S.; Simu, S.Y.; Aziz, M.A.; Sathishkumar, N.; Perez, Z.E.J.; Yang, D.C. Inhibition of osteoclast differentiation by ginsenoside Rg3 in RAW264.7 cells via RANKL, JNK and p38 MAPK pathways through a modulation of cathepsin K: An in silico and in vitro study. Phytother. Res. 2015, 29, 1286–1296. [Google Scholar] [CrossRef] [PubMed]
- Siddiqi, M.Z.; Siddiqi, M.H.; Kim, Y.J.; Jin, Y.; Huq, M.A.; Yang, D.C. Effect of fermented red ginseng extract enriched in ginsenoside Rg3 on the differentiation and mineralization of preosteoblastic MC3T3-E1 cells. J. Med. Food 2015, 18, 542–548. [Google Scholar] [CrossRef] [PubMed]
- Siddiqi, M.H.; Siddiqi, M.Z.; Ahn, S.; Kang, S.; Kim, Y.J.; Veerappan, K.; Yang, D.U.; Yang, D.C. Stimulative effect of ginsenosides Rg5:Rk1 on murine osteoblastic MC3T3-E1 cells. Phytother. Res. 2014, 28, 1447–1455. [Google Scholar] [CrossRef] [PubMed]
- Siddiqi, M.H.; Siddiqi, M.Z.; Ahn, S.; Kim, Y.J.; Yang, D.C. Ginsenoside Rh1 induces mouse osteoblast growth and differentiation through the bone morphogenetic protein 2/RUNT-related gene 2 signalling pathway. J. Pharm. Pharmacol. 2014, 66, 1763–1773. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Lee, J.; Jang, J.H.; Lee, S.H.; Nan, M.H.; Oh, B.C.; Lee, S.G.; Kim, H.H.; Soung, N.K.; Ahn, J.S.; et al. Ginsenoside Rh2 inhibits osteoclastogenesis through down-regulation of NF-κB, NFATc1 and c-Fos. Bone 2012, 50, 1207–1213. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Shiono, J.; Shimizu, K.; Yu, H.; Zhang, C.; Jin, F.; Kondo, R. 20(R)-Ginsenoside Rh2, not 20(S), is a selective osteoclastogenesis inhibitor without any cytotoxicity. Bioorg. Med. Chem. Lett. 2009, 19, 3320–3323. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.Y.; Jung, M.S.; Park, Y.G.; Yuan, H.D.; Quan, H.Y.; Chung, S.H. Ginsenosdie Rh2(S) induces the differentiation and mineralization of osteoblastic MC3T3-E1 cells through activation of PKD and p38 MAPK pathways. BMB Rep. 2011, 44, 659–664. [Google Scholar]
- Kim, D.Y.; Park, K.H.; Jung, M.S.; Huang, B.; Yuan, H.D.; Quan, H.Y.; Chung, S.H. Gensenoside Rh2(S) induces differentiation and mineralization of MC3T3-E1 cells through activation of the PKD/AMPK signaling pathways. Int. J. Mol. Med. 2011, 28, 753–759. [Google Scholar]








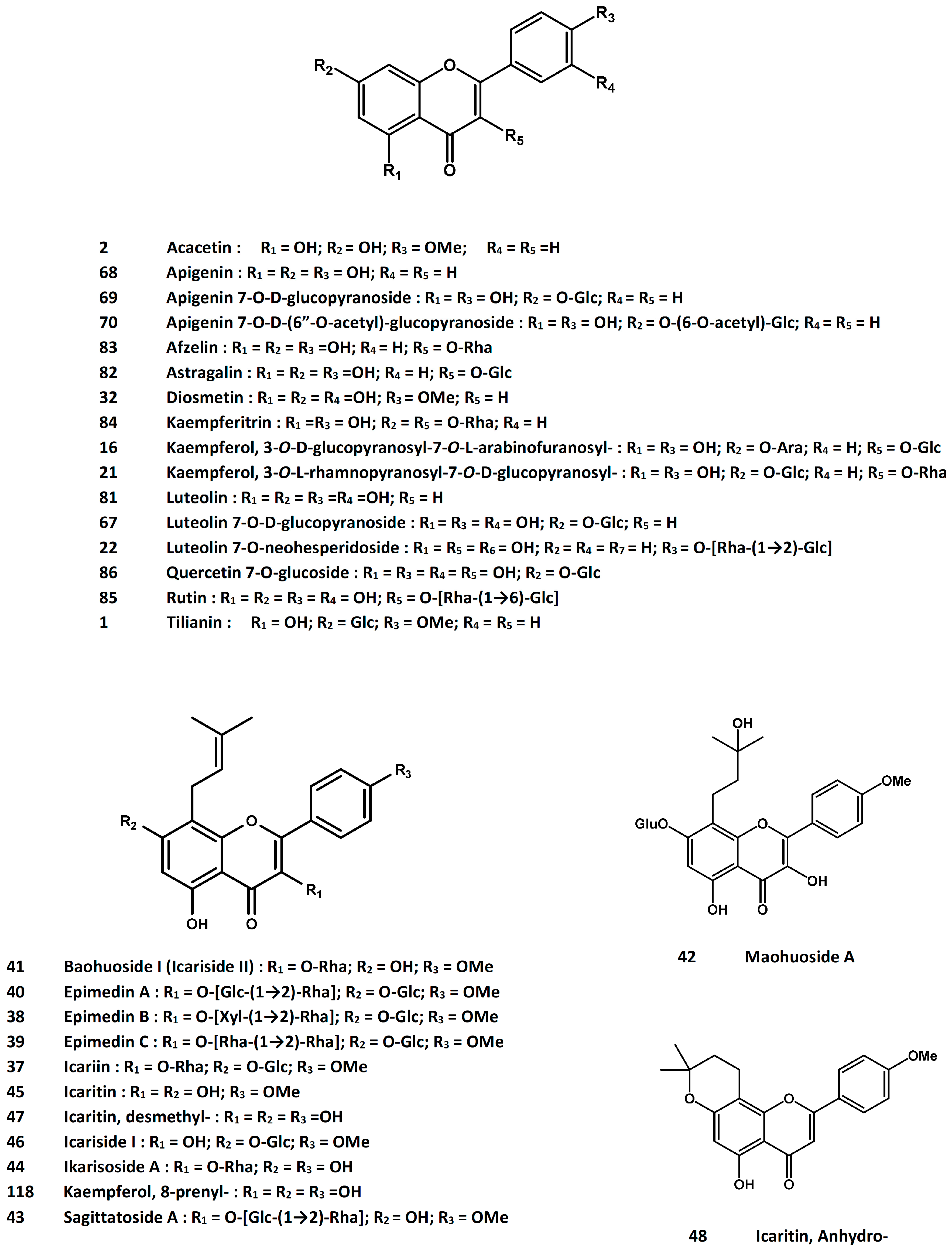{kind=link}
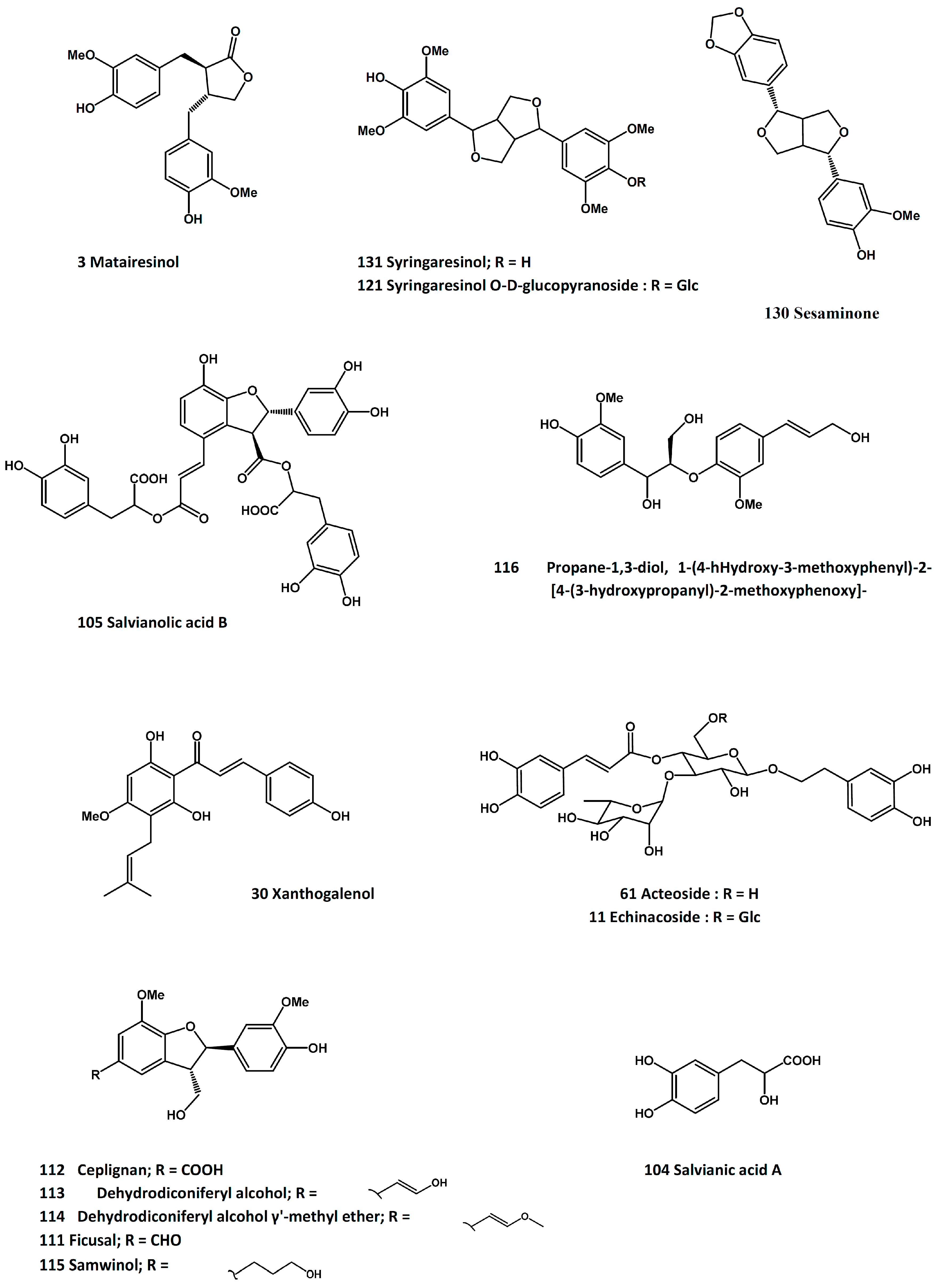{kind=link}
{kind=link}
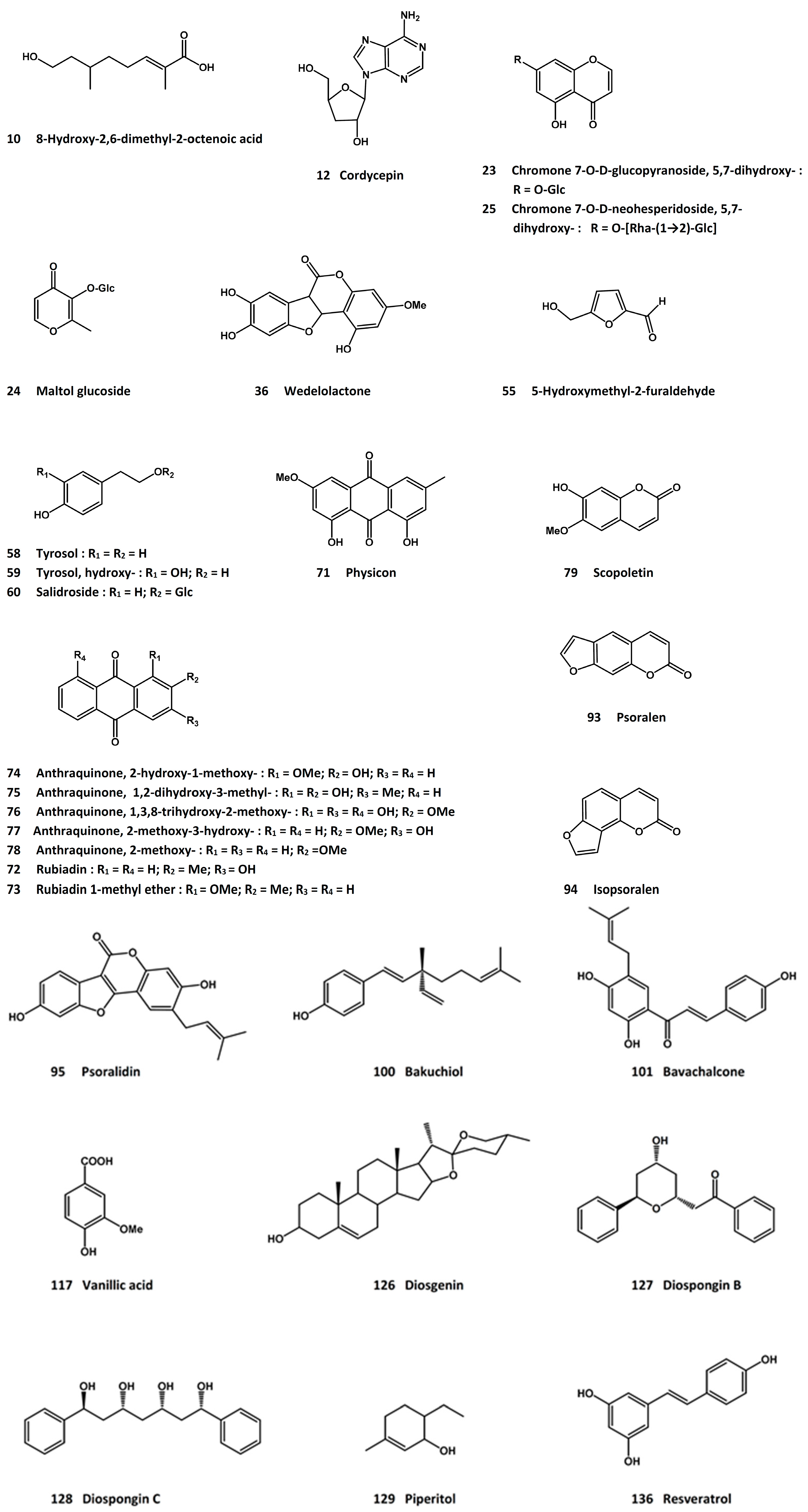{kind=link}
{kind=link}
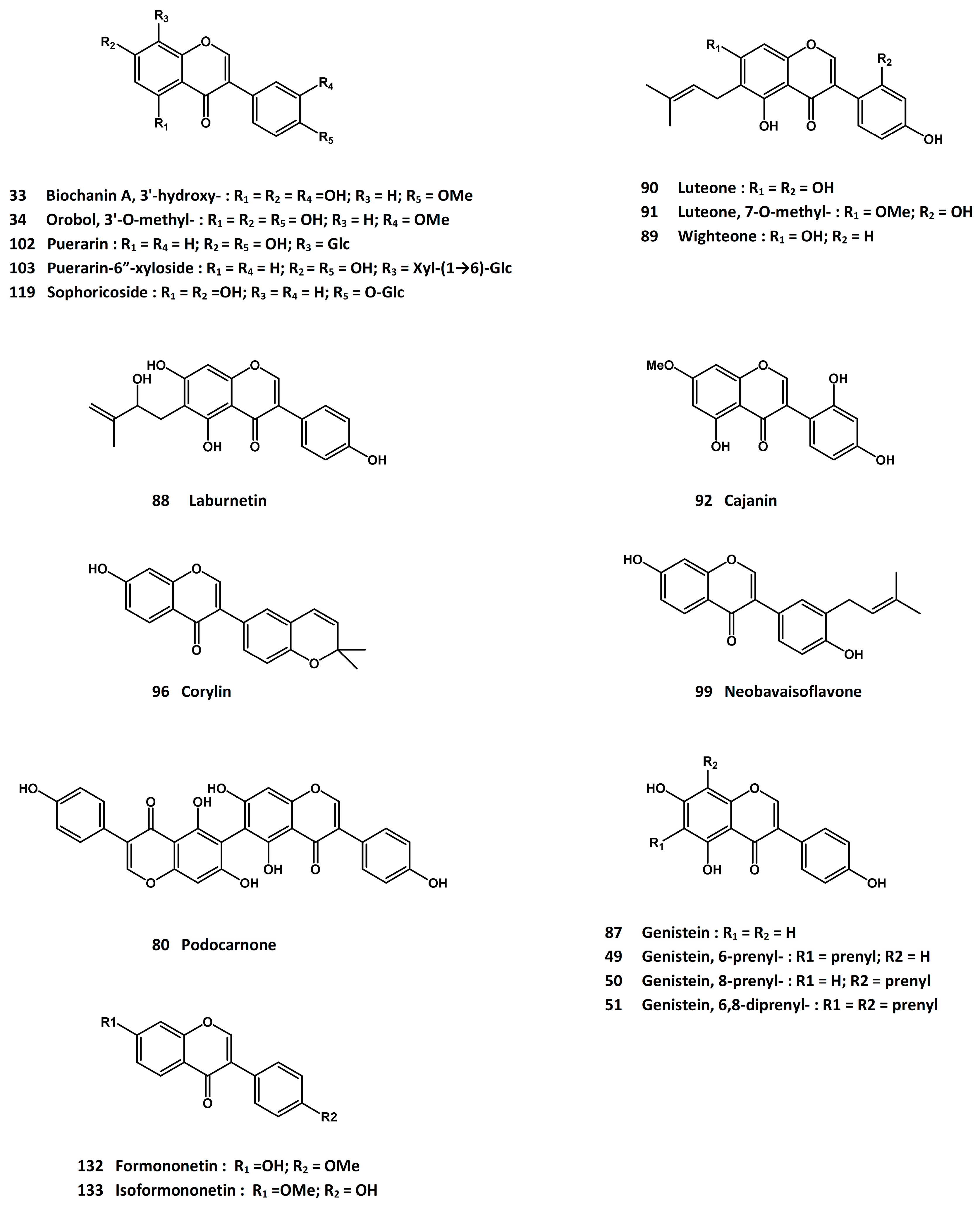{kind=link}
{kind=link}
| Bone Formation | Bone Resorption |
|---|---|
| Osteoblast enzyme | Osteoclast enzyme |
| Total alkaline phosphatase [52] | Tartrate-resistant acid phosphatase (TRAP) [54] |
| Bone-specific alkaline phosphatase [53] | Cathepsin K [55] |
| Matrix protein | Collagen degradation product |
| Osteocalcin [56,57] | Hydroxyproline [57] |
| Pyridinoline [58] | |
| Deoxypyridinoline [58] | |
| Product of collagen synthesis | Cross-linked telopeptide of type I collagen |
| Procollagen type I C-terminal extension peptide (P1CP) [59] | N-telopeptide of type I collagen [59] |
| Procollagen type I N-terminal extension peptide (P1NP) [59] | C-telopeptide of type I collagen [59] |
| C-telopeptide generated by matrix metalloprotieinases [59] | |
| Others | |
| Osteopontin (OPN) [60] | |
| Receptor activator of nuclear factor Kappa B ligand (RANKL) [61] | |
| Osteoprotegerin (OPG) [62] |
| Latin Name | Chinese Name | Plant Part | Major Active Molecule |
|---|---|---|---|
| Actaea heracleifolia (Kom.) J. Compton; A. dahurica (Turcz. ex Fisch. & C.A. Mey.) Franch.; A. foetida L. | Sheng-Ma | Rhizome | cimicidol-3-O-β-xyloside, cimicidanol-3-O-β-xyloside, acetylacteol-3-O-arabinoside [73] |
| Astragalus mongholicus Bunge | Huang-Qi | Root | formononetin [355,356,357,358,359] |
| Carthamus tinctorius L. | Hong-Hua | Flower | matairesinol, tilianine, acacetin and their derivatives [72] |
| Cistanche deserticola Y.C. Ma; C. tubulosa (Forssk.) Beck | Rou-Cong-Rong | Stem | 8-hydroxy-2,6-dimethyl-2-octenoic acid [81], echinacoside [85,86,87] |
| Cordyceps sinensis (Berk.) Sacc. | Dong-Cong-Xia-Cao | cordycepin [94,95,96,97] | |
| Dioscorea spp. | Rhizome | diosgenin [353], diospongins B and C, piperitol, sesqminone, syrinaresinol [353] | |
| Dipsacus asper Wall. ex C.B. Clarke; D. japonicas Miq. | Xu-Duan | Root | asperosaponins V and VI [103,106], hederagenin-3-O-(2-O-acetyl)-α-l-arabinopyranoside [104] |
| Drynaria fortunei (Kunze ex Mett.) J. Sm. | Gu-Sui-Bu | Rhizome | naringin and other flavos [116,117,118,119,120,121,122,123,124,125,126,127,128,129,130,131,132,133,134,135,136,137,138,139,140,141,142,143,144,145,146,147] |
| Eclipta prostrata L. | Mo-Han-Lian | Above-ground parts | diosmetin, 3′-hydroxybiochanin A, 3′-O-methylorobol [150], echinocystic acid [156], wedelolactone [157] |
| Epimedium brevicornum Maxim.; E. sagittatum (Siebold & Zucc.) Maxim.; E. pubescens Maxim.; E. koreanum Nakai | Yin-Yang-Huo | Leaf | icariin [158,159,160,161,162,163,164,165,166,167,168,169,170,171,172,173,174,175,176,177,178,179,180,181,182,183,184,185,186,187,188,189,190,191,192,193,194,195,196,197,198,199,200,201,202,203,204,205,206,207,208,209,210], epimedins A, B and C [178,211], baohuoside-1 [189,212,213], maohuoside A [214,215], sagittatoside A [189], ikarisoside A [216], icaritin [209,220,221,222,223,224,225,226,227,228,229,230], icariside I and II [209,219] |
| Erythina variegate L. | Hai-Tong-Pi | Bark | 6-prenylgenistein, 8-prenylgenistein, 6,8-diprenylgenistein [233] |
| Eucommia ulmoides Oliv. | Du-Zhong | Stem bark | geniposidic acid, geniposide, aucubin [237,238], 5-hydroxymethyl-2-furaldehyde [239] |
| Ferula spp. | A-Wei | Resin | Ferutinin [370,371,372,373,374,375,376] |
| Ligustrum lucidum W.T. Aiton | Lu-Zhen-Zhi | Fruit | oleanolic acid [244,245,246,247,248,249,250], ursolic acid [250,251,252,253], tyrosol, hydroxytyrosol, oleuropein, and others [256] |
| Lycium babarum L. | Gou-Qi-zi | Fruit | polysaccharide [364] |
| Morinda officinalis F.C. How | Ba-Ji-tian | Root | physicion, rubiadin, rubiadin-1-methyl ether, 2-hydroxy-1-methoxyanthraquinone, 1,2-dihydroxy-3-methylanthraquinone, 1,3,8-trihydroxy-2-methoxyanthraquinone, 2-hydroxymethyl-3-hydroxyanthraquinone, 2-methoxyanthraquinone, scopoletin [261,262] |
| Ormosia henryi Prain | Lu-Mu | Root | isoformononetin [266,363] |
| Oxytropis falcata Bunge | Lian-Xing-Ji-Dou | Whole plant | isoformononetin [266,363] |
| Paeonia lactiflora Pall. | Bai-Shao | Root | 6′-O-β-d-glucopyranosylalbiflorin [366] |
| Panax spp. | Ren-Shen | Root | ginsenosides [397,398,399,400,401,402,403,404,405,406,407,408,409,410,411,412,413] |
| Podocarpium podocarpum (DC.) Yang et Huang | Whole plant | podocarnone, luteolin, astragalin, afzelin, kaempferitrin, rutin, quercetin-7-O-d-glucopyranoside, genistein, laburnetin, wighteone, luteone, 7-O-methyl-luteone [265], cajanin [266] | |
| Polygonum cuspidatum Siebold & Zucc. | Hu-Zhang | Root and Rhizome | resveratrol [378,379,380,381,382,383,384,385,386,387,388,389,390,391,392,393,394,395,396] |
| Psoralea corylifolia L. | Bu-Gu-Zhi | Fruit | psoralen [274,275,276], isopsoralen [278,279], psoralidin [280], corylin [281], bavachin, isobavachin [282,283], neobavaisoflavone [284], bakuchiol [283,285], bavachalcone [286] |
| Pueraria lobate (Willd.) Ohwi | Ge-Gen | Root | isoformononetin [266,363], puerarin and its 6′′-xyloside [297,298,299,300,301,302,303,304,305,306,307,308,309,310,311] |
| Rehmannia glutinosa (Gaertn.) DC. | Di-Huang | Root | acteoside [314,315,316] |
| Salvia miltiorrhiza Bunge | San-Shen | Root/rhizome | salvianolic acids A and B [322,323], tanshinones I, IIA and VI, cryptotanshinone, 15,16-dihydrotanshinone I [324,325,326,327,328,329,330,331] |
| Sambucus williamsii Hance | Jie-Gu-Mu | Stem | ficusal, ceplignan, dehydrodiconiferyl alcohol, dehydrodiconiferyl alcohol-γ′-methyl ether, samwinol [338], erythro-1-(4-hydroxy-3-methoxyphenyl)-2-[4-(3-hydroxypropanyl) -2-methoxyphenoxy]-1,3-propanediol [339], vanillic acid [340] |
| Sophora japonica L. | Huai-Jiao | Fruit, seed and root | genistein [342,343], 8-prenylkaempferol [344], sophoricoside [345], 2′-methoxykurarinone [346] |
| Sophora flavescens Aiton | Ku-Shen | Root | formononetin [355,356,357,358,359] |
| Trifolium pretense L. | Hong-Che-Hou-Cao | Inflorescence and twig | formononetin [355,356,357,358,359] |
| Viscum coloratum (Kom.) Nakai | Hu-Ji-Sheng | Twig | syringareninol O-β-glucopyranoside, 2-homoeriodictyol 7-O-β-glucopyranoside, viscumneosides I, IX and X [347,348] |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Che, C.-T.; Wong, M.S.; Lam, C.W.K. Natural Products from Chinese Medicines with Potential Benefits to Bone Health. Molecules 2016, 21, 239. https://doi.org/10.3390/molecules21030239
Che C-T, Wong MS, Lam CWK. Natural Products from Chinese Medicines with Potential Benefits to Bone Health. Molecules. 2016; 21(3):239. https://doi.org/10.3390/molecules21030239
Chicago/Turabian StyleChe, Chun-Tao, Man Sau Wong, and Christopher Wai Kei Lam. 2016. "Natural Products from Chinese Medicines with Potential Benefits to Bone Health" Molecules 21, no. 3: 239. https://doi.org/10.3390/molecules21030239
APA StyleChe, C. -T., Wong, M. S., & Lam, C. W. K. (2016). Natural Products from Chinese Medicines with Potential Benefits to Bone Health. Molecules, 21(3), 239. https://doi.org/10.3390/molecules21030239