
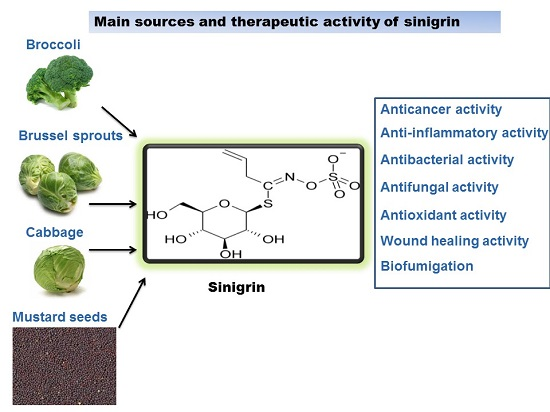
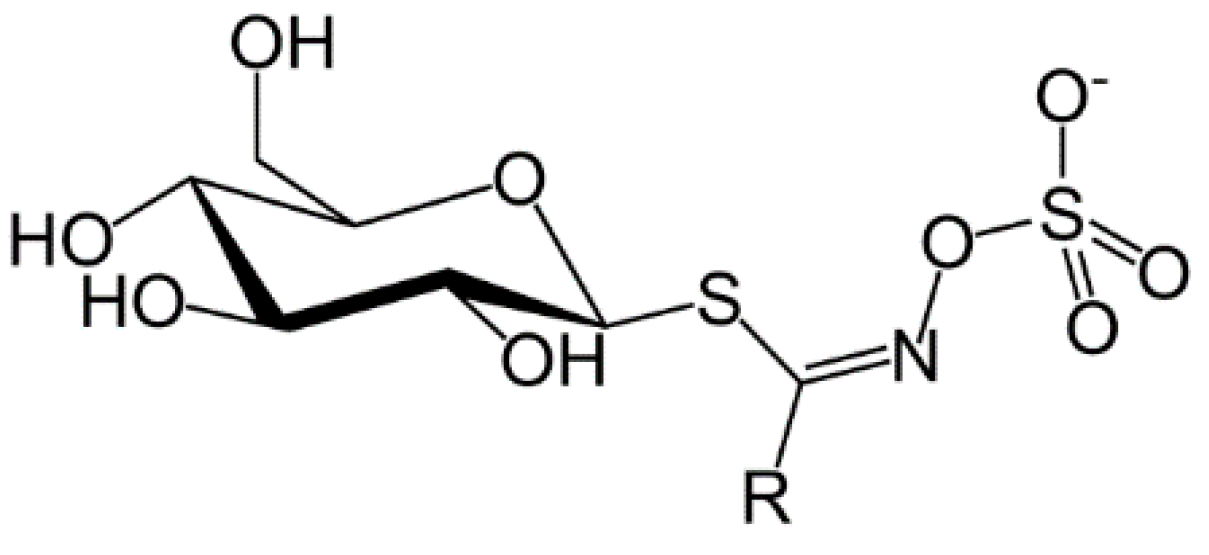
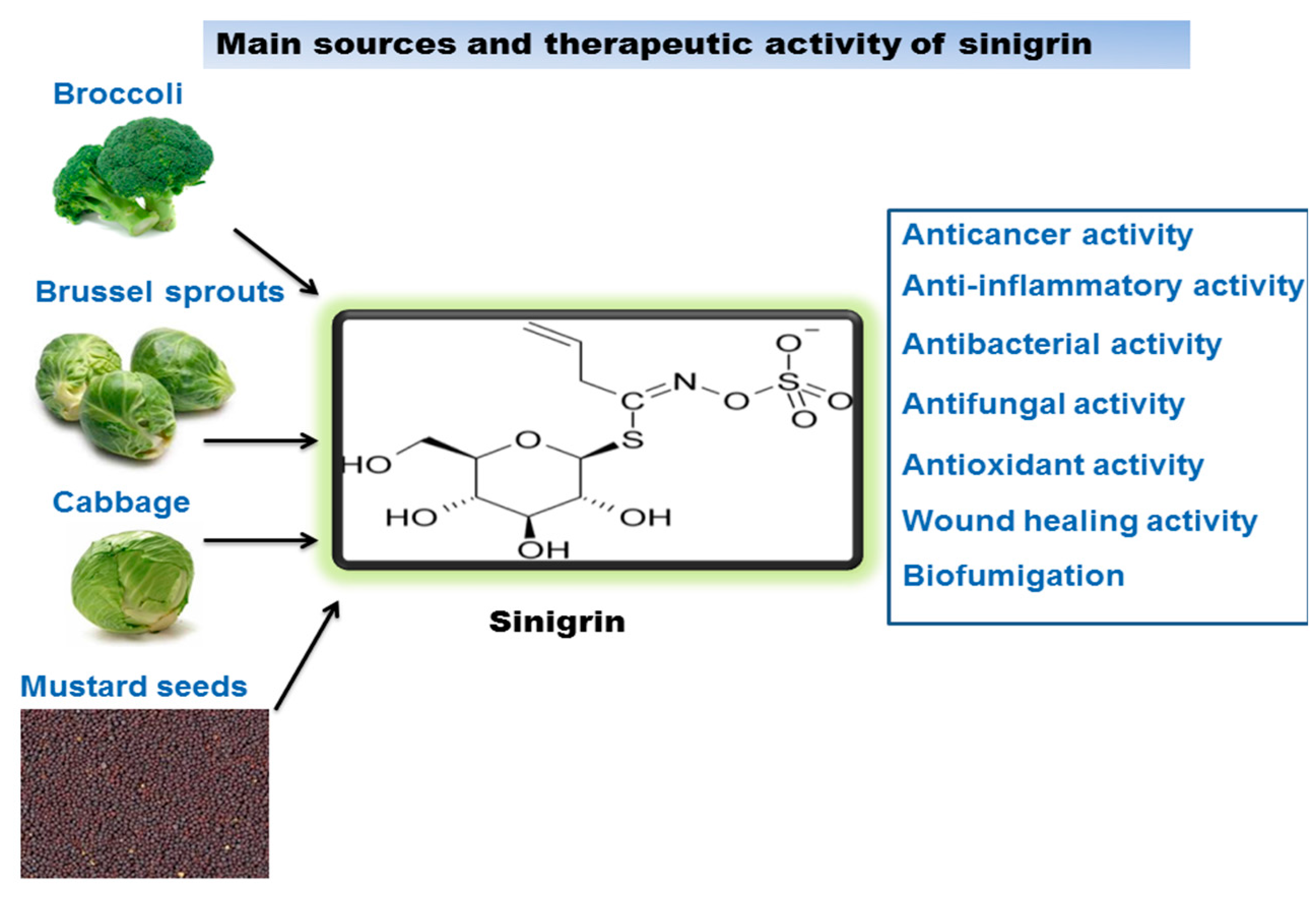
Sinigrin and Its Therapeutic Benefits


Abstract
:
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Extraction of Sinigrin
3. Therapeutic Benefits of Sinigrin
3.1. Anticancer Activity
3.2. Anti-Inflammatory Activity
3.3. Antibacterial Activity
3.4. Antifungal Activity
3.5. Antioxidant Activity
3.6. Wound Healing Activity
3.7. Biofumigation
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ekor, M. The growing use of herbal medicines: Issues relating to adverse reactions and challenges in monitoring safety. Front. Pharmacol. 2014, 4, 177. [Google Scholar] [CrossRef] [PubMed]
- Johnson, I.T. New approaches to the role of diet in the prevention of cancers of the alimentary tract. Mutat. Res. 2004, 551, 9–28. [Google Scholar] [CrossRef] [PubMed]
- Johnson, I.T. Phytochemicals and cancer. Proc. Nutr. Soc. 2007, 66, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, D.W.; Birch, A.N.E.; Hillman, J.R. Antinutritional coumponds in the Brassicaceae: Analysis, biosynthesis, chemistry and dietary effects. J. Hortic. Sci. Biotechnol. 1998, 73, 1–18. [Google Scholar] [CrossRef]
- Chew, F.S. Biological effects of glucosinolates. In Biologically Active Natural Products: Potential Use in Agriculture; Cutler, H.G., Ed.; American Chemical Society: Washington, DC, USA, 1988; pp. 155–181. [Google Scholar]
- Jahgangir, M.J.; Kim, H.K.; Choi, Y.H.; Verpoorte, R. Health-affecting compounds in Brassicaceae. Compr. Rev. Food Sci. Food Saf. 2009, 8, 31–34. [Google Scholar] [CrossRef]
- Challenger, F. The natural mustard oil glucosides and the related isothiocyanates and nitriles. In Aspects of the Organic Chemistry of Sulphur; Academic Press: New York, NY, USA, 1959; pp. 115–161. [Google Scholar]
- Tianxin, W.; Liang, H.; Qipeng, Y. Separation of sinigrin from Indian mustard (Brassica juncea L.) seed using macroporous ion-exchange resin. Korean J. Chem. Eng. 2012, 29, 396–403. [Google Scholar]
- Redovnikovic, I.R.; Glivetic, T.; Delonga, K.; Vorkapić-Furač, J. Glucosinolates and their potential role in plant. Period. Biol. 2008, 110, 297–309. [Google Scholar]
- Manohar, R.P.; Pushpan, R.; Rohini, S. Mustard and its uses in Ayurveda. Indian J. Tradit. Know. 2009, 8, 400–404. [Google Scholar]
- Nomura, T.; Shinoda, S.; Yamori, T.; Sawaki, S.; Nagata, I.; Ryoyama, K.; Fuke, Y. Selective sensitivity to wasabi-derived 6-(methylsulfinyl)hexyl isothiocyanate of human breast cancer and melanoma cell lines studied in vitro. Cancer Detect. Prev. 2005, 29, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Washida, K.; Miyata, M.; Koyama, T.; Yazawa, K.; Nomoto, K. Suppressive effect of Yamato-mana (Brassica rapa L. Oleifera Group) constituent 3-butenyl glucosinolate (gluconapin) on postprandial hypertriglyceridemia in mice. Biosci. Biotechnol. Biochem. 2010, 74, 1286–1289. [Google Scholar] [CrossRef] [PubMed]
- Holst, B.; Williamson, G. A critical review of the bioavailability of glucosinolates and related compounds. Nat. Prod. Rep. 2004, 21, 425–447. [Google Scholar] [CrossRef] [PubMed]
- Rangkadilok, N.; Nicolasa, M.E.; Bennettc, R.N.; Premierb, R.R.; Eaglingb, D.R.; Taylora, P.W.J. Determination of sinigrin and glucoraphanin in Brassica species using a simple extraction method combined with ion-pair HPLC analysis. Sci. Hortic. 2002, 96, 27–41. [Google Scholar] [CrossRef]
- Kushad, M.M.; Brown, A.F.; Kurilich, A.C.; Juvik, J.A.; Klein, B.P.; Wallig, M.A.; Jeffery, E.H. Variation of glucosinolates in vegetable crops of Brassica oleracea. J. Agric. Food Chem. 1999, 47, 1541–1548. [Google Scholar] [CrossRef] [PubMed]
- Szmigielska, A.M.; Schoenau, J.J.; Levers, V. Exchange membrane extraction combined with the high-pressure liquid chromatography detection. J. Agric. Food Chem. 2000, 48, 4487–4491. [Google Scholar] [CrossRef] [PubMed]
- Mohn, T.; Cutting, B.; Ernst, B.; Hamburger, M. Extraction and analysis of intact glucosinolates—A validated pressurized liquid extraction/liquid chromatography-mass spectrometry protocol for Isatis tinctoria, and qualitative analysis of other cruciferous plants. J. Chromatogr. A 2007, 1166, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Liang, H.; Yuan, Q. Optimization of ultrasonic-stimulated solvent extraction of sinigrin from Indian mustard seed (Brassica Juncea L.) using response surface methodology. Phytochem. Anal. 2011, 22, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Cools, K.; Terry, L.A. Comparative study between extraction techniques and column separation for the quantification of sinigrin and total isothiocyanates in mustard seed. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2012, 15, 115–118. [Google Scholar] [CrossRef] [PubMed]
- Belliveau, K.A.; Romero-Zerón, L.B. Monitoring the enzymatic degradation of sinigrin from B. juncea meal using 1H-NMR spectroscopy. Nat. Prod. Res. 2010, 24, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.H.; Huang, J.W.; Kumar, P.V.; Jen, J.F. Determination of sinigrin in vegetables seeds by online microdialysis sampling coupled reverse- phase ion-pair liquid chromatography. J. Agric. Food Chem. 2010, 58, 4571–4575. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.L.; Chen, C.; Zhou, M.; Wang, J.Z.; Luo, X.; Huang, G.; Ye, L.M. Determination of sinigrin in Semen Thalaspi from Sichun and Tibet using near infrared diffuse reflectance spectroscopy. Guang Pu Xue Yu Guang Pu Fen Xi 2009, 29, 2673–2676. [Google Scholar] [PubMed]
- Rochfort, S.J.; Trenerry, V.C.; Imsic, M.; Panozzo, J.; Jones, R. Class targeted metabolomics: ESI ion trap screening methods for glucosinolates based on MSN fragementation. Phytochemistry 2008, 69, 1671–1679. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.C.; Cheuk, M.W.; Chan, W.; Lee, A.W.; Zhao, Z.Z.; Jiang, Z.H.; Cai, Z. Determination of glucosinolates in traditional Chinese herbs by high-performance liquid chromatography and electrospray ionization mass spectrometry. Anal. Bioanal. Chem. 2006, 386, 2225–2232. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, A.; Li, Y.; Wade, K.L.; Paonessa, J.D.; Fahey, J.W.; Zhang, Y. Allyl isothiocyanate-rich mustard seed powder inhibits bladder cancer growth and muscle invasion. Carcinogenesis 2010, 31, 2105–2110. [Google Scholar] [CrossRef] [PubMed]
- Jie, M.; Cheung, W.M.; Yu, V.; Zhou, Y.; Tong, P.H.; Ho, J.W.S. Anti-proliferative activities of sinigrin on carcinogen-induced hepatotoxicity in rats. PLoS ONE 2014, 9, e110145. [Google Scholar] [CrossRef] [PubMed]
- Lozano-Baena, M.D.; Tasset, I.; Obregon-Cano, S.; de Haro-Bailon, A.; Muñoz-Serrano, A.; Alonso-Moraga, A. Antigenotoxicity and tumor growing inhibition by leafy Brassica carinata and Sinigrin. Molecules 2015, 20, 15748–15765. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.K.; Lund, E.K.; Johnson, I.T. Inhibition of dimethylhydrazine-induced aberrant crypt foci and induction of apoptosis in rat colon following oral administration of the glucosinolate sinigrin. Carcinogenesis 1998, 19, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Morse, M.A.; Wang, C.X.; Amin, S.G.; Hecht, S.S.; Chung, F.L. Effects of dietary sinigrin or indole-3-carbinol on O6-methylguanine-DNA-transmethylase activity and 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone-induced DNA methylation and tumorigenicity in F344 rats. Carcinogenesis 1988, 9, 1891–1895. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Mori, Y.; Morishita, Y.; Hara, A.; Ohno, T.; Kojima, T.; Mori, H. Inhibitory effect of sinigrin and indole-3-carbinol on diethylnitrosamine-induced hepatocarcinogenesis in male ACI/N rats. Carcinogenesis 1990, 11, 1403–1406. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Kojima, T.; Morishta, Y.; Moori, H. Inhibitory effects of the natural products indole-3-carbinol and Sinigrin during initiation and promotion phases of 4-nitroquinoline 1-oxide-induced rat tongue carcinogenesis. Jpn. J. Cancer Res. 1992, 83, 835–842. [Google Scholar] [CrossRef] [PubMed]
- Wattenberg, L.W. Inhibition of carcinogenesis by minor nutrient constituents of the diet. Proc. Nutr. Soc. 1990, 49, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Barcelo, S.; Gardiner, J.M.; Gescher, A.; Chipman, J.K. CYP2E1-mediated mechanism of anti-genotoxicity of the broccoli constituent sulforaphane. Carcinogenesis 1996, 17, 277–282. [Google Scholar] [CrossRef] [PubMed]
- Thompson, C.B. Apoptosis in the pathogenesis and treatment of disease. Science 1995, 267, 1456–1462. [Google Scholar] [CrossRef] [PubMed]
- Mazumder, A.; Dwivedi, A.; Du Preez, J.; Du Plessis, J. In vitro wound healing and cytotoxic effects of sinigrin–phytosome complex. Int. J. Pharm. 2015, 498, 283–293. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Lee, C.; Kim, J.; Pyo, S. The inhibitory effect of sinigrin on the production of inflammatory mediators induced by lipopolysaccharide in RAW 264.7 macrophages (1056.5). FASEB J. 2014, 28, 1056. [Google Scholar]
- Jang, Y.J.; Pyo, S. Anti-atherosclerotic effect of sinigrin in ApoE-deficient mice. FASEB J. 2015, 29, 609.1. [Google Scholar]
- Lee, H.W.; Lee, K.R. Effect of sinigrin on vascular cell adhesion molecule-1 expression in TNF-α-stimulated mouse vascular smooth muscle cells via downregulation of NF-κB signalling pathways. FASEB J. 2015, 29, 593.15. [Google Scholar]
- Luciano, F.B.; Holley, R.A. Enzymatic inhibition by allyl isothiocyanate and factors affecting its antimicrobial action against Escherichia coli O157:H7. Int. J. Food Microbiol. 2009, 131, 240–245. [Google Scholar] [CrossRef] [PubMed]
- Gamage, G.R.; Park, H.J.; Kim, K.M. Effectiveness of antimicrobial coated oriented polypropylene/polyethylene films in sprout packaging. Food Res. Int. 2009, 42, 832–839. [Google Scholar] [CrossRef]
- Zsolnai, T. The antimicrobial activity of thiocyanates and isothiocyantes. Arzneimittel-Forschung 1966, 16, 870–876. [Google Scholar] [PubMed]
- Li, T.; Zhang, Y. Mitochondria are the primary target in isothiocyanate-induced apoptosis in human bladder cancer cells. Mol. Cancer Ther. 2005, 4, 1250–1259. [Google Scholar]
- Lin, C.M.; Preston, J.F.; Wei, C.I. Antibacterial mechanism of allyl isothiocyanate. J. Food Prot. 2000, 63, 727–734. [Google Scholar] [PubMed]
- Cordeiro, R.P.; Wu, C.; Holley, R.A. Contribution of endogenous plant myrosinase to the antimicrobial activity of deodorized mustard against Escherichia coli O157: H7 in fermented dry sausage. Int. J. Food Microbiol. 2014, 189, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Herzallah, S.; Holley, R. Use of a nanoparticulate carboxymethyl cellulose film containing sinigrin as an antimicrobial precursor to kill Escherichia coli O157:H7 on fresh beef. Lett. Appl. Microbiol. 2015, 61, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Shofran, B.G.; Purrington, S.; Breidt, F.; Fleming, H. Antimicrobial properties of sinigrin and its hydrolysis products. J. Food Sci. 1998, 63, 621–624. [Google Scholar] [CrossRef]
- Brabban, A.D.; Edwards, C. The effects of glucosinolates and their hydrolysis products on microbial growth. J. Appl. Bacteriol. 1995, 79, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Lara-Lledo, M.; Olaimat, A.; Holley, R.A. Inhibition of Listeria monocytogenes on bologna sausages by an antimicrobial film containing mustard extract or sinigrin. Int. J. Food Microbiol. 2012, 156, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Schreiner, R.P.; Koide, R.T. Antifungal compounds from the roots of mycotrophic and non-mycotrophic plant species. New Phytol. 1993, 123, 99–105. [Google Scholar] [CrossRef]
- Ocampo, J.A.; Martin, J.; Hayman, D.S. Influence of plant interactions on vesicular-arbuscular mycorrhizal infections. Host and non-host plants grown together. New Phytol. 1980, 84, 27–35. [Google Scholar] [CrossRef]
- Maria, M.; Leoni, O.; Lori, R.; Cembali, T. Antifungal vapour–phase activity of allyl–isothiocyanate against Penicillium expansum on pears. Plant Pathol. 2002, 51, 231–236. [Google Scholar] [CrossRef]
- Ippoushi, K.; Takeuchi, A.; Azuma, K. Sinigrin suppresses nitric oxide production in rats administered intraperitoneally with lipopolysaccharide. Food Chem. 2010, 120, 1119–1121. [Google Scholar] [CrossRef]
- Peluffo, G.; Radi, R. Biochemistry of protein tyrosine nitration in cardiovascular pathology. Cardiovasc. Res. 2007, 75, 291–302. [Google Scholar] [CrossRef] [PubMed]
- Hanschen, F.S.; Yim, B.; Winklemann, T.; Smalla, K.; Schreiner, M. Degradation of biofumigant isothiocyanates and allyl glucosinolate in soil and their effects on the microbial community composition. PLoS ONE 2015, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fenwick, G.R.; Heaney, R.K.; Mullin, W.J. Glucosinolates and their breakdown products in food and food plants. CRC Crit. Rev. Food Sci. Nutr. 1983, 18, 123–201. [Google Scholar] [CrossRef]
- Wittstock, U.; Burow, M. Glucosinolate breakdown in Arabidopsis: Mechanism, regulation and biological significance. Arabidopsis Book 2010, 8, e0134. [Google Scholar] [CrossRef] [PubMed]
- MacLeod, A.J.; Rossiter, J.T. Non-enzymic degradation of 2-hydroxybut 3-enylglucosinolate (progoitrin). Phytochemistry 1986, 25, 855–858. [Google Scholar] [CrossRef]
- Borek, V.; Morra, M.J.; Brown, P.D.; McCaffrey, J.P. Allelochemicals produced during sinigrin decomposition in soil. J. Agric. Food Chem. 1994, 42, 1030–1034. [Google Scholar] [CrossRef]
- Donkin, S.G.; Eiteman, M.A.; Williams, P.L. Toxicity of glucosinolates and their enzymatic decomposition products to Caenorhabditis elegans. J. Nematol. 1995, 27, 258–262. [Google Scholar] [PubMed]
- Pratt, C.; Pope, T.W.; Powell, G.; Rossiter, J.T. Accumulation of glucosinolates by the cabbage aphid brevicoryne brassicae as a defense against two coccinellid species. J. Chem. Ecol. 2008, 34, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Kazana, E.; Pope, T.W.; Tibbles, L.; Bridges, M.; Pickett, J.A.; Bones, A.M.; Powell, G.; Rossiter, J.T. The cabbage aphid: A walking mustard oil bomb. Proc. R. Soc. B 2007, 274, 2271–2277. [Google Scholar] [CrossRef] [PubMed]


© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mazumder, A.; Dwivedi, A.; Du Plessis, J. Sinigrin and Its Therapeutic Benefits. Molecules 2016, 21, 416. https://doi.org/10.3390/molecules21040416
Mazumder A, Dwivedi A, Du Plessis J. Sinigrin and Its Therapeutic Benefits. Molecules. 2016; 21(4):416. https://doi.org/10.3390/molecules21040416
Chicago/Turabian StyleMazumder, Anisha, Anupma Dwivedi, and Jeanetta Du Plessis. 2016. "Sinigrin and Its Therapeutic Benefits" Molecules 21, no. 4: 416. https://doi.org/10.3390/molecules21040416
APA StyleMazumder, A., Dwivedi, A., & Du Plessis, J. (2016). Sinigrin and Its Therapeutic Benefits. Molecules, 21(4), 416. https://doi.org/10.3390/molecules21040416