
Nanopore Event-Transduction Signal Stabilization for Wide pH Range under Extreme Chaotrope Conditions

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. Channel Current Detectors
1.1.1. Transduction vs. Translation
1.1.2. Single-Molecule vs. Ensemble
1.1.3. Biomedicine Needs Biosensing with High Sensitivity in Presence of Interference
1.2. Nanopore Transduction Detection and Stochastic Carrier Wave Signal Analysis
1.2.1. Ubiquitous Transduction Channel-Modulator Capability via Laser Modulation
1.2.2. Antibody Binding Studies
2. Background
2.1. Pathogen Detection, miRNA Detection, and miRNA Haplotyping
2.2. Glycoassayer, Posttranslational Assayer
2.3. Enzyme studies: HIV integrase
2.4. Streptavidin Toxin Biosensor
3. Results
3.1. Biotin-Streptavidin Binding Experiments
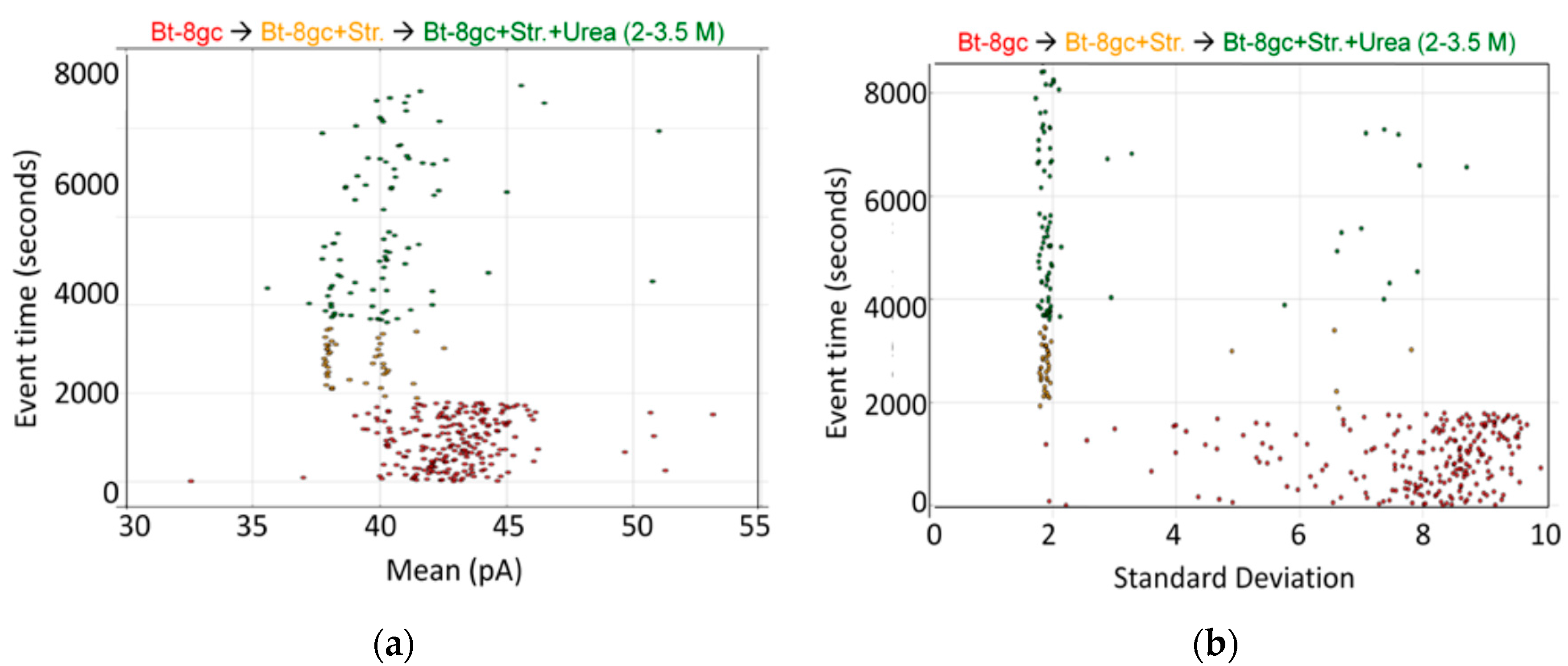
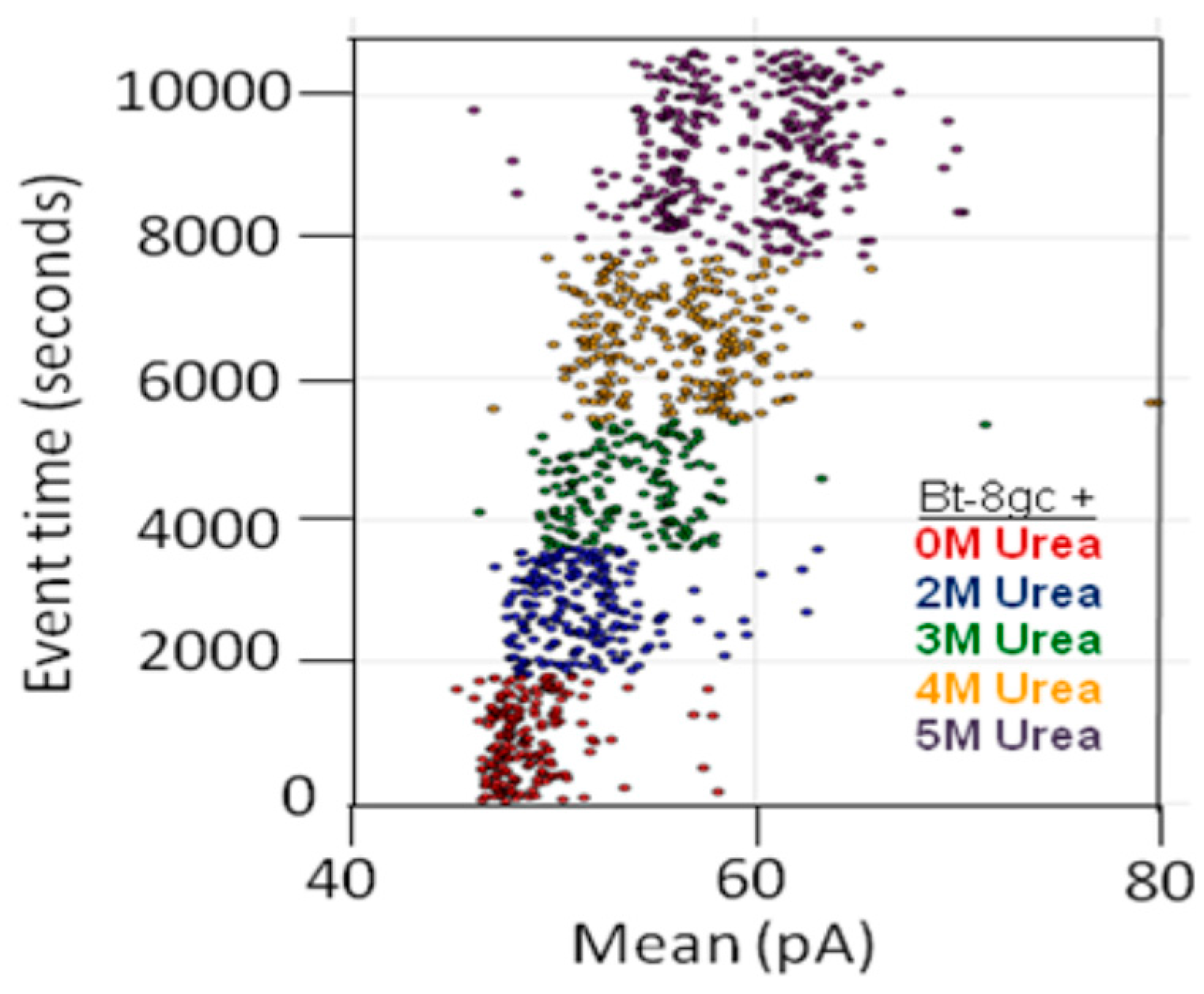
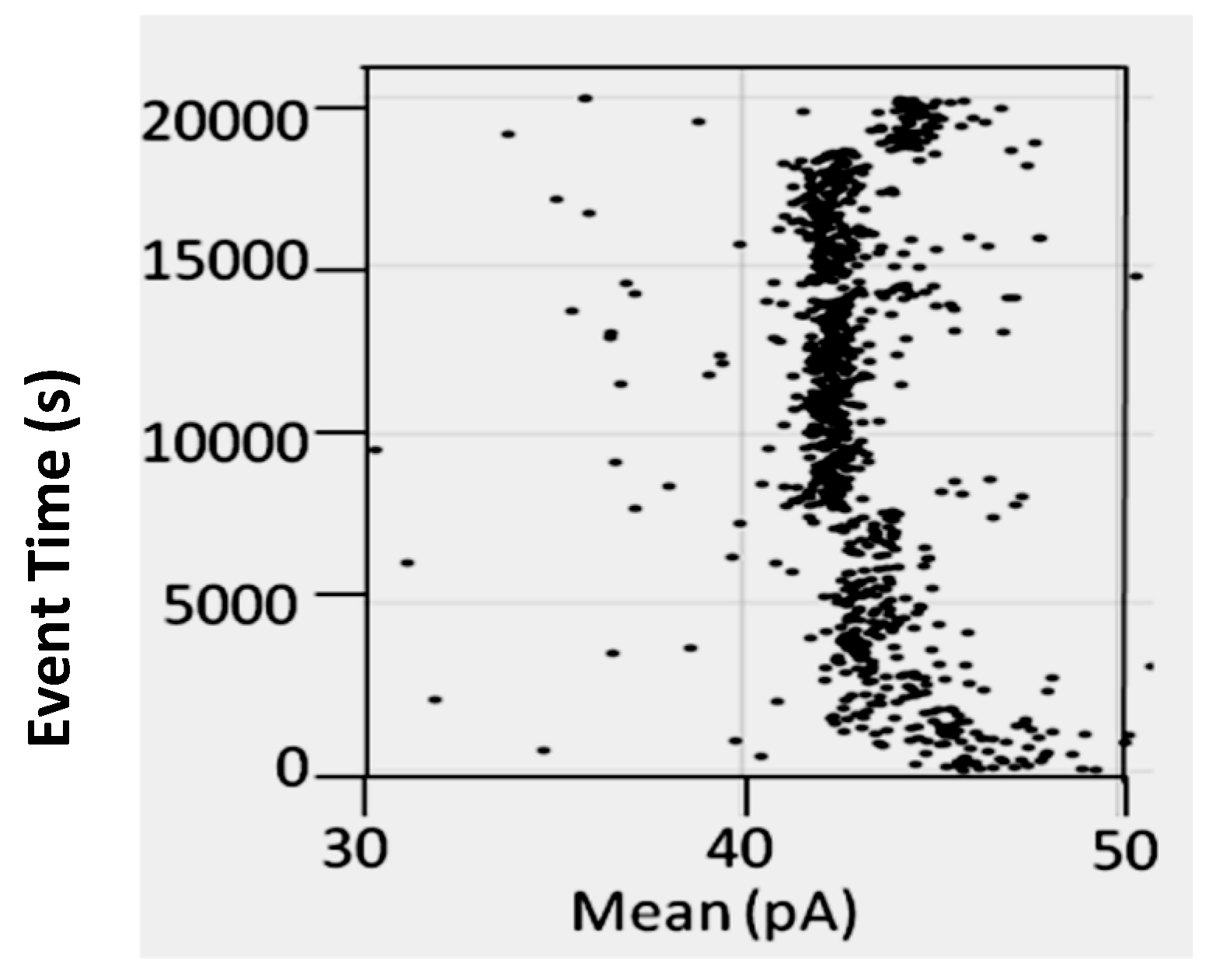
3.1.1. The BT-8gc Transducer Retains its Viability in the Presence of Urea up to 5M Concentrations
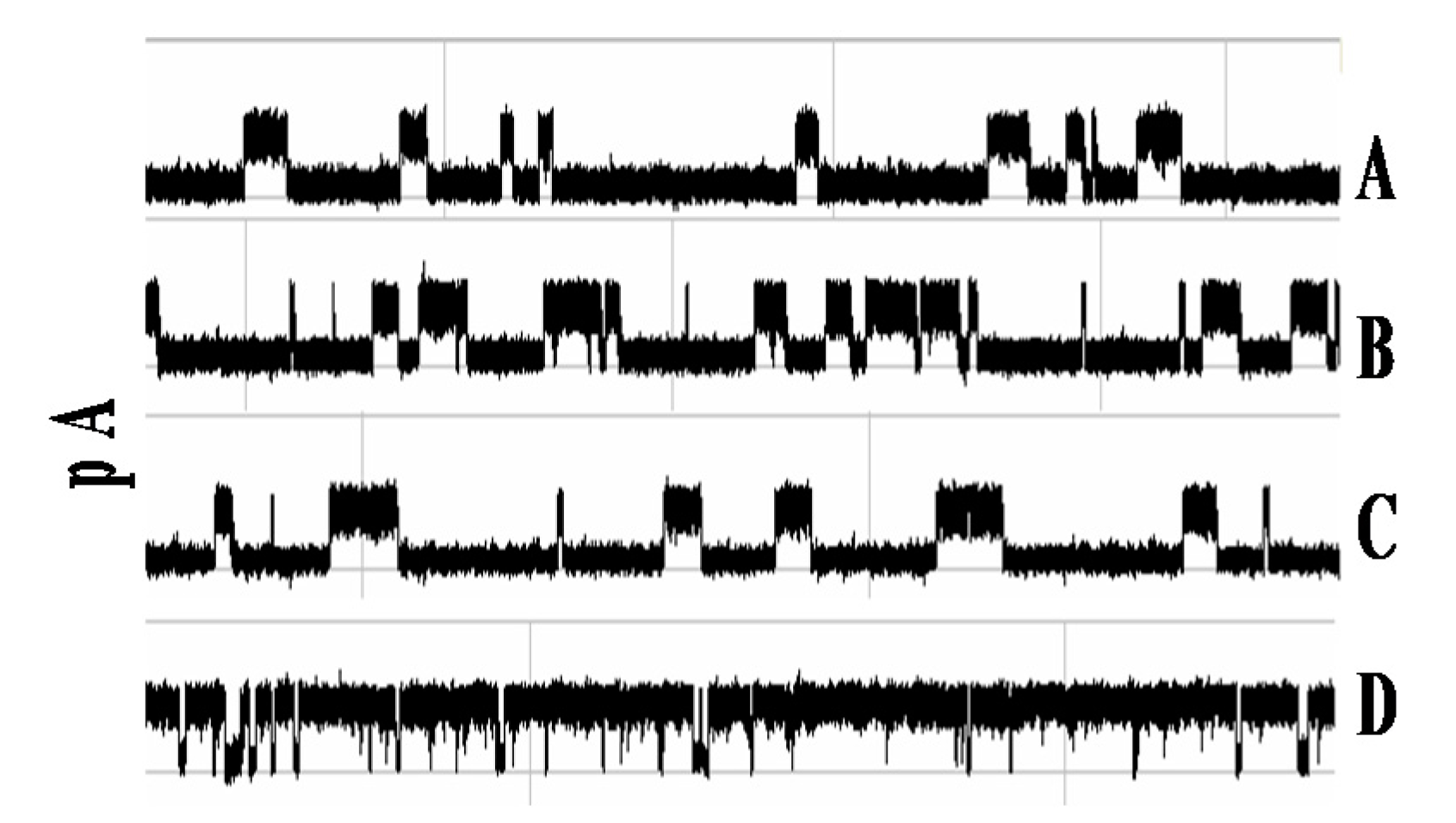
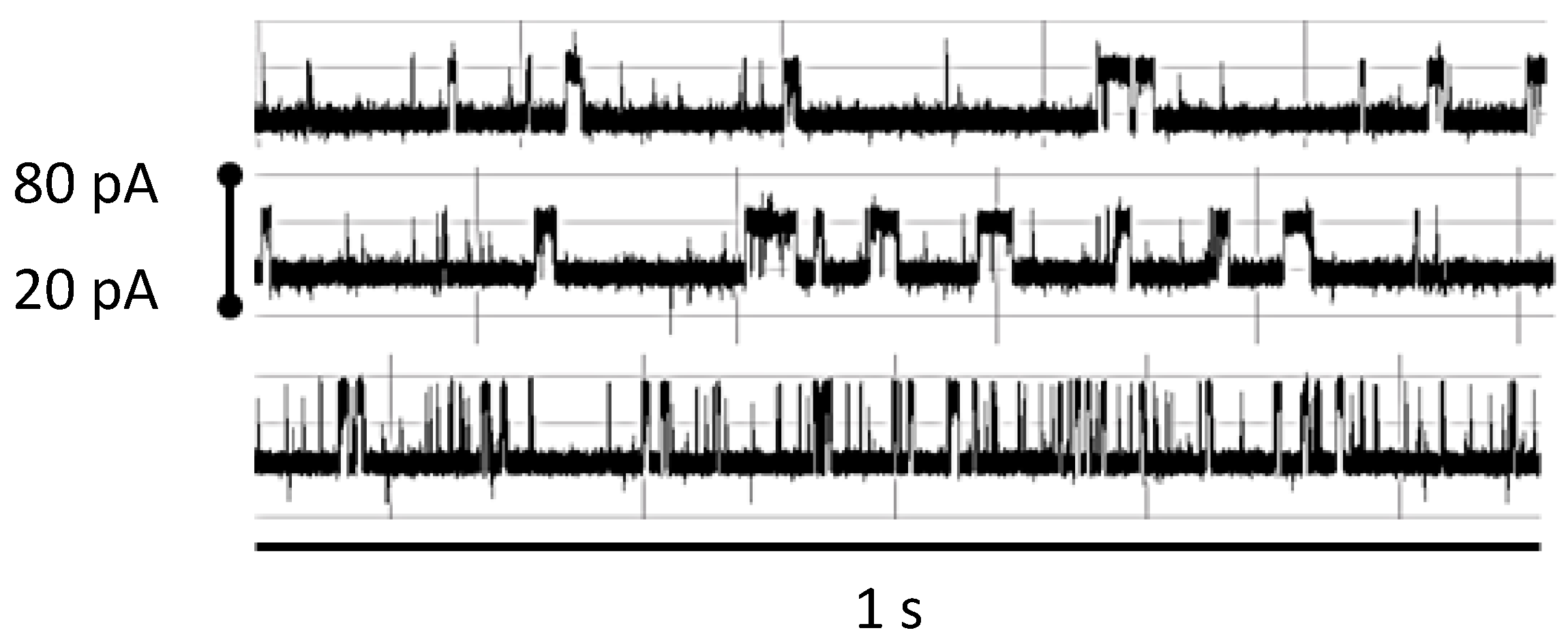
3.1.2. Observations of Biotin-Streptavidin Binding on the NTD Nanoscope
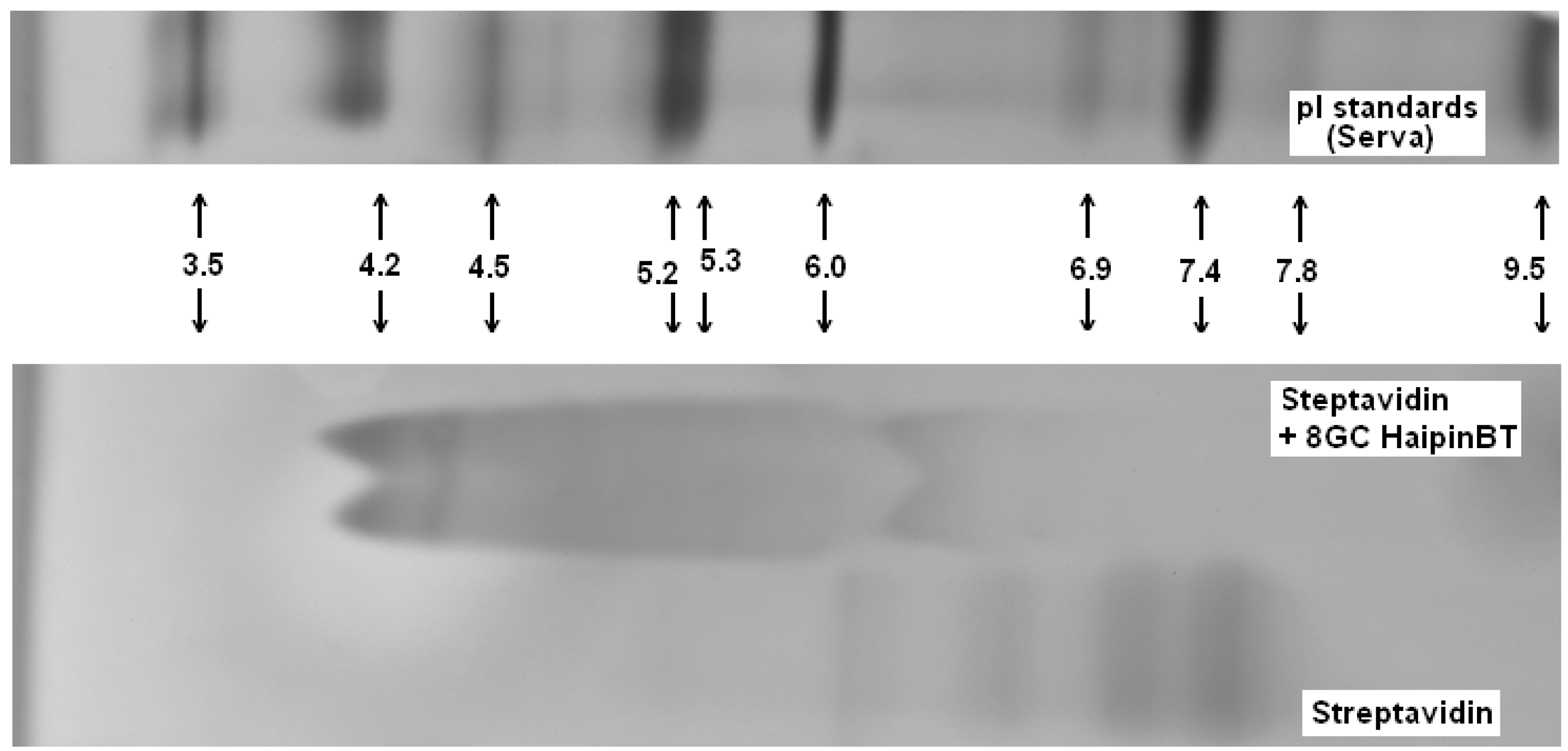
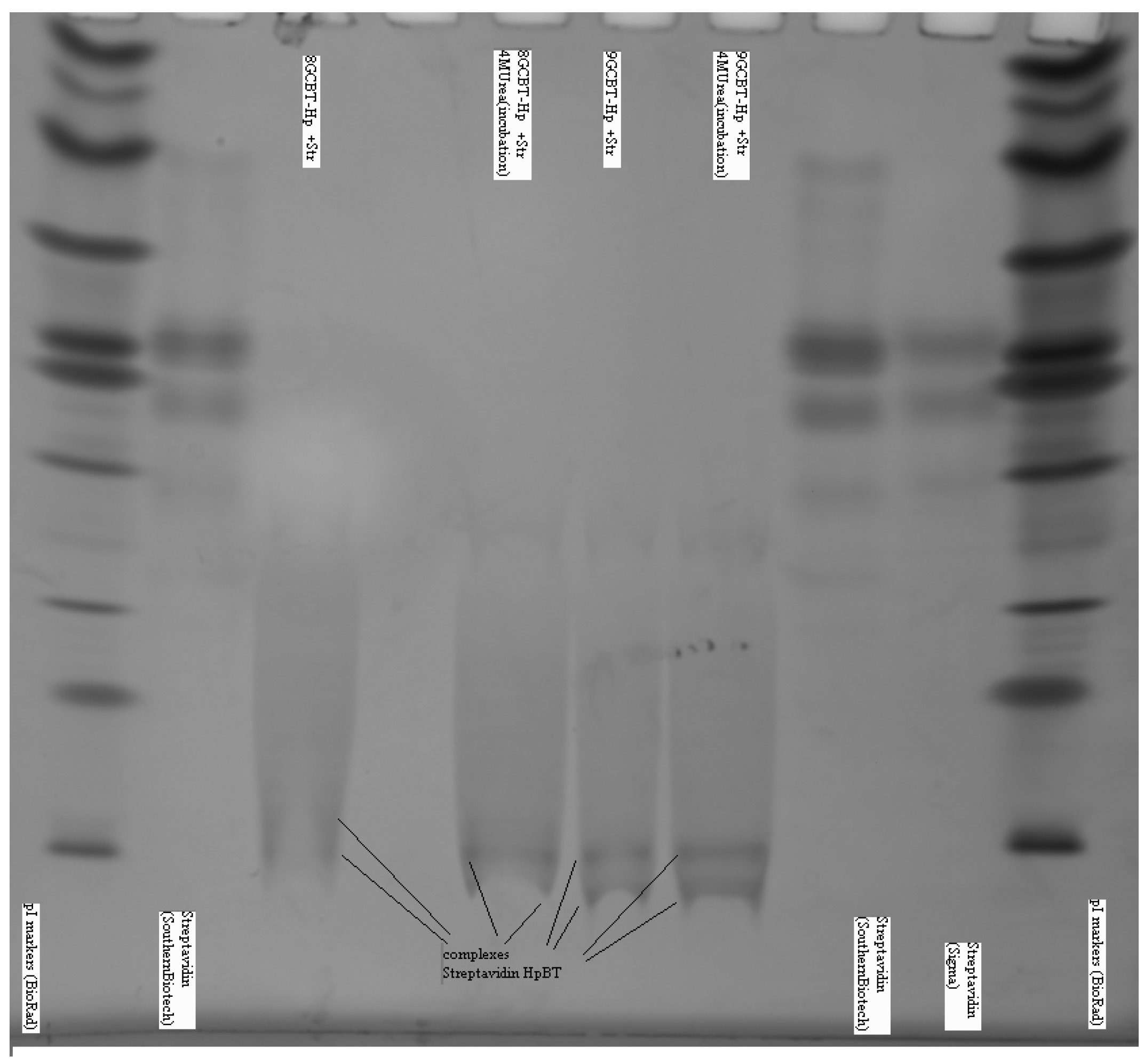
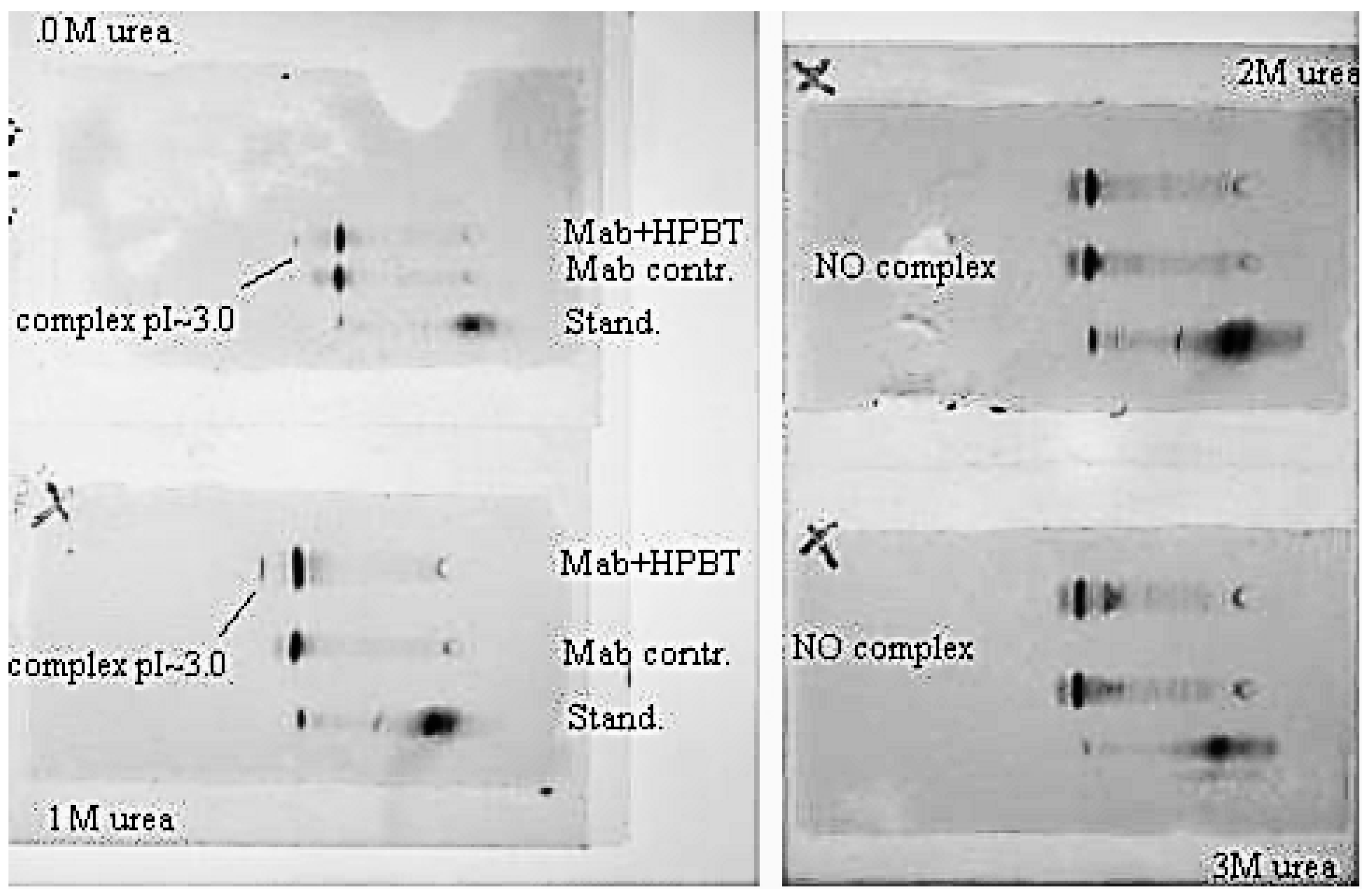
3.1.3. Bt-8gc—Streptavidin Binding Validation Using Gel Isoelectric Focusing (IEF) (3–10 pH Range) in Presence of Chaotropes
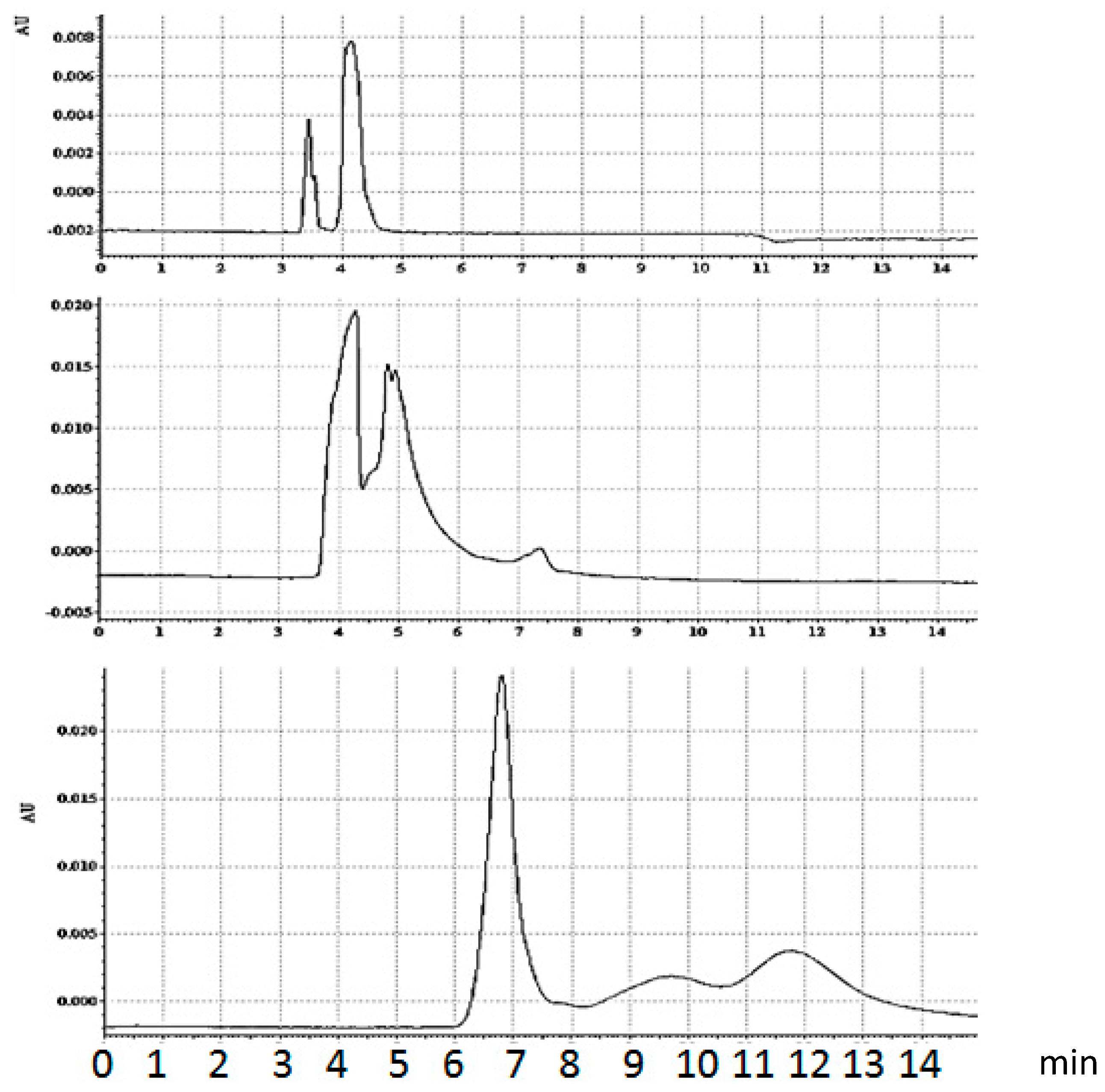
3.1.4. Bt-8gc—Streptavidin Binding Validation Using CE in the Presence of Chaotropes
3.2. Biotin-mAb Binding Experiments
3.2.1. Observations of Biotin-mAb Binding on the NTD Nanoscope
3.2.2. Bt-8gc—mAb Binding Validation Using gel IEF (3–10) in presence of chaotropes
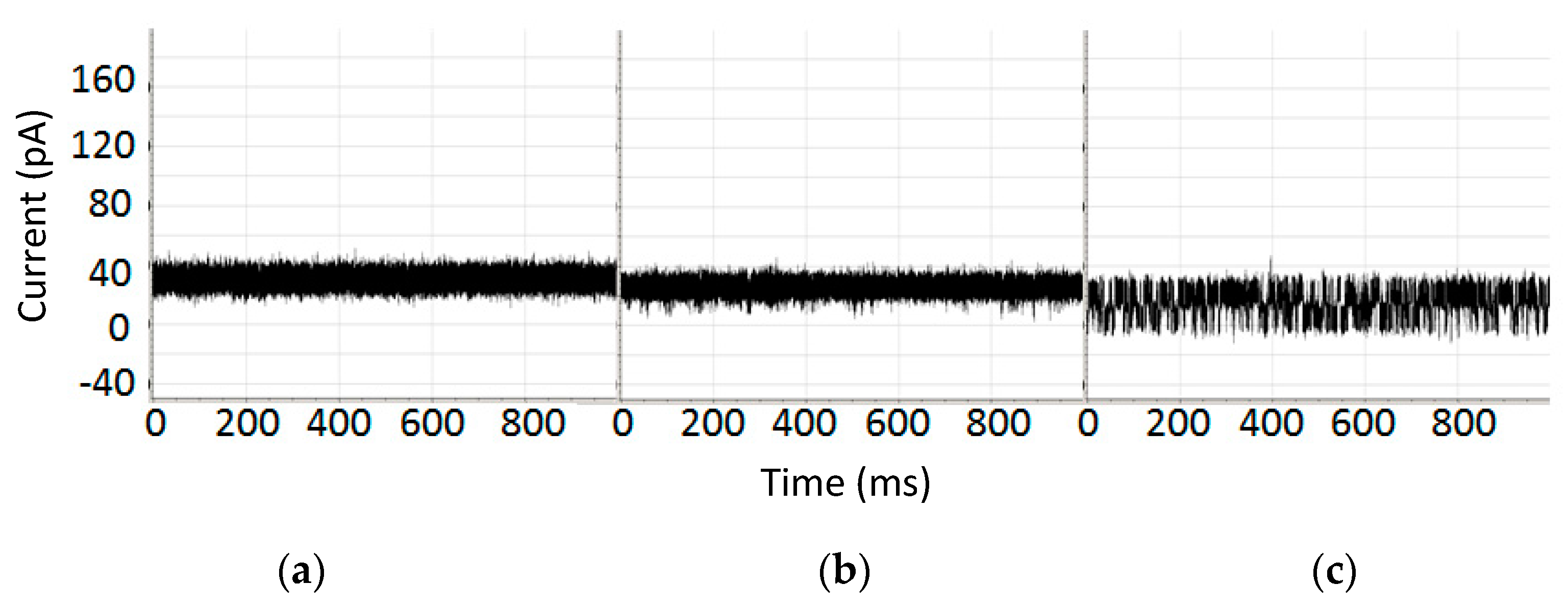
3.3. Alpha-Hemolysin Nanoscope Operational pH Range
4. Experimental Section
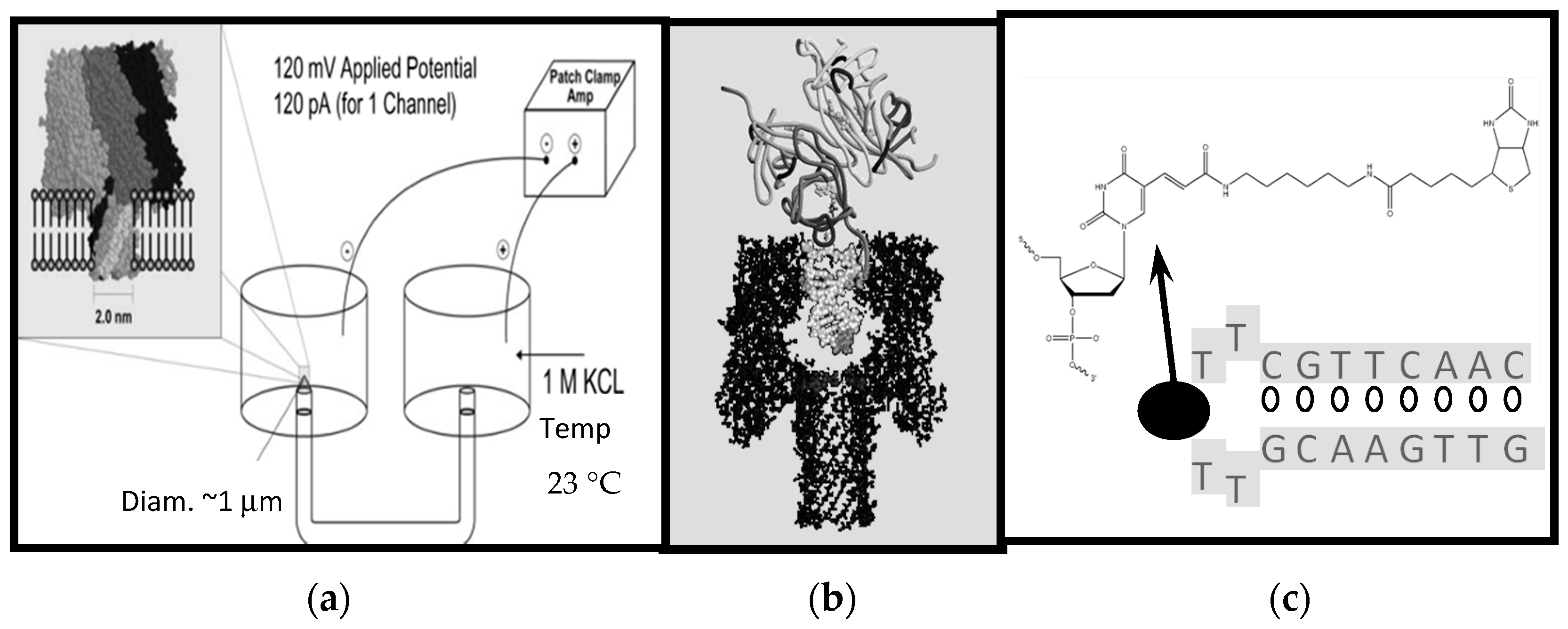
4.1. Nanopore Experiments
4.2. Control Probe Design
4.3. NTD-Aptamer Design
4.4. Gel Electrophoresis and Image Analysis
4.5. Capillary Electrophoresis (CE)
4.6. Chemicals
4.7. Methods for Channel Current Cheminformatics
5. Discussion
5.1. Validation of NTD Complexation Detection Using Standard Electrophoretic Techniques
5.2. Nanopore Detectors as In-Silico Gels
5.3. Nanopore Detector Tolerance of Chaotropes and High Salt
5.4. Purity Tests
5.5. NTD Capabilities and Limitations
5.6. Processive DNA Enzyme with Laser/Dye info for Enhanced Fidelity for Very Long Reads
6. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Coulter, W.H. 1953. Means for Counting Particles Suspended in a Fluid. U.S. Patent 2656508, 20 October 1953. [Google Scholar]
- DeBlois, R.W.; Bean, C.P. Counting and sizing of submicron particles by the resistive pulse technique. Rev. Sci. Instrum. 1970, 41, 909–916. [Google Scholar] [CrossRef]
- Bean, C.P. The physics of porous membranes—Neutral pores. In Membranes; Eisenman, G., Ed.; Marcel Dekker: New York, NY, USA, 1972; pp. 1–54. [Google Scholar]
- DeBlois, R.W.; Bean, C.P.; Wesley, R.K.A. Electrokinetic measurements with submicron particles and pores by the resistive pulse technique. J. Colloid Interface Sci. 1977, 61, 323–335. [Google Scholar] [CrossRef]
- Hladky, S.B.; Haydon, D.A. Ion transfer across lipid membranes in the presence of gramicidin A. Biochim. Biophys. Acta 1972, 274, 294–312. [Google Scholar] [CrossRef]
- Bezrukov, S.M.; Vodyanoy, I.; Parsegian, V.A. Counting polymers moving through a single ion channel. Nature 1994, 370, 279–281. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; McMullan, C.; Stein, D.; Branton, D.; Golovchenko, J. Solid state nanopores for single molecule detection. Biophys. J. 2001, 80, 339A. [Google Scholar]
- Li, J.; Stein, D.; McMullan, C.; Branton, D.; Aziz, M.J.; Golovchenko, J.A. Ion beam sculpting at nanometer length scales. Nature 2001, 412, 166–169. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Hobaugh, M.R.; Shustak, C.; Cheley, S.; Bayley, H.; Gouaux, J.E. Structure of staphylococcal α-hemolysin, a heptameric transmembrane pore. Science 1996, 274, 1859–1866. [Google Scholar] [CrossRef] [PubMed]
- Kullman, L.; Winterhalter, M.; Bezrukov, S.M. Transport of maltodextrins through maltoporin: A single-channel study. Biophys. J. 2002, 82, 803–812. [Google Scholar] [CrossRef]
- Palonsky, S.; Rossnagel, S.; Stolovitsky, G. Nanopore in metal-dielectric sandwich for DNA positional control. Appl. Phys. Lett. 2007, 91. [Google Scholar] [CrossRef]
- Luan, B.; Pemg, H.; Polonsky, S.; Rossnagel, S.; Stolovitsky, G.; Martyna, G. Base-by-Base ratcheting of single stranded DNA through a solid-state nanopore. Phys. Rev. Lett. 2010, 104. [Google Scholar] [CrossRef] [PubMed]
- Jain, M.; Fiddes, I.T.; Miga, K.H.; Olsen, H.; Paten, B.; Akeson, M. Improved data analysis for the MinION nanoipore sequencer. Nat. Methods 2015, 12, 351–356. [Google Scholar] [CrossRef] [PubMed]
- Schreiber, J.; Wescoe, Z.L.; Abu-Shumays, R.; Vivian, J.T.; Baatar, B.; Karplus, K.; Akeson, M. Error rates for nanopore discrimination among cytosine, methylcytosine, and hydroxymethylcytosine along individual DNA strands. Proc. Natl. Acad. Sci. USA 2013, 110, 18910–18915. [Google Scholar] [CrossRef] [PubMed]
- Ying, Y.-L.; Zhang, J.; Gao, R.; Long, Y.-T. Nanopore-based sequencing and detection of nucleic acids. Angew. Chem. Int. Ed. 2013, 42, 13154–13161. [Google Scholar]
- Winters-Hilt, S.; Pincus, S. Nanopore-Based Biosensing. US 2009/0054919 A2, 16 February 2012. [Google Scholar]
- Winters-Hilt, S.; and Pincus, S. Channel Current Cheminformatics and Bioengineering Methods for Immunological Screening, Single-Molecule Analysis, and Single Molecular-Interaction Analysis. WO2006041983, 8 March 2007. [Google Scholar]
- Winters-Hilt, S. Channel Current Cheminformatics and Bioengineering Methods for Immunological Screening, Single-Molecule Analysis, and Single Molecular-Interaction Analysis. EP 1805680, 24 March 2015. [Google Scholar]
- Winters-Hilt, S. Nanopore Detector based analysis of single-molecule conformational kinetics and binding interactions. BMC Bioinform. 2006, 7, S2–S21. [Google Scholar] [CrossRef] [PubMed]
- Winters-Hilt, S.; Horton-Chao, E.; Morales, E. The NTD Nanoscope: potential applications and implementations. BMC Bioinform. 2011, 12, S10–S21. [Google Scholar] [CrossRef] [PubMed]
- Winters-Hilt, S.; Adelman, R. Method and System for Characterizing or Identifying Molecules and Molecular Mixtures. U.S. Patent 20130071837 A1, 21 March 2013. [Google Scholar]
- Winters-Hilt, S. Machine-Learning Based Sequence Analysis, Bioinformatics & Nanopore Transduction Detectio; Lulu Press: Raleigh, NC, USA, 2011; ISBN 978-1-257-64525-1. [Google Scholar]
- Eren, A.M.; Amin, I.; Alba, A.; Morales, E.; Stoyanov, A.; Winters-Hilt, S. Pattern recognition informed feedback for nanopore detector cheminformatics. Adv. Exp. Med. Biol. 2010, 680, 99–108. [Google Scholar] [PubMed]
- Winters-Hilt, S. Method and System for Stochastic Carrier Wave Communications, Radio-Noise Embedded Steganography, and Robust Self-Tuning Signal Discovery and Data-Mining. US Patent Pending SN 62/186827.
- Vercoutere, W.; Winters-Hilt, S.; Olsen, H.; Deamer, D.; Haussler, D.; Akeson, M. Rapid discrimination among individual DNA molecules at single nucleotide resolution using an ion channel. Nat. Biotechnol. 2001, 19, 248–252. [Google Scholar] [CrossRef] [PubMed]
- Winters-Hilt, S.; Vercoutere, W.; DeGuzman, V.S.; Deamer, D.; Akeson, M.; Haussler, D. Highly accurate classification of watson-crick base-pairs on termini of single DNA molecules. Biophys. J. 2003, 84, 967–976. [Google Scholar] [CrossRef]
- Winters-Hilt, S.; Davis, A.; Amin, I.; Morales, E. Nanopore current transduction analysis of protein binding to non-terminal and terminal DNA regions: Analysis of transcription factor binding, retroviral DNA terminus dynamics, and retroviral integrase-DNA binding. BMC Bioinform. 2007, 8, S7–S10. [Google Scholar] [CrossRef] [PubMed]
- Winters-Hilt, S.; Landry, M.; Akeson, M.; Tanase, M.; Amin, I.; Coombs, A.; Morales, E.; Millet, J.; Baribault, C.; Sendamangalam, S. Cheminformatics methods for novel nanopore analysis of HIV DNA termini. BMC Bioinform. 2006, 7, S2–S22. [Google Scholar] [CrossRef] [PubMed]
- Righetti, P.G.; Stoyanov, A.V.; Zhukov, M. The Proteome Revisited; Elsevier: Amsterdam, The Netherlands, 2001. [Google Scholar]
- Cann, J.R.; Stimpson, D.I.; Cox, D.J. Isoelectric focusing of interacting systems: Carrier ampholyte-induced macromolecular association or dissociation into subunits. Anal. Biochem. 1978, 86, 34–49. [Google Scholar] [CrossRef]
- Kabytaev, K.; Durairaj, A.; Shin, D.; Rohlfing, C.L.; Connolly, S.; Little, R.R.; Stoyanov, A.V. Two-step ion-exchange chromatographic purification combined with reversed-phase chromatography to isolate C-peptide for mass spectrometric analysis. J. Sep. Sci. 2016, 39, 676–681. [Google Scholar] [CrossRef] [PubMed]
- Stoyanov, A.V.; Rohlfing, C.L.; Connolly, S.; Roberts, M.L.; Nauser, C.L.; Little, R.R. Use of cation exchange chromatography for human C-peptide isotope dilution–mass spectrometric assay. J. Chromatogr. A 2011, 1218, 9244–9249. [Google Scholar] [CrossRef] [PubMed]
- Stoyanov, A.V.; Righetti, P.G. Ampholyte dissociation theory and properties of ampholyte aqueous solutions. Electrophoresis 1987, 18, 1944–1950. [Google Scholar] [CrossRef] [PubMed]
- Andreev, V.P.; Makarova, E.; Pliss, N.S. New capability of electroinjection analysis: Investigation of chemical reaction kinetics. Anal. Chem. 2001, 73, 1316–1323. [Google Scholar] [CrossRef]
- Patterson, D.H.; Harmon, B.J.; Regnier, F.E. Dynamic modeling of electrophoretically mediated microanalysis. J. Chromatogr. A 1996, 732, 119–132. [Google Scholar] [CrossRef]
- Shalongo, W.; Jagannadham, M.; Stellwagen, E. Kinetic analysis of the hydrodynamic transition accompanying protein folding using size exclusion chromatography. Comparison of spectral and chromatographic kinetic analyses. Biopolymers 1993, 33, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Stoyanov, A.V.; Righetti, P.G. Steady-state electrolysis of a solution of nonamphotheric compounds. Electrophoresis 1999, 20, 718–722. [Google Scholar] [CrossRef]
- Stoyanov, A.V. Water electrolysis in mono-and hetero-phase low conductive systems and a secondary pH gradient establishment. In Electrolysis: Theory, Types and Applications; Kuai, S., Meng, J., Eds.; Nova Science Publishers Inc.: Hauppauge, NY, USA, 2010; pp. 511–516. [Google Scholar]
- Stoyanov, A.V.; Pawliszyn, J. Buffer composition changes in background electrolyte during electrophoretic run in capillary zone electrophoresis. Analyst 2004, 129, 979–982. [Google Scholar] [CrossRef] [PubMed]
- Stoyanov, A. IEF-based multidimensional applications in proteomics: Toward higher resolution. Electrophoresis 2012, 33, 3281–3290. [Google Scholar] [CrossRef] [PubMed]
- Stoyanov, A.V.; Rogatsky, E.; Stein, D.; Connolly, S.; Rohlfing, C.L.; Little, R.R. Isotope dilution assay in peptide quantification: The challenge of microheterogeneity of internal standard. Proteom. Clin. Appl. 2013, 7, 825–828. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the compounds are not available from the authors.












© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Winters-Hilt, S.; Stoyanov, A. Nanopore Event-Transduction Signal Stabilization for Wide pH Range under Extreme Chaotrope Conditions. Molecules 2016, 21, 346. https://doi.org/10.3390/molecules21030346
Winters-Hilt S, Stoyanov A. Nanopore Event-Transduction Signal Stabilization for Wide pH Range under Extreme Chaotrope Conditions. Molecules. 2016; 21(3):346. https://doi.org/10.3390/molecules21030346
Chicago/Turabian StyleWinters-Hilt, Stephen, and Alexander Stoyanov. 2016. "Nanopore Event-Transduction Signal Stabilization for Wide pH Range under Extreme Chaotrope Conditions" Molecules 21, no. 3: 346. https://doi.org/10.3390/molecules21030346
APA StyleWinters-Hilt, S., & Stoyanov, A. (2016). Nanopore Event-Transduction Signal Stabilization for Wide pH Range under Extreme Chaotrope Conditions. Molecules, 21(3), 346. https://doi.org/10.3390/molecules21030346