
Chemical Structure and Immunomodulating Activities of an α-Glucan Purified from Lobelia chinensis Lour


 ,
,  ,
,
Abstract
:1. Introduction
2. Results and Discussion
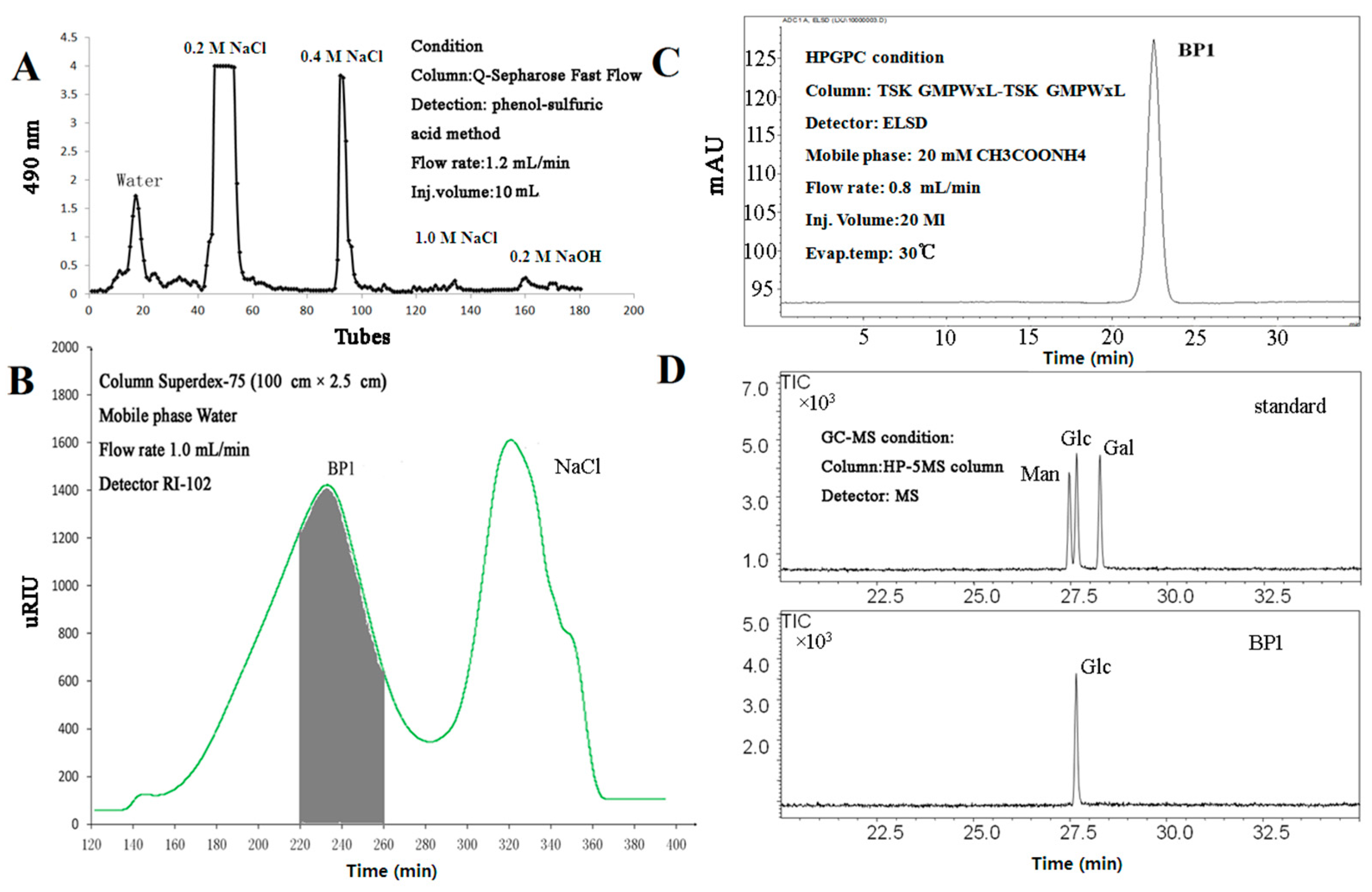
2.1. Isolation and Purification of BP1
2.2. FT-IR Spectral Analysis
2.3. Protein Assay and Monosaccharide Composition Analysis
2.4. Methylation Analysis
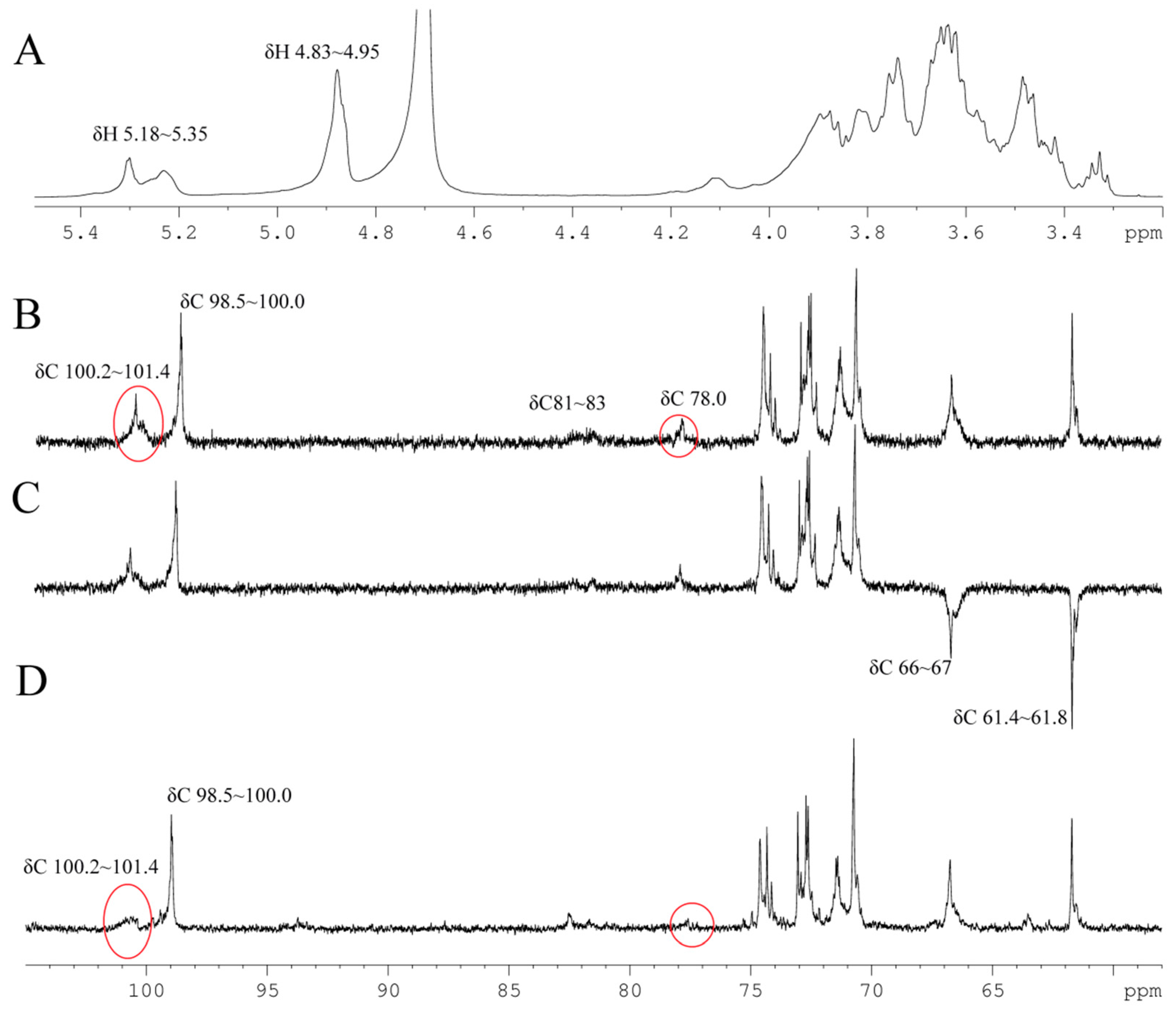
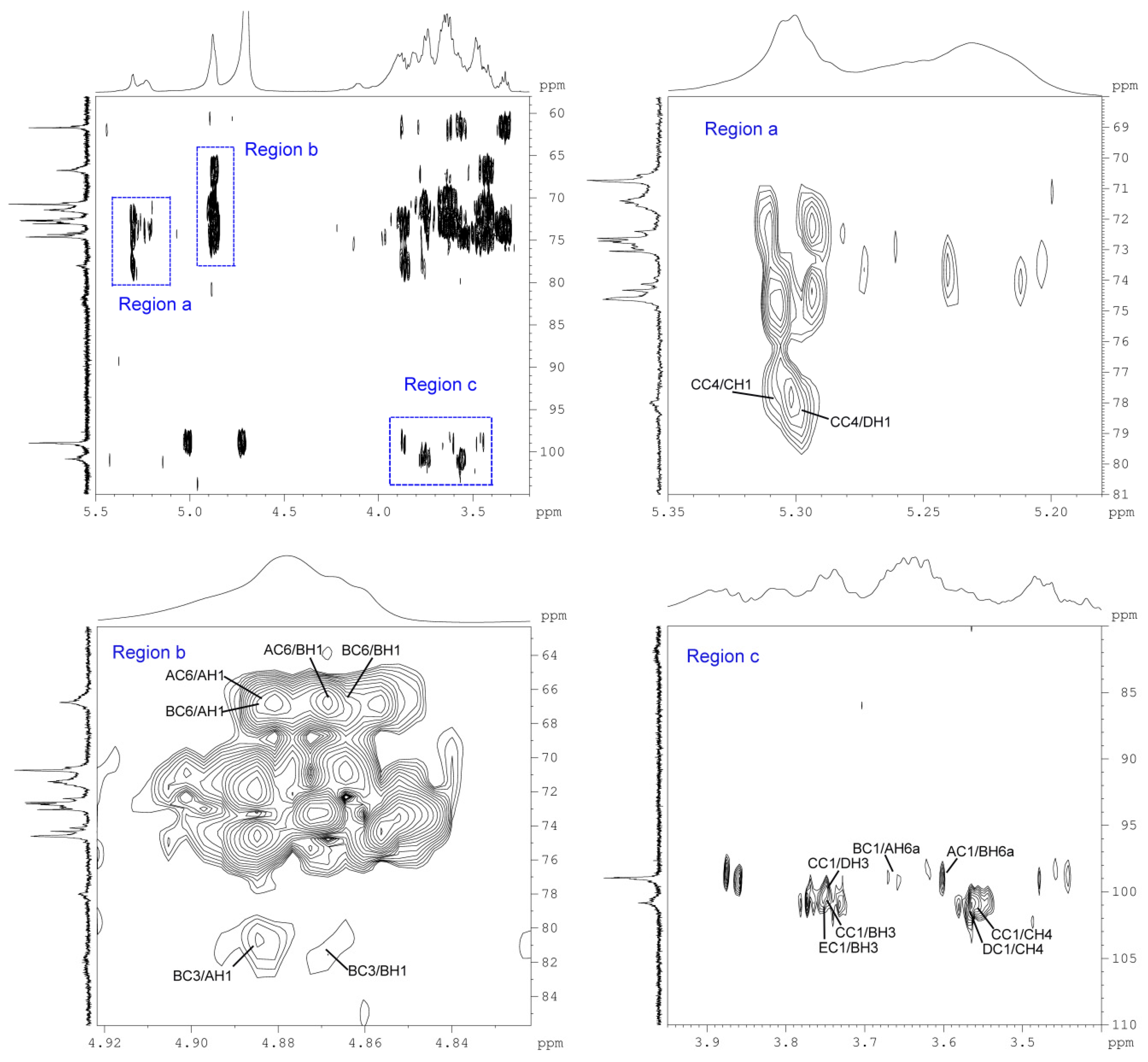
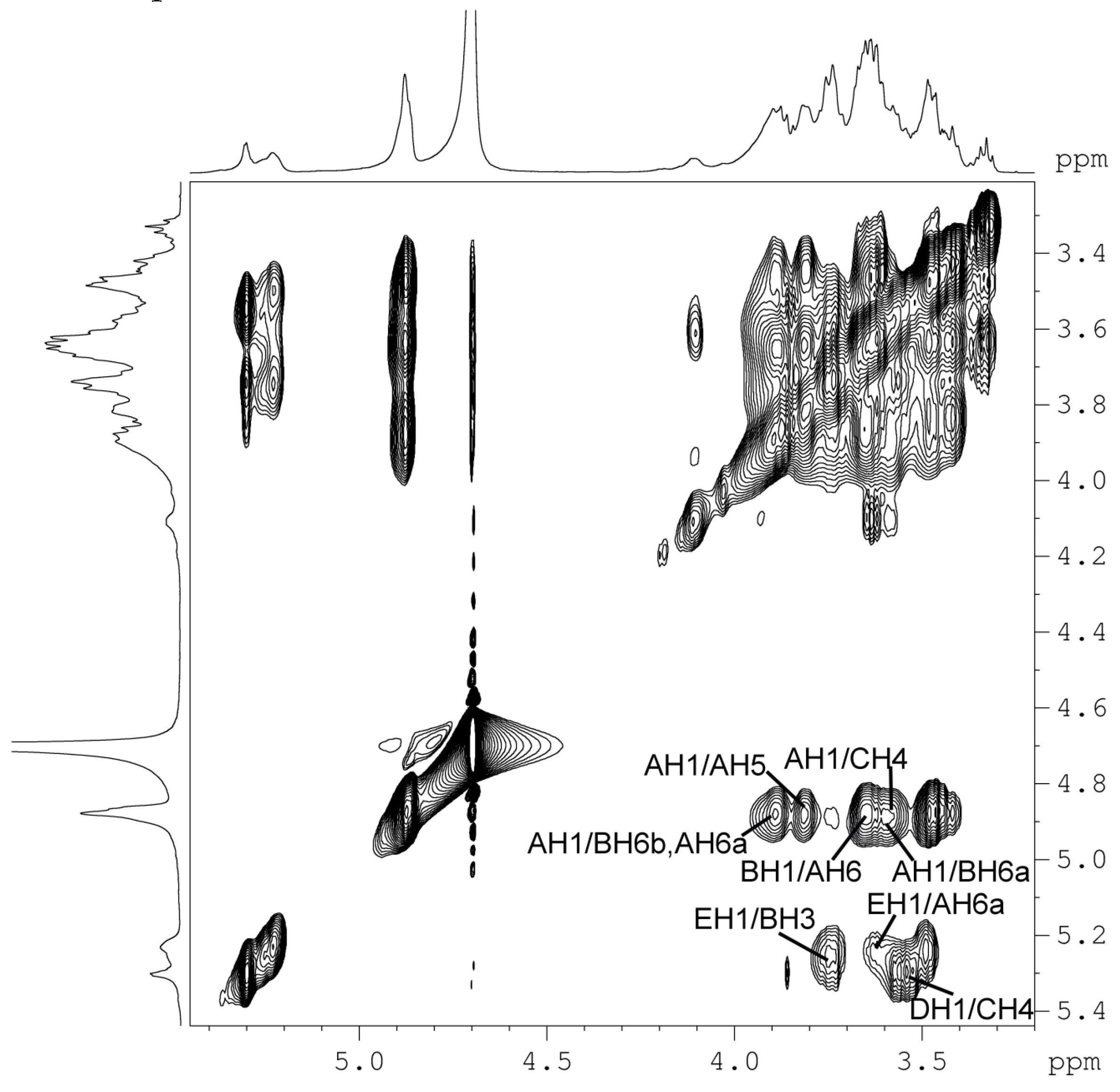
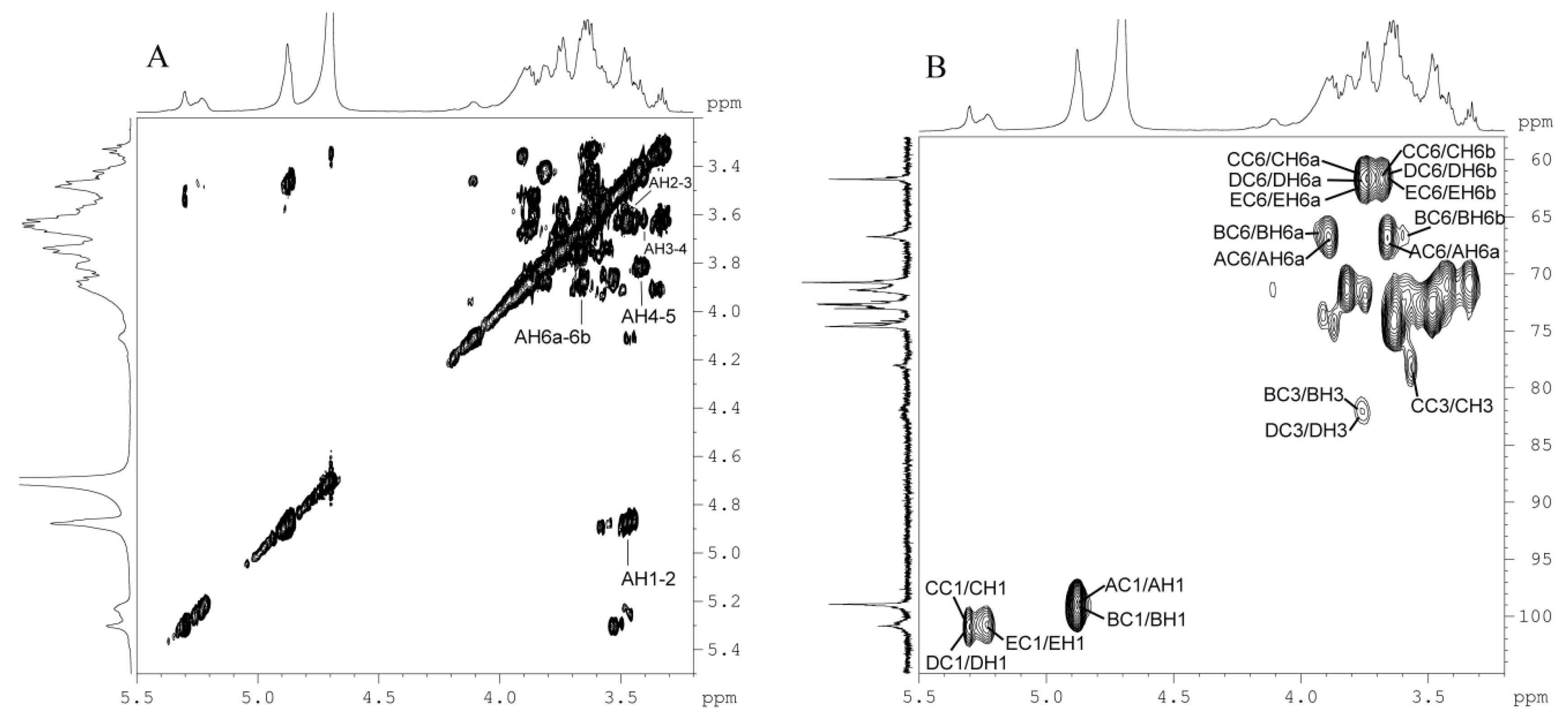
2.5. NMR and Structure Analysis
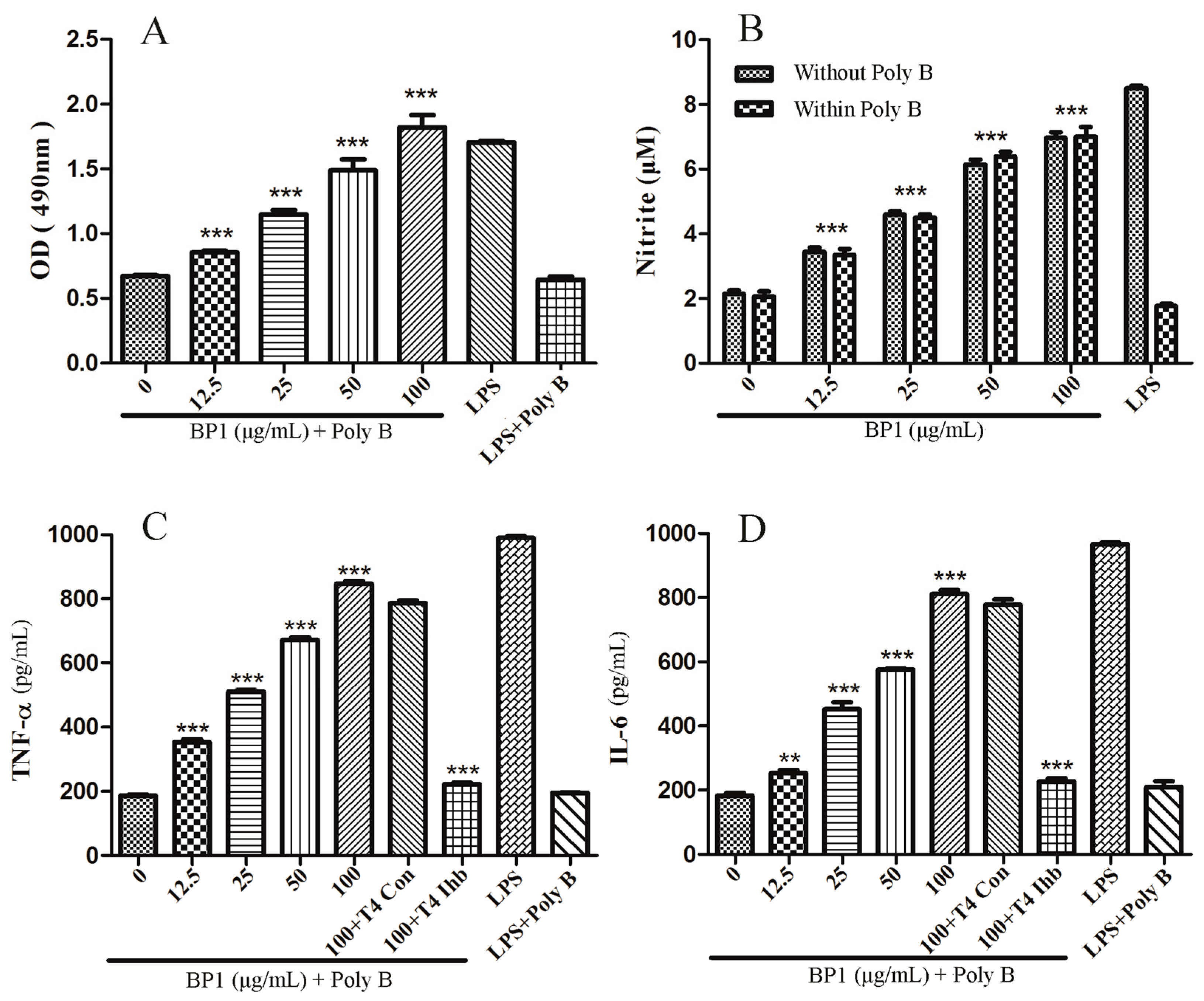
2.6. MTT Assay
2.7. Effect of BP1 on NO Release by RAW 264.7 Cells
2.8. Cytokine Production and the Role of TLR 4
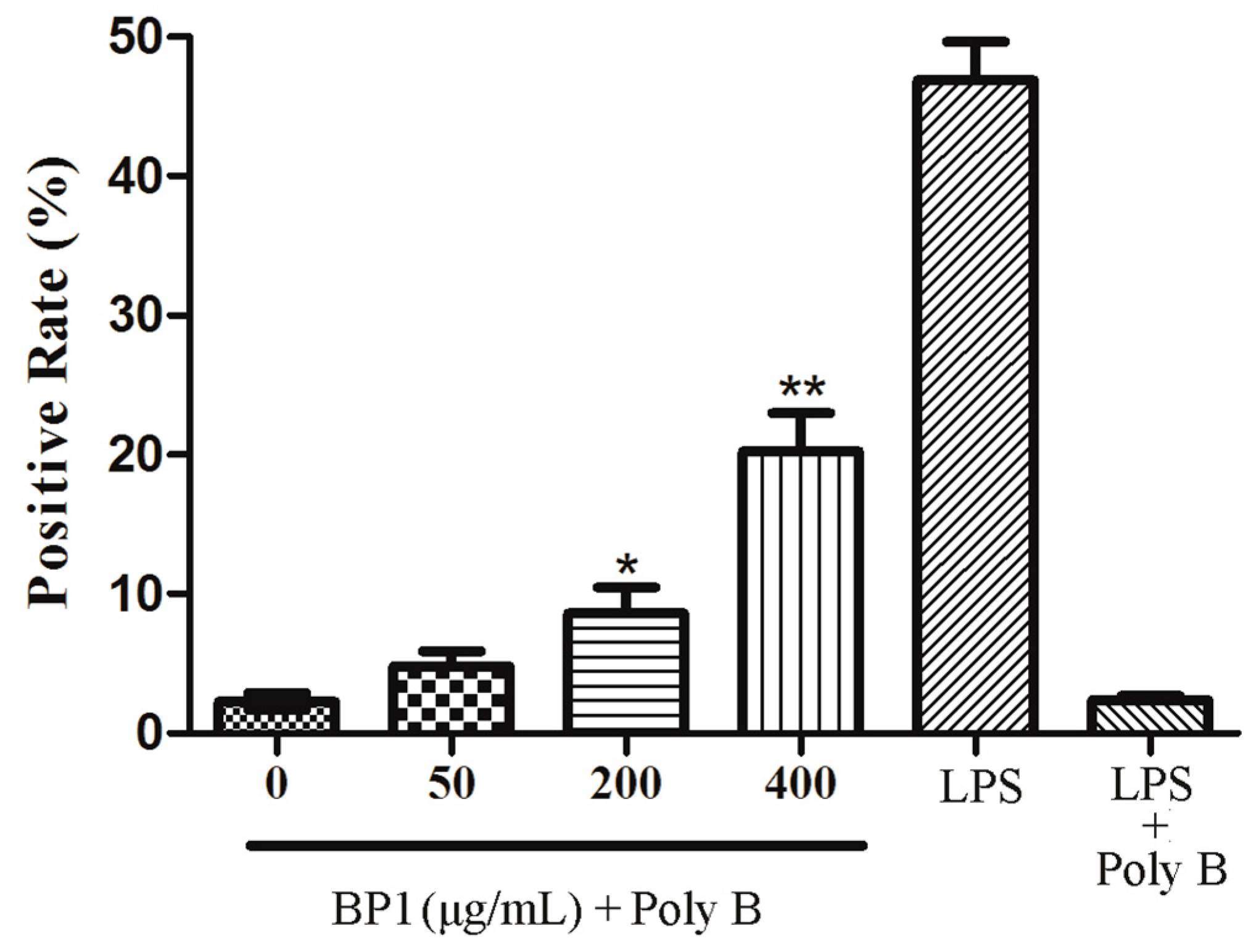
2.9. Assay of Phagocytosis
3. Materials and Methods
3.1. Materials
3.2. Isolation and Purification of BP1
3.3. Determination of the Molecular Weight
3.4. Determination of the Protein Content
3.5. Monosaccharide Composition Analysis
3.6. Methylation Analysis
3.7. Partial Acid Hydrolysis
3.8. NMR Analysis
3.9. MTT Assay
3.10. Nitrite Assay
3.11. Cytokine Determination and the Effect of Toll-Like Receptor 4 Inhibitor
3.12. Phagocytic Assay
3.13. Data Analysis
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Yong, Z.; Lei, Z. Research progress in Lobelia chinensis. Med. Inform. 2006, 19, 1115–1116. [Google Scholar]
- Chen, M.W.; Chen, W.R.; Zhang, J.M.; Long, X.Y.; Wang, Y.T. Lobelia chinensis: Chemical constituents and anticancer activity perspective. Chin. J. Nat. Med. 2014, 12, 103–107. [Google Scholar] [PubMed]
- Yang, S.; Shen, T.; Zhao, L.; Li, C.; Zhang, Y.; Lou, H.; Ren, D. Chemical constituents of Lobelia chinensis. Fitoterapia 2014, 93, 168–174. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Wang, J.X.; Li, Y.Y.; Kang, W.Y. Studies on biological activity of compound recipe of Lobelia chinensis. Chin. J. Exp. Tradit. Med. Form. 2010, 16, 57–61. [Google Scholar]
- Kuo, P.C.; Hwang, T.L.; Lin, Y.T.; Kuo, Y.C.; Leu, Y.L. Chemical constituents from Lobelia chinensis and their anti-virus and anti-inflammatory bioactivities. Arch. Pharm. Res. 2011, 34, 715–722. [Google Scholar] [CrossRef] [PubMed]
- Kuo, Y.C.; Lee, Y.C.; Leu, Y.L.; Tsai, W.J.; Chang, S.C. Efficacy of orally administered Lobelia chinensis extracts on herpes simplex virus type 1 infection in balb/c mice. Antivir. Res. 2008, 80, 206–212. [Google Scholar] [CrossRef] [PubMed]
- Shao, J.H.; Zhang, H. Influence of Lobelia chinensis lour. Decoction on expression of C-erbB-2 and P53 on H22 tumor-bearing mice. Chin. J. Clin. Pharm. 2010, 19, 372–375. [Google Scholar]
- Wang, H.Y.; Quan, K.; Jiang, Y.L.; Wu, J.G.; Tang, X.W. Effect of luteolin and its combination with chemotherapeutic drugs on cytotoxicity of cancer cells. J. Zhejiang Univ. Med. Sci. 2010, 39, 30–36. [Google Scholar]
- Huang, X.X.; Lai, H.F.; Luo, L.C. Optimization of extraction technology for polysaccharide from Lobelia chinensis by ultrasonic-composite enzyme synergistic method. Chin. J. Exp. Tradit. Med. Form. 2012, 18, 44–46. [Google Scholar]
- Yan, J.K.; Wang, W.Q.; Wu, J.Y. Recent advances in Cordyceps sinensis polysaccharides: Mycelial fermentation, isolation, structure, and bioactivities: A review. J. Funct. Foods 2014, 6, 33–47. [Google Scholar] [CrossRef]
- Zhu, F.M.; Du, B.; Bian, Z.X.; Xu, B.J. β-glucans from edible and medicinal mushrooms: Characteristics, physicochemical and biological activities. J. Food Compos. Anal. 2015, 41, 165–173. [Google Scholar] [CrossRef]
- Xie, S.Z.; Hao, R.; Zha, X.Q.; Pan, L.H.; Liu, J.; Luo, J.P. Polysaccharide of Dendrobium huoshanense activates macrophages via toll-like receptor 4-mediated signaling pathways. Carbohydr. Polym. 2016, 146, 292–300. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zou, Y.; Li, Q.; Mao, R.W.; Shao, X.J.; Jin, D.; Zheng, D.H.; Zhao, T.; Zhu, H.F.; Zhang, L.; et al. Immunomodulatory effects of a polysaccharide purified from Lepidium meyenii walp. On macrophages. Process Biochem. 2016, 51, 542–553. [Google Scholar] [CrossRef]
- Huang, J.Q.; Nie, Q.X.; Liu, X.Z.; Zhang, S.S.; Nie, S.P.; Huang, D.F.; Wang, S.A.; Zhu, F.; Xie, M.Y. Ganoderma atrum polysaccharide modulates TNF-α secretion and mRNA expression in macrophages of S-180 tumor-bearing mice. Food Hydrocoll. 2016, 53, 24–30. [Google Scholar] [CrossRef]
- Goo, B.G.; Baek, G.; Choi, D.J.; Park, Y.I.; Synytsya, A.; Bleha, R.; Seong, D.H.; Lee, C.G.; Park, J.K. Characterization of a renewable extracellular polysaccharide from defatted microalgae Dunaliella tertiolecta. Bioresour. Technol. 2013, 129, 343–350. [Google Scholar] [PubMed]
- Wang, L.C.; Zhang, K.; Di, L.Q.; Liu, R.; Wu, H. Isolation and structural elucidation of novel homogenous polysaccharide from Mactra veneriformis. Carbohydr. Polym. 2011, 86, 982–987. [Google Scholar] [CrossRef]
- Zhao, T.; Mao, G.H.; Feng, W.W.; Mao, R.W.; Gu, X.Y.; Li, T.; Li, Q.; Bao, Y.T.; Yang, L.Q.; Wu, X.Y. Isolation, characterization and antioxidant activity of polysaccharide from Schisandra sphenanthera. Carbohydr. Polym. 2014, 105, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.X.; Liu, Q.; Tao, Y.Z.; Zhang, H.F.; Zhang, L.; Ding, K. Structure and chain conformation of a (1→6)-α-d-glucan from the root of Pueraria lobata (willd.) ohwi and the antioxidant activity of its sulfated derivative. Carbohydr. Polym. 2008, 74, 771–778. [Google Scholar] [CrossRef]
- Di, H.; Zhang, Y.; Chen, D. An anti-complementary polysaccharide from the roots of Bupleurum chinense. Int. J. Biol. Macromol. 2013, 58, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Di, H.; Zhang, Y.; Zhang, J.; Chen, D. A protein-bound polysaccharide from the stem bark of Eucommia ulmoides and its anti-complementary effect. Carbohydr. Res. 2009, 344, 1319–1324. [Google Scholar] [CrossRef] [PubMed]
- Han, X.Q.; Wu, X.M.; Chai, X.Y.; Chen, D.; Dai, H.; Dong, H.L.; Ma, Z.Z.; Gao, X.M.; Tu, P.F. Isolation, characterization and immunological activity of a polysaccharide from the fruit bodies of an edible mushroom, Sarcodon aspratus (berk.) s. Ito. Food Res. Int. 2011, 44, 489–493. [Google Scholar] [CrossRef]
- Villares, A.; García-Lafuente, A.; Guillamón, E.; Mateo-Vivaracho, L. Separation and characterization of the structural features of macromolecular carbohydrates from wild edible mushrooms. Bioact. Carbohydr. Diet. Fibre 2013, 2, 15–21. [Google Scholar] [CrossRef]
- Wu, Y.L.; Sun, C.R.; Pan, Y.J. Studies on isolation and structural features of a polysaccharide from the mycelium of an chinese edible fungus (Cordyceps sinensis). Carbohydr. Polym. 2006, 63, 251–256. [Google Scholar]
- Zhao, C.; Li, M.; Luo, Y.F.; Wu, W.K. Isolation and structural characterization of an immunostimulating polysaccharide from fuzi, Aconitum carmichaeli. Carbohydr. Res. 2006, 341, 485–491. [Google Scholar] [CrossRef] [PubMed]
- Carbonero, E.R.; Ruthes, A.C.; Freitas, C.S.; Utrilla, P.; Gálvez, J.; da Silva, E.V.; Sassaki, G.L.; Gorin, P.A.J.; Iacomini, M. Chemical and biological properties of a highly branched β-glucan from edible mushroom Pleurotus sajor-caju. Carbohydr. Polym. 2012, 90, 814–819. [Google Scholar]
- Li, X.J.; Jiang, J.Y.; Shi, S.S.; Li, Y.; Jiang, Y.B.; Ke, Y.; Wang, S.C. Anti-complementary activities of a (1→6) linked glucan from korean mondshood root and its sulfated derivatives. Chem. J. Chin. Univ. Chin. 2014, 35, 1423–1426. [Google Scholar]
- Cordeiro, L.M.; Reinhardt Vde, F.; Iacomini, M. Glucomannan and branched (1→3)(1→6) β-glucan from the aposymbiotically grown Physcia kalbii mycobiont. Phytochemistry 2012, 84, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Dobruchowska, J.M.; Hoogenkamp, M.A.; Gerwig, G.J. Structural investigation of an extracellular polysaccharide produced by the cariogenic bacterium Streptococcus mutans strain ua159. Carbohydr. Polym. 2012, 90, 675–682. [Google Scholar] [CrossRef] [PubMed]
- Lammers, T.G. Revision of the infrageneric classification of Lobelia (Campanulaceae: Lobelioideae). Ann. MO. Bot. Gard. 2011, 98, 37–62. [Google Scholar] [CrossRef]
- Chi, A.P.; Chen, J.P.; Wang, Z.Z.; Xiong, Z.Y.; Li, Q.X. Morphological and structural characterization of a polysaccharide from Gynostemma pentaphyllum makino and its anti-exercise fatigue activity. Carbohydr. Polym. 2008, 74, 868–874. [Google Scholar] [CrossRef]
- Rudd, T.R.; Skidmore, M.A.; Guerrini, M.; Hricovini, M.; Powell, A.K.; Siligardi, G.; Yates, E.A. The conformation and structure of gags: Recent progress and perspectives. Curr. Opin. Struct. Biol. 2010, 20, 567–574. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.Q.; Zhang, L.M. Chemical structural and chain conformational characterization of some bioactive polysaccharides isolated from natural sources. Carbohydr. Polym. 2009, 76, 349–361. [Google Scholar] [CrossRef]
- Brown, G.D.; Gordon, S. Immune recognition. A new receptor for β-glucans. Nature 2001, 413, 36–37. [Google Scholar] [CrossRef] [PubMed]
- Rappleye, C.A.; Eissenberg, L.G.; Goldman, W.E. Histoplasma capsulatum α-(1,3)-glucan blocks innate immune recognition by the β-glucan receptor. Proc. Natl. Acad. Sci. USA 2007, 104, 1366–1370. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yuan, Y.; Yue, T. Immunostimulatory activities of β-d-glucan from Ganoderma lucidum. Carbohydr. Polym. 2014, 102, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Bittencourt, V.C.B.; Figueiredo, R.T.; da Silva, R.B.; Mourão-Sá, D.S.; Fernandez, P.L.; Sassaki, G.L.; Mulloy, B.; Bozza, M.T.; Barreto-Bergter, E. An α-glucan of Pseudallescheria boydii is involved in fungal phagocytosis and toll-like receptor activation. Int. J. Biol. Macromol. Chem. 2006, 281, 22614–22623. [Google Scholar] [CrossRef] [PubMed]
- Zhu, R.; Zhang, X.; Liu, W.; Zhou, Y.; Ding, R.; Yao, W.; Gao, X. Preparation and immunomodulating activities of a library of low-molecular-weight α-glucans. Carbohydr. Polym. 2014, 111, 744–752. [Google Scholar] [CrossRef] [PubMed]
- Dong, Q.; Yao, J.; Fang, J.N.; Ding, K. Structural characterization and immunological activity of two cold-water extractable polysaccharides from Cistanche deserticola y. C. Ma. Carbohydr. Res. 2007, 342, 1343–1349. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Shen, X.; Liao, W.; Fang, J.; Chen, X.; Dong, Q.; Ding, K. A heteropolysaccharide, l-fuco-d-manno-1,6-α-d-galactan extracted from Grifola frondosa and antiangiogenic activity of its sulfated derivative. Carbohydr. Polym. 2014, 101, 631–641. [Google Scholar] [CrossRef] [PubMed]
- Xing, X.H.; Cui, S.W.; Nie, S.P.; Phillips, G.O.; Goff, D.H.; Wang, Q. Study on Dendrobium officinale O-acetyl-glucomannan (Dendronan®): Part II. Fine structures of O-acetylated residues. Carbohydr. Polym. 2015, 117, 422–433. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.W.; Liu, F.; Bligh, S.W.; Shi, S.S.; Wang, S.C. Structure of a homofructosan from Saussurea costus and anti-complementary activity of its sulfated derivatives. Carbohydr. Polym. 2014, 105, 152–160. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Fan, Y.; Wang, W.; Liu, N.; Zhang, H.; Zhu, Z.; Liu, A. Polysaccharides from Lycium barbarum leaves: Isolation, characterization and splenocyte proliferation activity. Int. J. Biol. Macromol. 2012, 51, 417–422. [Google Scholar] [CrossRef] [PubMed]
- Parente, J.P.; Cardon, P.; Leroy, Y.; Montreuil, J.; Fournet, B.; Ricart, G. A convenient method for methylation of glycoprotein glycans in small amounts by using lithium methylsulfinyl carbanion. Carbohydr. Res. 1985, 141, 41–47. [Google Scholar] [CrossRef]
- Duan, J.; Chen, V.L.; Dong, Q.; Ding, K.; Fang, J. Chemical structure and immunoinhibitory activity of a pectic polysaccharide containing glucuronic acid from the leaves of Diospyros kaki. Int. J. Biol. Macromol. 2010, 46, 465–470. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Wang, Z.J.; Sun, G.L.; Shen, L.; Xu, D.S.; Feng, Y. A sensitive and specific hpgpc-fd method for the study of pharmacokinetics and tissue distribution of Radix Ophiopogonis polysaccharide in rats. Biomed. Chromatogr. 2010, 24, 820–825. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Not available.









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Methylation Sugars | Linkages | Retention Time (min) | Molar Ratio | Main Mass Fragment (m/z *, Intensity %) |
|---|---|---|---|---|
| 2,3,4,6-Me4-Glcp | Glc-1 | 28.358 | 2.01 | 43 (100), 71 (20), 87 (25), 101 (65), 117 (50), 129 (48), 145 (40), 161 (45), 205 (10) |
| 2,3,4-Me3-Glcp | Glc-1,6 | 36.061 | 5.97 | 43 (100), 71 (15), 87 (33), 99 (40), 101 (52), 117 (55), 129 (37), 161 (15), 173 (5), 189 (10), 233 (10) |
| 2,3,6-Me3-Glcp | Glc-1,4 | 34.539 | 2.03 | 43 (100), 71 (5), 87 (18), 99 (15), 101 (20), 113 (15), 117 (45), 129 (5), 161 (3), 233 (20) |
| 2,4,6-Me3-Glcp | Glc-1,3 | 33.862 | 0.99 | 43 (100), 71 (16), 87 (25), 99 (15), 101 (38), 113 (5), 117 (85), 129 (70), 161 (30), 173 (5), 233 (10) |
| 2,4-Me2-Glcp | Glc-1,3,6 | 42.231 | 2.01 | 43 (100), 71 (5), 87 (25), 99 (10), 101 (5), 117 (32), 129 (45), 189 (15), 233 (5) |
| Glycosyl Residues | C1 | C2 | C3 | C4 | C5 | C6 | |
|---|---|---|---|---|---|---|---|
| H1 | H2 | H3 | H4 | H5 | H6a | H6b | |
| A: 6α-d-Glcp1 | 99.0 | 73.1 | 74.3 | 70.8 | 71.4 | 66.8 | |
| 4.85~4.90 | 3.44~3.48 | 3.60~3.64 | 3.40~3.43 | 3.80~3.83 | 3.64~3.66 | 3.86~3.90 | |
| B: 3,6α-d-Glcp1 | 98.9 | 72.4 | 81.7 | 74.1 | 70.6 | 66.6 | |
| 4.88~4.91 | 3.54~3.59 | 3.73~3.77 | 3.89~3.93 | 3.33~3.37 | 3.58~3.67 | 3.86~3.90 | |
| C: 4α-d-Glcp1 | 101.0 | 71.4 | 74.6 | 78.0 | 71.6 | 61.8 | |
| 5.29~5.32 | 3.52~3.54 | 3.84~3.88 | 3.55~3.58 | 3.73~3.77 | 3.67~3.69 | 3.73~3.75 | |
| D: 3α-d-Glcp1 | 100.8 | 72.4 | 82.3 | 71.4 | 74.3 | 61.8 | |
| 5.28~5.30 | 3.49~3.50 | 3.73~3.77 | 3.36~3.38 | 3.62~3.66 | 3.67~3.69 | 3.73~3.75 | |
| E: α-d-Glcp1 | 100.6 | 72.7 | 71.4 | 73.9 | 71.5 | 61.6 | |
| 5.22~5.26 | 3.44~3.48 | 4.09~4.12 | 3.89~3.92 | 3.34~3.38 | 3.67~3.69 | 3.73~3.75 | |
| No. | Glycosyl Residues | Atom | Residue | δC | Atom | Residue | δH |
|---|---|---|---|---|---|---|---|
| A | 6α-d-Glcp1 | AH1 | AC3 a | 74.6 | AC1 | AH2 | 3.44~3.48 |
| 4.88 | AH1 | AC5 a | 71.4 | AC1 | AH6a | 3.64~3.66 | |
| AH1 | AC6 | 66.8 | AC1 | BH6a b | 3.56~3.61 | ||
| AH1 | BC3 | 81.7 | AC1 | CC3 | 3.84~3.88 | ||
| AH1 | BC6 b | 66.6 | |||||
| AH1 | CC2 | 71.4 | |||||
| AH1 | CC3 | 74.6 | |||||
| B | 3,6α-d-Glcp1 | BH1 | AC2 | 73.1 | BC1 | AH6a c | 3.64~3.66 |
| 4.87 | BH1 | AC4 | 70.8 | BC1 | BH6a | 3.56~3.61 | |
| BH1 | AC6 c | 66.8 | |||||
| BH1 | BC3 a | 81.7 | |||||
| BH1 | BC5 a | 70.6 | |||||
| BH1 | BC6 | 66.6 | |||||
| C | 4α-d-Glcp1 | CH1 | BC2 | 72.4 | CC1 | DH3 d | 3.73~3.77 |
| 5.31 | CH1 | BC3 e | 74.6 | CC1 | BH3 e | 3.73~3.77 | |
| CH1 | DC2 | 72.4 | CC1 | CH4 | 3.57~3.58 | ||
| CH1 | CC4 | 78.0 | |||||
| D | 3α-d-Glcp1 | DH1 | BC2 | 72.4 | DC1 | CH4 f | 3.57~3.58 |
| 5.29 | DH1 | CC3 | 74.6 | ||||
| DH1 | DC2 | 72.4 | |||||
| DH1 | CC4 f | 78.0 | |||||
| E | α-d-Glcp1 | EH1 | BC4 | 74.1 | EC1 | BH3 g | 3.73~3.77 |
| Anomeric Proton | NOE Contact Proton | δH | Glycosyl Residue |
|---|---|---|---|
| AH1 | AH2 | 3.44~3.48 | |
| AH5 | 3.80~3.83 | ||
| AH6a | 3.64~3.66 | ||
| AH6b | 3.88~3.90 | ||
| BH6a | 3.59~3.61 | →6α-d-Glcp1→6,3α-d-Glcp1→ | |
| BH6b | 3.88~3.90 | →6α-d-Glcp1→6,3α-d-Glcp1→ | |
| CH4 | 3.57~3.58 | →6α-d-Glcp1→4α-d-Glcp1→ | |
| BH1 | BH2 | 3.54~3.59 | |
| BH3 | 3.73~3.77 | ||
| CH1 | CH2 | 3.52~3.54 | |
| CH3 | 3.73~3.77 | ||
| BH3 | 3.73~3.77 | ||
| DH1 | CH4 | 3.57~3.58 | →3α-d-Glcp1→4α-d-Glcp1→ |
| DH1 | CH3 | 3.84~3.88 | |
| EH1 | BH3 | 3.73~3.77 | α-d-Glcp1→3,6α-d-Glcp1→ |
| AH2 | 3.44~3.48 | ||
| AH3 | 3.60~3.64 | ||
| AH6a | 3.64~3.66 | α-d-Glcp1→6α-d-Glcp1→ | |
| AH6b | 3.88~3.90 | α-d-Glcp1→6α-d-Glcp1→ |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.-J.; Bao, W.-R.; Leung, C.-H.; Ma, D.-L.; Zhang, G.; Lu, A.-P.; Wang, S.-C.; Han, Q.-B. Chemical Structure and Immunomodulating Activities of an α-Glucan Purified from Lobelia chinensis Lour. Molecules 2016, 21, 779. https://doi.org/10.3390/molecules21060779
Li X-J, Bao W-R, Leung C-H, Ma D-L, Zhang G, Lu A-P, Wang S-C, Han Q-B. Chemical Structure and Immunomodulating Activities of an α-Glucan Purified from Lobelia chinensis Lour. Molecules. 2016; 21(6):779. https://doi.org/10.3390/molecules21060779
Chicago/Turabian StyleLi, Xiao-Jun, Wan-Rong Bao, Chung-Hang Leung, Dik-Lung Ma, Ge Zhang, Ai-Ping Lu, Shun-Chun Wang, and Quan-Bin Han. 2016. "Chemical Structure and Immunomodulating Activities of an α-Glucan Purified from Lobelia chinensis Lour" Molecules 21, no. 6: 779. https://doi.org/10.3390/molecules21060779
APA StyleLi, X. -J., Bao, W. -R., Leung, C. -H., Ma, D. -L., Zhang, G., Lu, A. -P., Wang, S. -C., & Han, Q. -B. (2016). Chemical Structure and Immunomodulating Activities of an α-Glucan Purified from Lobelia chinensis Lour. Molecules, 21(6), 779. https://doi.org/10.3390/molecules21060779