
Structural Elucidation of a Novel Polysaccharide from Pseudostellaria heterophylla and Stimulating Glucose Uptake in Cells and Distributing in Rats by Oral


Abstract
:1. Introduction
2. Results and Discussion
2.1. Isolation and Purification of Pseudostellaria heterophylla Polysaccharides
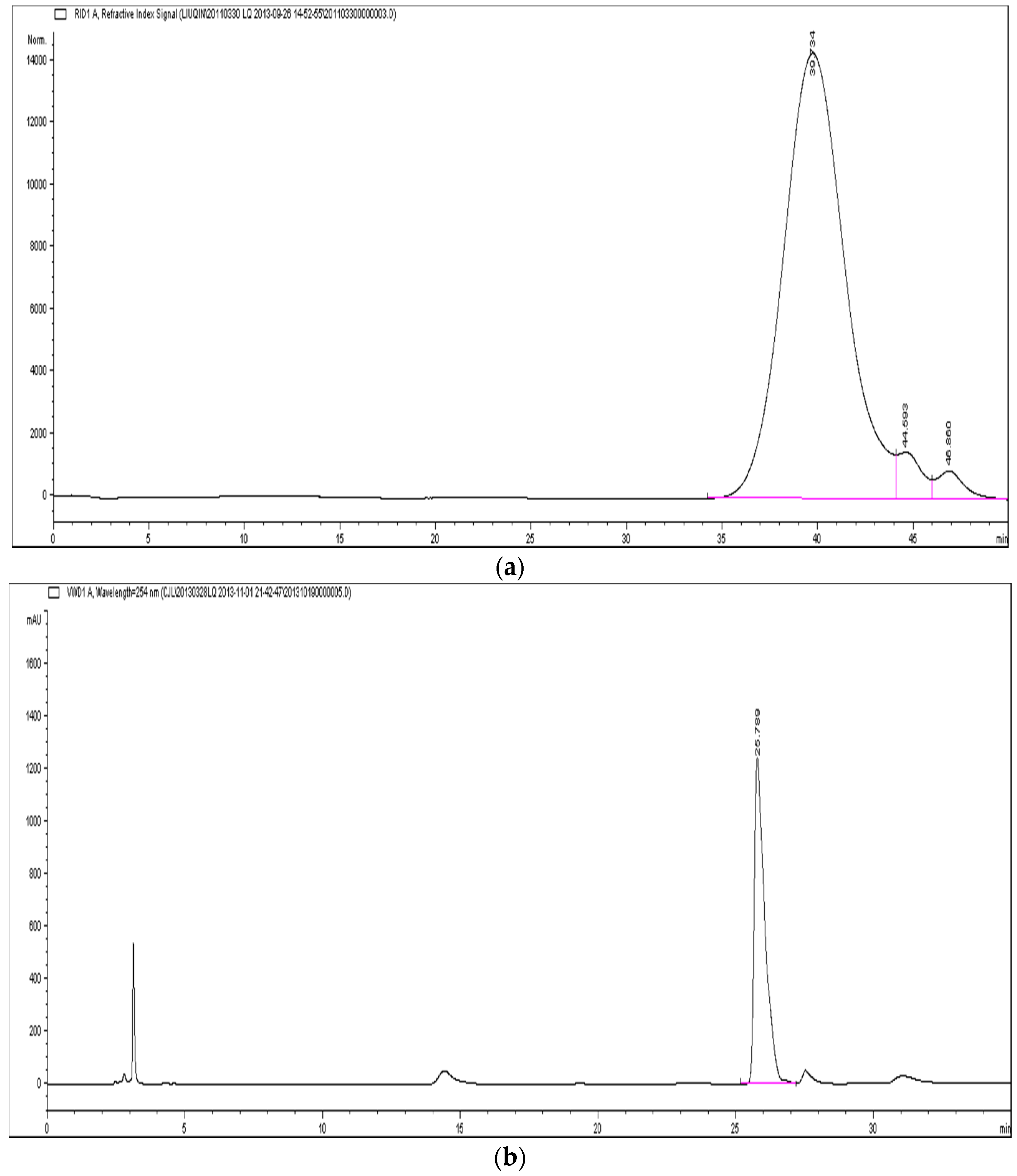
2.2. Molecular Weight, Specific Optical Rotation and Monosaccharide Composition Analysis
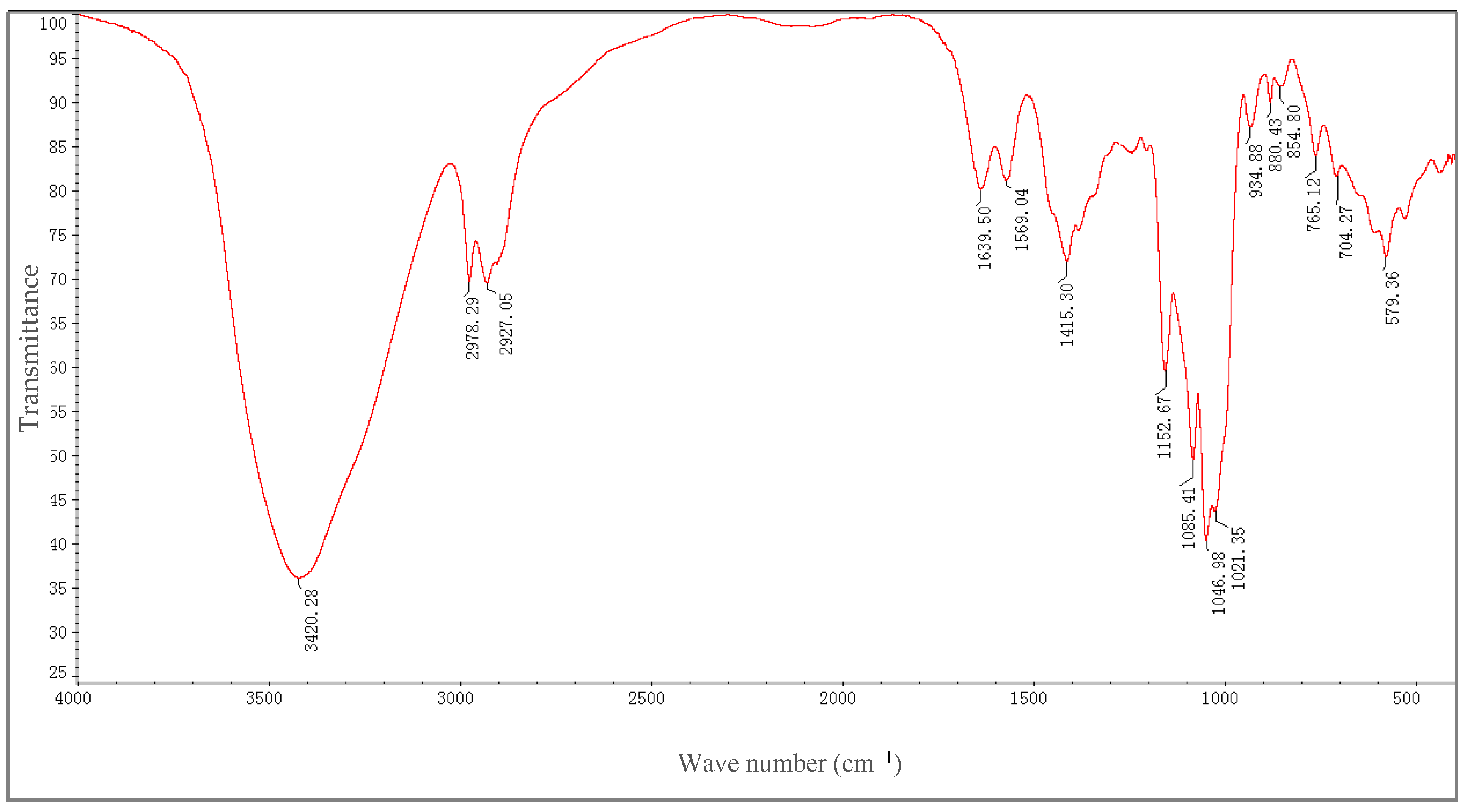
2.3. FT-IR Spectra Analysis
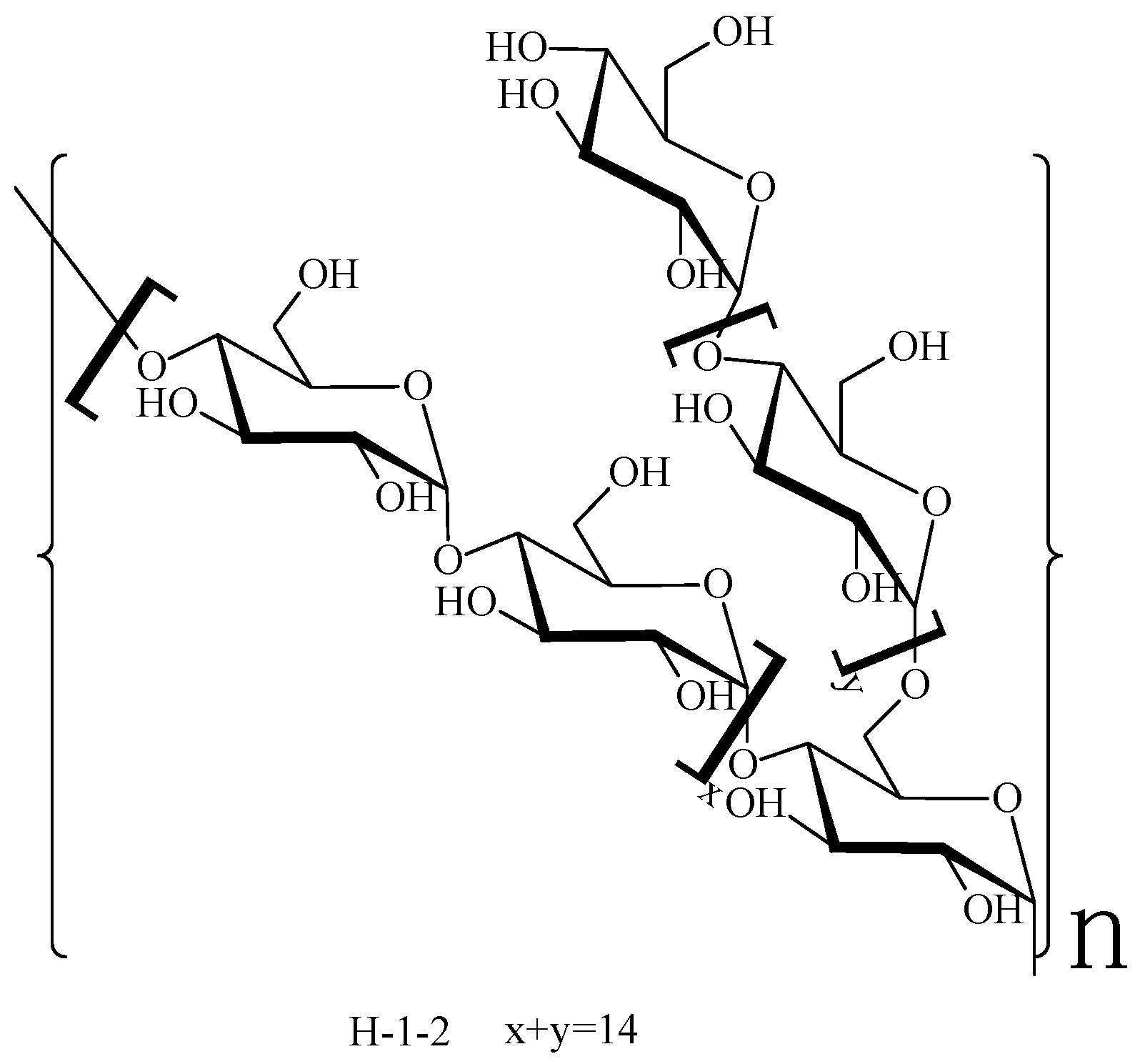
2.4. Methylation Analysis
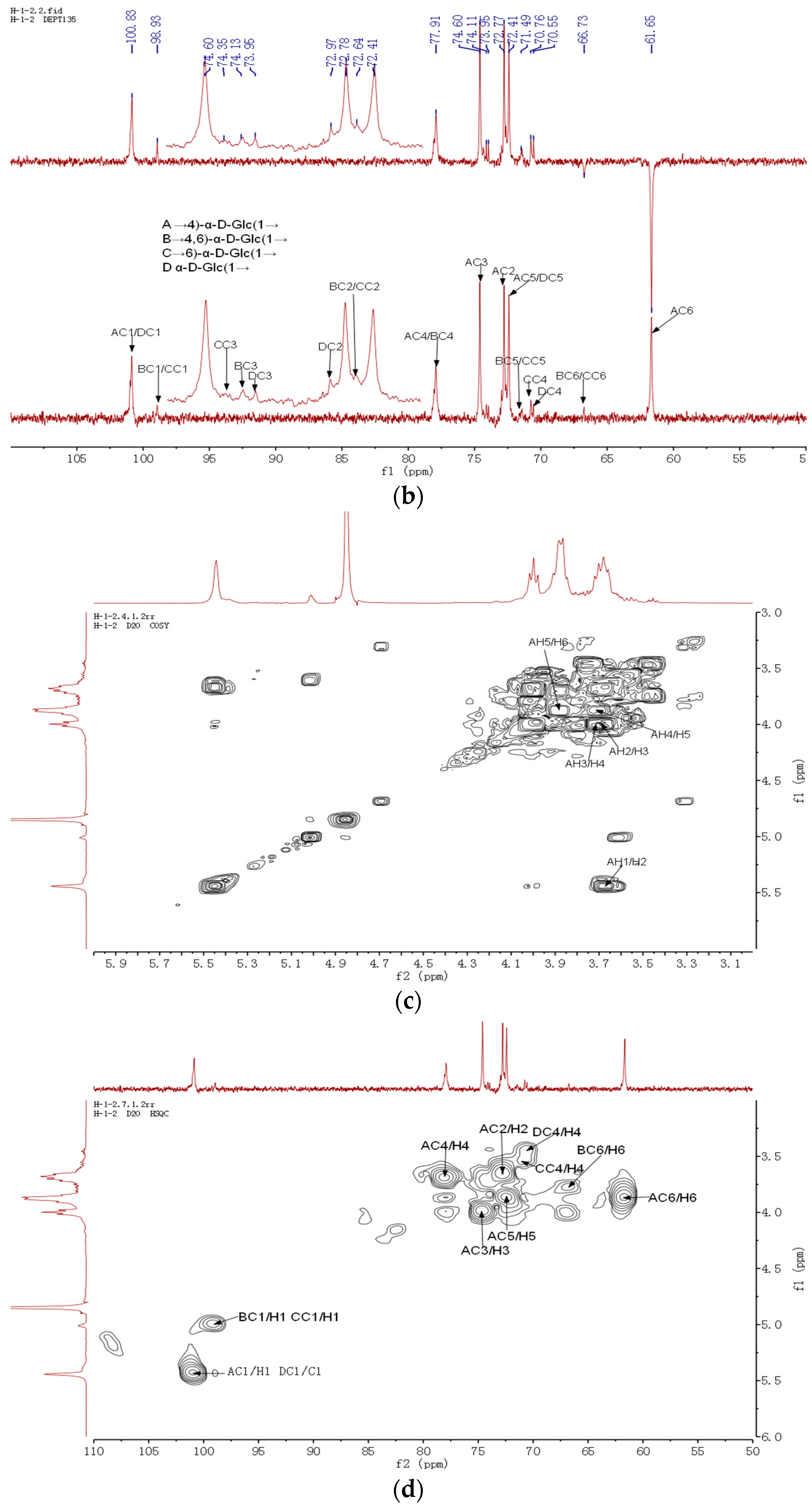
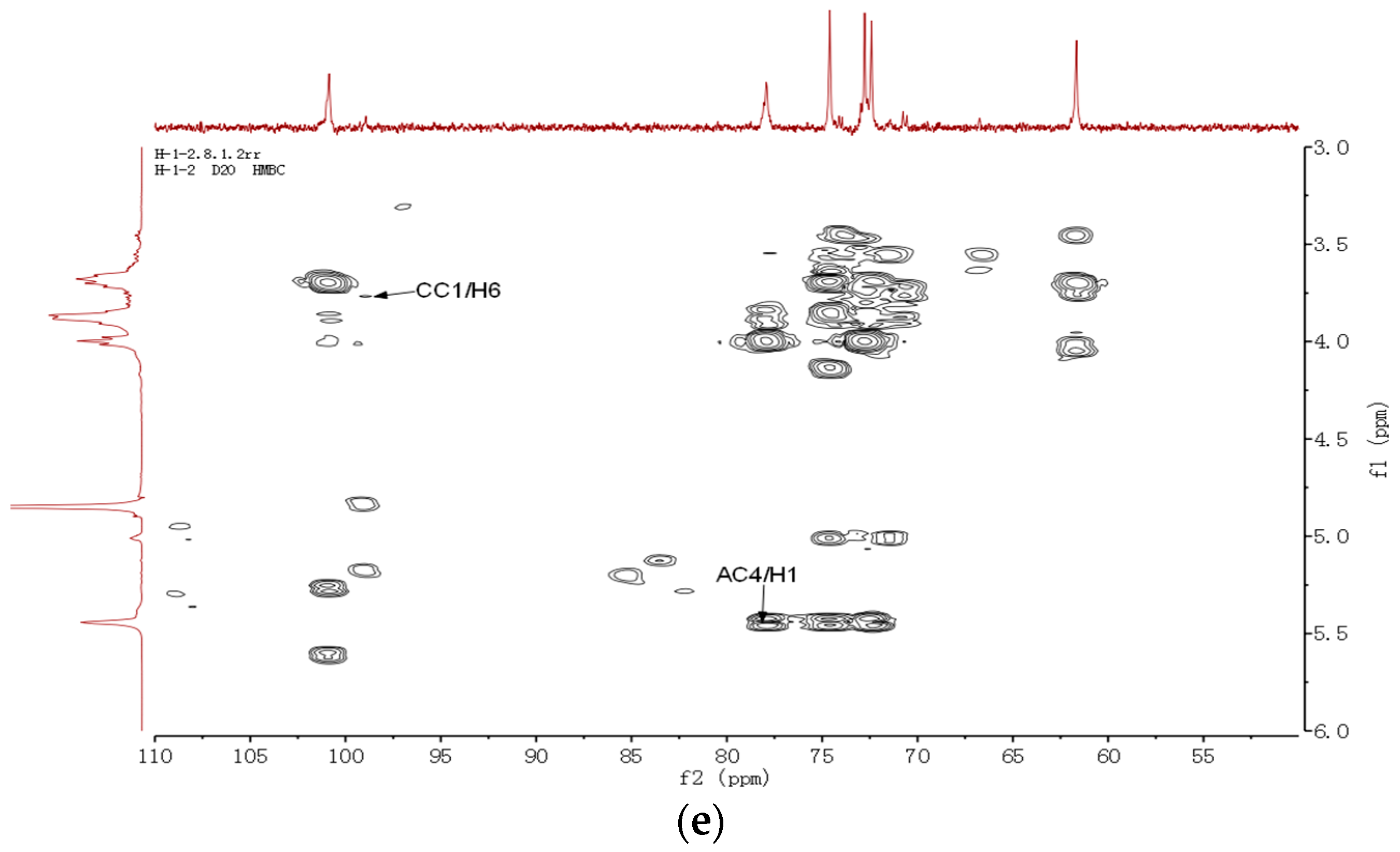
2.5. Nuclear Magnetic Resonance Spectroscopy Analysis
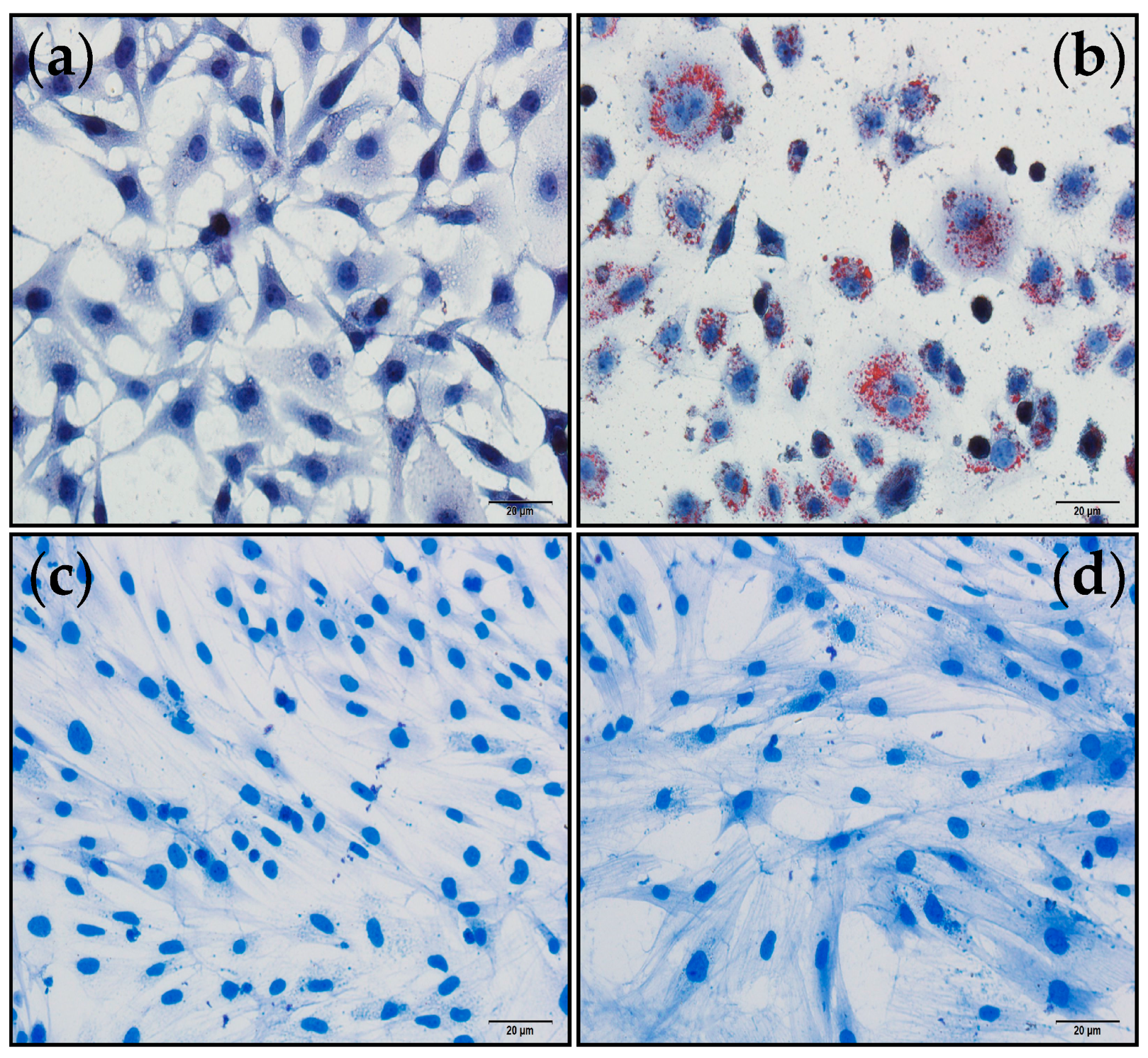
2.6. Assessment of the Anti-Diabetic Activities of Polysaccharide H-1-2 Using Cultured Cell Models
2.6.1. Effects of H-1-2 on Cellular Glucose Consumption
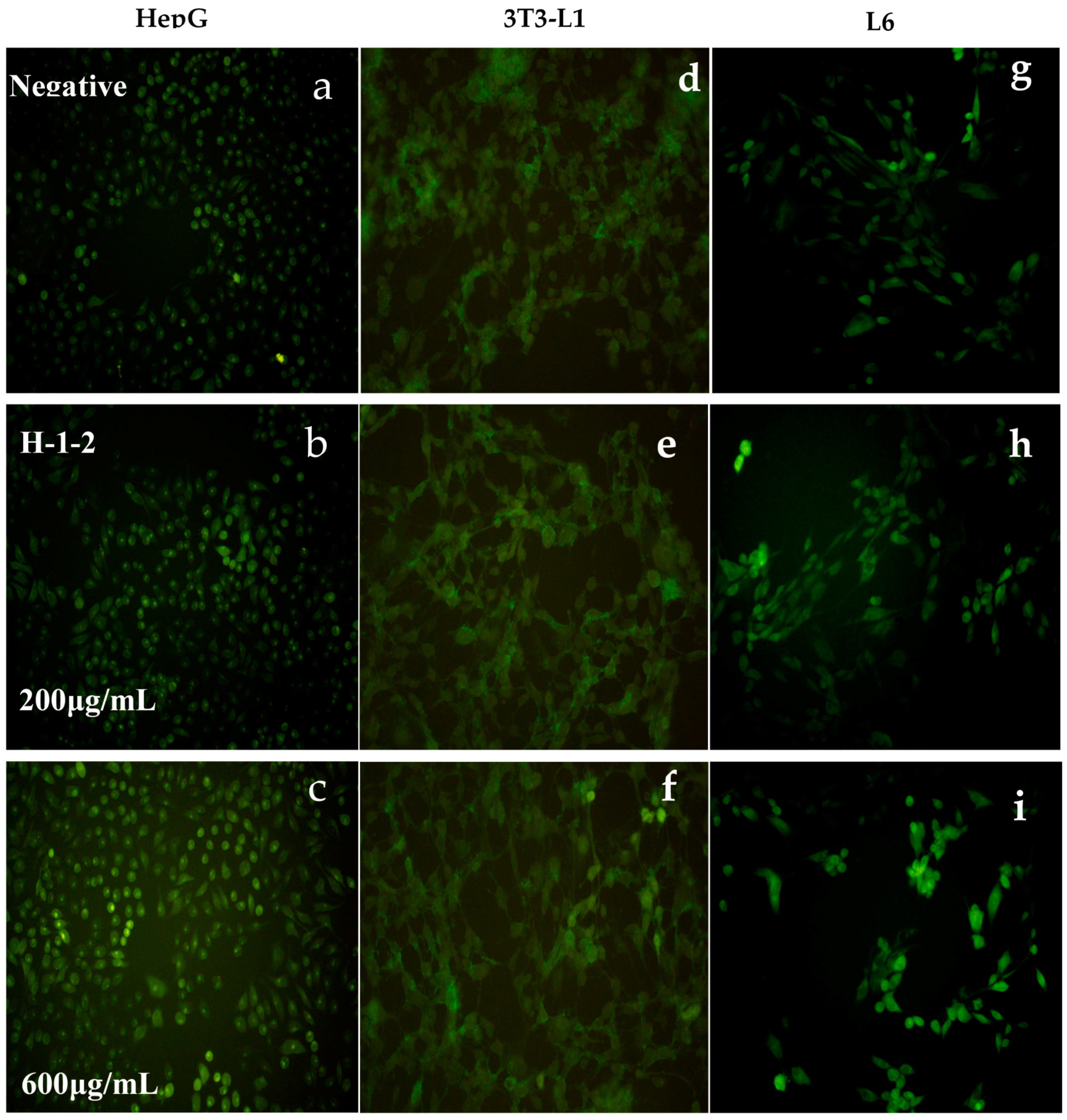
2.6.2. Effects of H-1-2 on Glucose Uptake
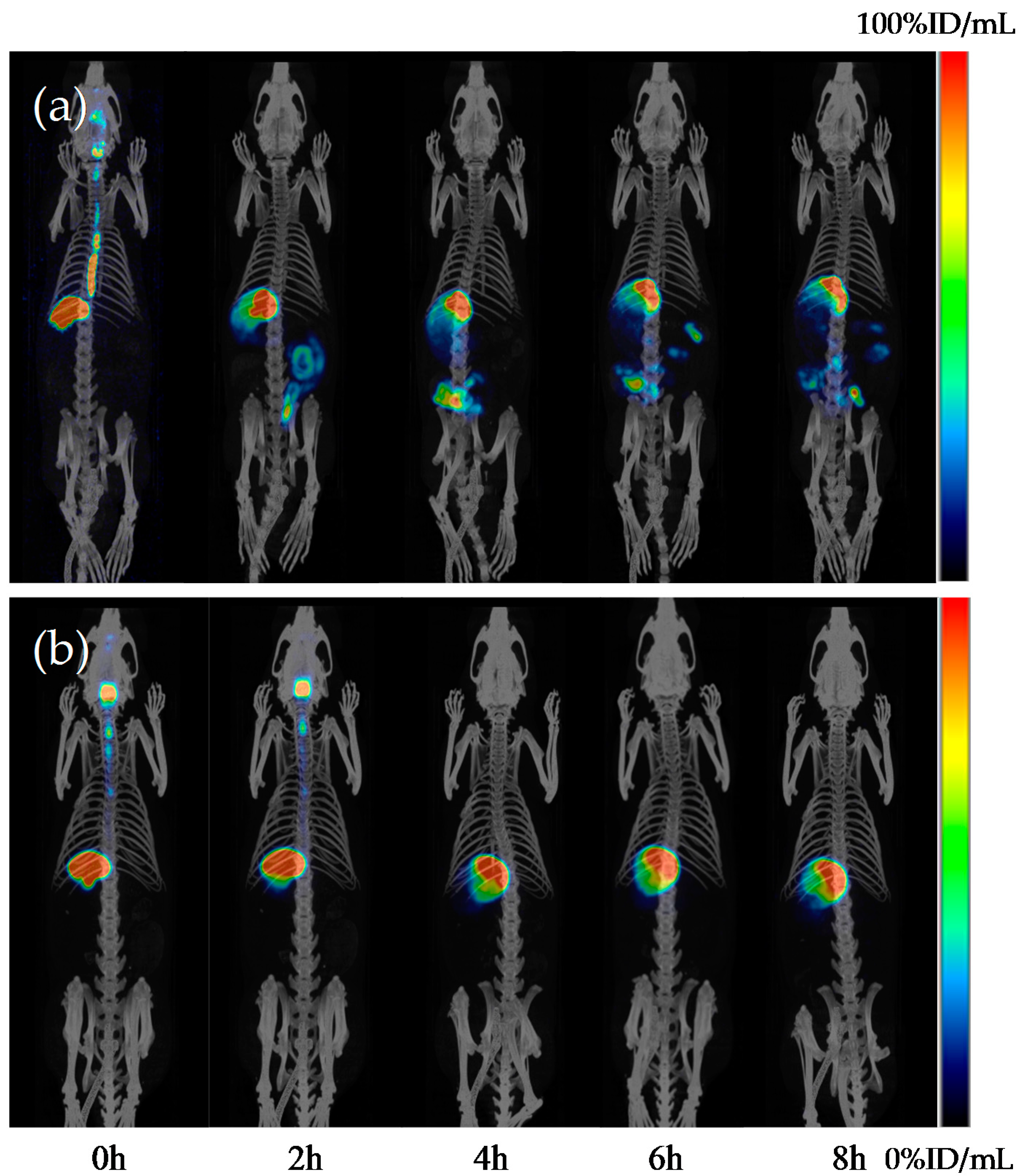
2.7. Distribution of H-1-2 Taken Orally in Abdominal Organs in Rat
3. Experimental Section
3.1. Plant Materials
3.2. Chemicals
3.3. Cells
3.4. Isolation and Purification of Pseudostellaria heterophylla Polysaccharides
3.5. Analytical Methods
3.5.1. Polysaccharide Content, Monosaccharide Composition, and Molecular Weight Distribution Determination
3.5.2. Sequence of Polysaccharide Determination
Periodate Oxidation
Smith Hydrolysis
Methylation Analysis
3.5.3. Polysaccharide Structure Analysis
Fourier Transforms Infrared Analysis
Nuclear Magnetism Spectra Analysis
3.6. Determination of Glucose Consumption and Glucose Uptake in Cells
3.7. Animal Study—Biodistribution of H-1-2
3.8. Statistical Analyses
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Pharmacopoeia Commission of the Ministry of Health of the People’s Republic of China. Pharmacopoeia of the People’s Republic of China; Medical Science and Technology Press: Beijing, China, 2015; Volume 1, p. 68.
- Pang, W.S.; Lin, S.D.; Dai, Q.W.; Zhang, H.C.; Hu, J. Antitussive activity of Pseudostellaria heterophylla (Miq.) Pax extracts and improvement in lung function via adjustment of multi-cytokine levels. Molecules 2011, 16, 3360–3370. [Google Scholar] [CrossRef] [PubMed]
- Ni, Q.; Chen, S.B.; Zhou, X.Z.; Wei, Z.X.; Gao, Y.B.; Li, Y.; Wang, X.M.; Li, P.; Lin, L.; Liu, B.Y. Study of relationship between formula (herbs) and syndrome about type 2 diabetes melliftus affiliated metablic syndrome based on the free-scale network. Chin. J. Inf. Tradit. Chin. Med. 2006, 13, 19–22. [Google Scholar]
- Gong, Z.; Dai, Y.; Ma, H.; Wang, Z.; Yu, G. The effect of Radix Pseudostellariae from 8 habitats on spleen-deficiency and immunologic function. Zhong Yao Cai 2001, 24, 281–282. [Google Scholar] [PubMed]
- Wong, C.K.; Leung, K.N.; Fung, K.P.; Choy, Y.M. The immunostimulating activities of anti-tumor Polysaccharides from Pseudostellaria heterophylla. Immunopharmacology 1994, 28, 47–54. [Google Scholar] [CrossRef]
- Cai, J.; Li, X.D.; Chen, X.Z.; Zheng, L.P.; Lin, J.M. The immune effects of crude extract of Pseudostellaria polysaccharide in mice. J. Fujian Coll. Tradit. Chin. Med. 2005, 15, 33–35. [Google Scholar]
- Xia, L.Z.; Xu, X.X.; Zhang, R. Effects of Pseudostellaria polysaccharides on glucose and lipid metabolism in diabetic rats. China Pharm. 2009, 18, 17–18. [Google Scholar]
- Bao, C. Effects of Pseudostellaria polysaccharides on glucose and lipid metabolism in diabetic mice. Chin. Arch. Tradit. Chin. Med. 2010, 28, 2195–2196. [Google Scholar]
- Xu, X.X.; Huang, Y.X.; Xia, L.Z.; Zhang, R. Effect of Pseudostellaria heterophylla polysaccharides on anti-oxidation capacity and pancreatic pathology of diabetic mouse. Sci. Technol. Food Ind. 2012, 34, 392–393. [Google Scholar]
- Xu, J.L.; Xu, A.R.; Ying, J.Y.; Ma, W.C.; Chen, Y.N. Effect of Pseudostellaria polysaccharides on glucose and insulin metabolism on diabetic rats. Chin. Arch. Tradit. Chin. Med. 2012, 30, 423–424. [Google Scholar]
- Hu, J.; Pang, W.S.; Chen, J.L.; Bai, S.W.; Zheng, Z.Z.; Wu, X.H. Hypoglycemic effect of polysaccharides with different molecular weight of Pseudostellaria heterophylla. BMC Complement. Altern. Med. 2013, 13, 267. [Google Scholar] [CrossRef] [PubMed]
- Yea, K.; Kim, J.; Yoon, J.H.; Kwon, T.; Kim, J.H.; Lee, B.D.; Lee, H.J.; Lee, S.J.; Kim, J.I.; Lee, T.G.; et al. Lysophosphatidylcholine activates adipocyte glucose uptake and lowers blood glucose levels in murine models of diabetes. J. Biol. Chem. 2009, 284, 33833–33840. [Google Scholar] [CrossRef] [PubMed]
- Lau, W.K.; Goh, B.H.; Kadir, H.A.; Shu-Chien, A.C.; Muhammad, T.S. Potent PPARγ ligands from Swietenia macrophylla are capable of stimulating glucose uptake in muscle cells. Molecules 2015, 20, 22301–22314. [Google Scholar] [CrossRef] [PubMed]
- Cao, W.; Li, X.Q.; Liu, L.; Wang, M.; Fan, H.T.; Li, C.; Lv, Z.; Wang, X.; Mei, Q. Structural analysis of water-soluble glucans from the root of Angelica sinensis (Oliv.) Diels. Carbohydr. Res. 2006, 341, 1870–1877. [Google Scholar] [CrossRef] [PubMed]
- Chandra, K.; Ghosh, K.; Roy, S.K.; Mondal, S.; Maiti, D.; Ojha, A.K.; Das, D.; Mondal, S.; Islam, S.S. A water-soluble glucan isolated from an edible mushroom Termitomyces microcarpus. Carbohydr. Res. 2007, 342, 2484–2489. [Google Scholar] [CrossRef] [PubMed]
- Qiu, H.; Tang, W.; Tong, X.; Ding, K.; Zuo, J. Structure elucidation and sulfated derivatives preparation of two α-d-glucans from Gastrodia elata Bl and their anti-dengue virus bioactivities. Carbohydr. Res. 2007, 342, 2230–2236. [Google Scholar] [CrossRef] [PubMed]
- Van Leeuwen, S.S.; Kralj, S.; van Geel-Schutten, I.H.; Gerwig, G.J.; Dijkhuizen, L.; Kamerling, J.P. Structural analysis of the α-d-glucan (EPS180) produced by the Lactobacillus reuteri strain 180 glucansucrase GTF180 enzyme. Carbohydr. Res. 2008, 343, 1237–1250. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, C.; Jansson, P.E.; Widmalm, G.; Sørensen, U.B.S. Structural elucidation of the capsular polysaccharide from Strep tococcus pneumoniae type 18B. Carbohydr. Res. 1997, 304, 165–172. [Google Scholar] [CrossRef]
- Fang, J.N. The application of C13-NMR in the structure analysis of polysaccharide. World Notes Antibiot. 1982, 3, 107–112. [Google Scholar]
- Yki-Järvinen, H. Fat in the liver and insulin resistance. Ann. Med. 2005, 37, 347–356. [Google Scholar] [CrossRef] [PubMed]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Shi, W.T.; Pang, W.S.; Hu, J. Determination of Molecular Size of Pseudostellaria heterophylla polysaccharide by HPGPC. Chin. J. Ethnomed. Ethnopharm. 2015, 20, 20–30. [Google Scholar]
- Qiao, D.L.; Liu, J.; Ke, C.L.; Sun, Y.; Ye, H.; Zeng, X.X. Structural characterization of polysaccharides from Hyriopsis cumingii. Carbohydr. Polym. 2010, 824, 1184–1190. [Google Scholar] [CrossRef]
- Jian, L.J.; Chang, J.M.; Ablise, M.; Li, G.R.; He, J.W. Isolation, purification, and structural elucidation of polysaccharides from Alhagi-honey. J. Asian Natl. Prod. Res. 2014, 16, 783–789. [Google Scholar] [CrossRef] [PubMed]
- Needs, P.W.; Selvendran, R.R. Avoiding oxidative degradation during sodium hydroxide/methyl iodide-mediated carbohydrate methylation indimethyl sulfoxide. Carbohydr. Res. 1993, 245, 1–10. [Google Scholar] [CrossRef]
- Gnanasambandam, R.; Proctor, A. Determination of pectin degree of ester-ification by diffuse reflectance Fourier transform infrared spectroscopy. Food Chem. 2000, 68, 327–332. [Google Scholar] [CrossRef]
- Cai, W.R.; Xie, L.L.; Chen, Y.; Zhang, H. Purification, characterization and anticoagulant activity of the polysaccharides from green tea. Carbohydr. Polym. 2013, 92, 1086–1090. [Google Scholar] [CrossRef] [PubMed]
- Tamaki, Y.; Konishi, T.; Fukuta, M.; Tako, M. Isolation and structural characterization of pectin from endocarp of Citrus depressa. Food Chem. 2008, 107, 352–361. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Zhao, X.Y.; Lu, H.Z.; Wu, Y.T.; Sun, Z.; Liu, Y.Z.; Wang, Y.M.; Sang, Q.; Liu, X.Y. Effects of zinc on cell viability and glucose consumption of insulin-resistant cells. Chin. J. Public Health 2015, 31, 461–464. [Google Scholar]
- Masaaki, S.; Nodi, D.; Masaaki, S.; Anette, I.Ö.; Roger, J.S.; Dana, S.H.; Tore, B. Response to comment on Sato et al. Improving type 2 diabetes through a distinct adrenergic signaling pathway involving mTORC2 that mediates glucose uptake in skeletal muscle. Diabetes 2014, 63, 4115–4129. [Google Scholar]
- Banerjee, T.; Singh, A.K.; Sharma, R.K.; Maitra, A.N. Labeling efficiency and biodistribution of Technetium-99m labeled nanoparticles: Interference by colloidal tin oxide particles. Int. J. Pharm. 2005, 289, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the compounds are not available from the authors.












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Methylated Glycosyl | Retention Time | Mole Ratio | Mass Charge Ratio (m/z) | Link Mode |
|---|---|---|---|---|
| 2,3,4,6-Me4-Glc | 14.275 | 1 | 43, 71, 87, 101, 117, 129, 145, 161, 205 | Glc(1→ |
| 2,3,6-Me3-Glc | 18.522 | 14 | 43, 71, 87, 99, 101, 117, 129, 161, 233 | →4)Glc(1→ |
| 2,3,4-Me3-Glc | 19.075 | 1.5 | 43, 71, 87, 101, 117, 129, 161, 189, 233 | →6)Glc(1→ |
| 2,3-Me2-Glc | 22.958 | 1.2 | 43, 85, 99, 101, 117, 127, 159, 201, 261 | →4,6)Glc(1→ |
| Glycosy Link Mode | Chemical Shift (ppm) | |||||
|---|---|---|---|---|---|---|
| H-1/C-1 | H-2/C-2 | H-3/C-3 | H-4/C-4 | H-5/C-5 | H-6/C-6 | |
| Glc(1→ | 5.44/100.83 | 3.66/72.97 | 3.98/73.95 | 3.45/70.55 | 4.02/72.41 | 3.88/61.65 |
| →4)Glc(1→ | 5.44/100.83 | 3.70/72.77 | 4.0/74.60 | 3.68/77.91 | 3.86/72.41 | 3.88/61.65 |
| →6)Glc(1→ | 5.01/98.93 | 3.63/72.64 | 3.75/74.35 | 3.54/70.76 | 3.93/71.49 | 3.78/66.73 |
| →4,6)Glc(1→ | 5.01/98.93 | 3.55/72.64 | 3.70/74.13 | n.d./77.91 | n.d./71.49 | 3.78/66.73 |
| Cells | HepG2 | 3T3L1 | L6 | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Group (μg/mL) | Escalating Rate (%) | |||||||||
| 12 h | 24 h | 48 h | 12 h | 24 h | 48 h | 12 h | 24 h | 48 h | ||
| H-1-2 | 50 | 8.06 | 9.92 | 10.37 | 2.52 | 4.68 | 6.17 | 3.78 | 3.92 | 7.73 |
| 100 | 12.20 | 14.06 | 16.93 | 5.21 | 5.90 | 9.11 | 4.81 | 5.30 | 7.84 | |
| 200 | 15.77 | 17.03 | 22.88 | 5.80 | 9.32 | 12.10 | 5.94 | 7.23 | 10.91 | |
| 400 | 18.09 | 20.39 | 23.96 | 8.78 | 10.22 | 15.16 | 8.41 | 8.66 | 12.29 | |
| 500 | 19.89 | 21.74 | 26.11 | 9.75 | 11.91 | 16.44 | 9.16 | 10.80 | 19.07 | |
| 600 | 23.21 | 23.95 | 30.02 | 10.91 | 13.08 | 19.21 | 10.29 | 14.69 | 19.92 | |
| Times | Uptake Values of 99mTc-Labeled H-1-2 (ID%/mL) | |||||
|---|---|---|---|---|---|---|
| Stomach | Intestine | Heart | Liver | Kidney | Bladder | |
| 0 h | 50.725 | 0.000 | 0.002 | 0.025 | 0.026 | 0.012 |
| 2 h | 19.057 | 13.123 | 0.050 | 0.026 | 0.029 | 0.024 |
| 4 h | 18.750 | 16.456 | 0.060 | 0.018 | 0.054 | 0.008 |
| 6 h | 18.191 | 6.264 | 0.026 | 0.022 | 0.114 | 0.015 |
| 8 h | 18.074 | 3.664 | 0.048 | 0.016 | 0.112 | 0.014 |
| Times | Uptake Values of Free Technetium (ID%/mL) | |||||
|---|---|---|---|---|---|---|
| Stomach | Intestine | Heart | Liver | Kidney | Bladder | |
| 0 h | 36.527 | 0.000 | 0.028 | 0.042 | 0.100 | 0.028 |
| 2 h | 34.240 | 0.025 | 0.039 | 0.041 | 0.132 | 0.044 |
| 4 h | 24.566 | 0.057 | 0.054 | 0.084 | 0.090 | 0.157 |
| 6 h | 22.249 | 0.131 | 0.010 | 0.057 | 0.097 | 0.108 |
| 8 h | 20.229 | 0.033 | 0.008 | 0.147 | 0.064 | 0.066 |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, J.; Pang, W.; Shi, W.; Yang, B.; Kan, Y.; He, Z.; Hu, J. Structural Elucidation of a Novel Polysaccharide from Pseudostellaria heterophylla and Stimulating Glucose Uptake in Cells and Distributing in Rats by Oral. Molecules 2016, 21, 1233. https://doi.org/10.3390/molecules21091233
Chen J, Pang W, Shi W, Yang B, Kan Y, He Z, Hu J. Structural Elucidation of a Novel Polysaccharide from Pseudostellaria heterophylla and Stimulating Glucose Uptake in Cells and Distributing in Rats by Oral. Molecules. 2016; 21(9):1233. https://doi.org/10.3390/molecules21091233
Chicago/Turabian StyleChen, Jinlong, Wensheng Pang, Wentao Shi, Bin Yang, Yongjun Kan, Zhaodong He, and Juan Hu. 2016. "Structural Elucidation of a Novel Polysaccharide from Pseudostellaria heterophylla and Stimulating Glucose Uptake in Cells and Distributing in Rats by Oral" Molecules 21, no. 9: 1233. https://doi.org/10.3390/molecules21091233
APA StyleChen, J., Pang, W., Shi, W., Yang, B., Kan, Y., He, Z., & Hu, J. (2016). Structural Elucidation of a Novel Polysaccharide from Pseudostellaria heterophylla and Stimulating Glucose Uptake in Cells and Distributing in Rats by Oral. Molecules, 21(9), 1233. https://doi.org/10.3390/molecules21091233