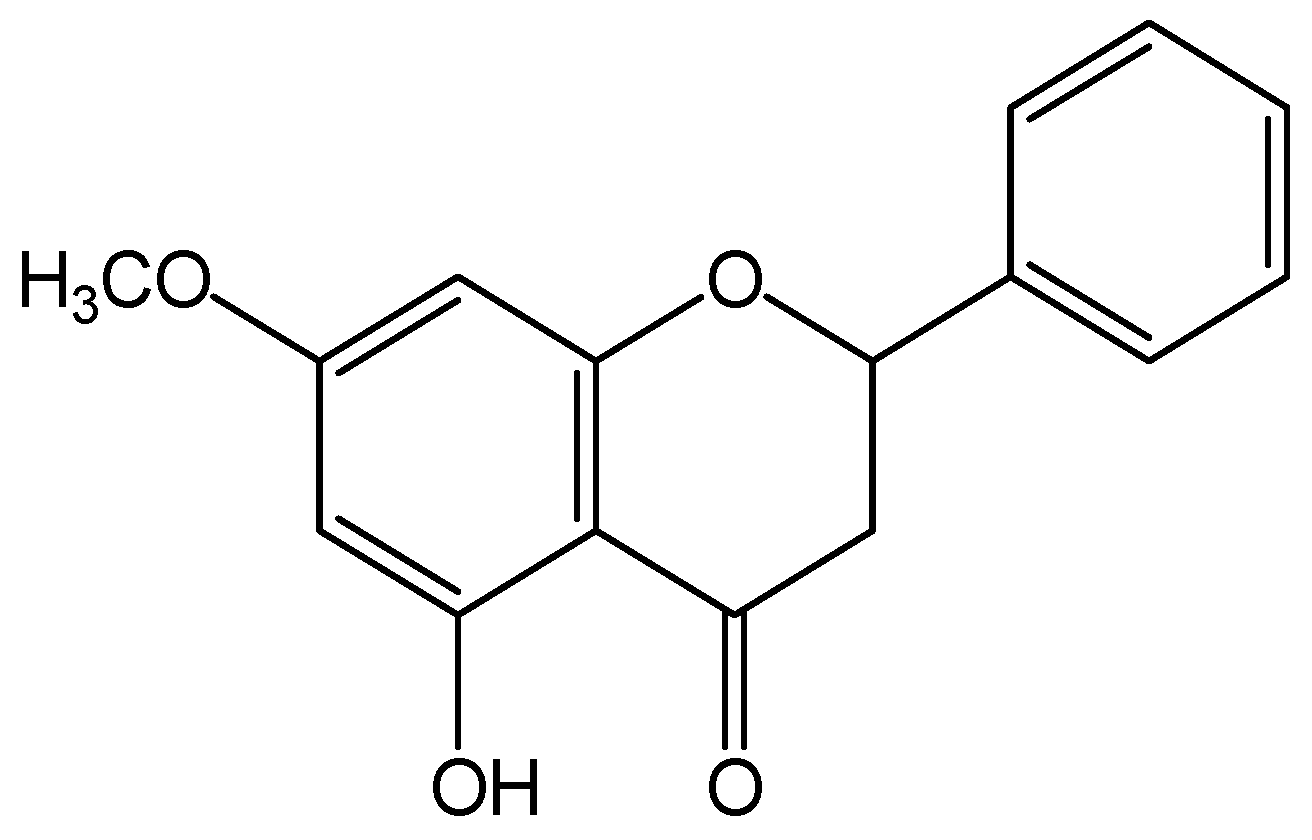
Promoting Effect of Pinostrobin on the Proliferation, Differentiation, and Mineralization of Murine Pre-osteoblastic MC3T3-E1 Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
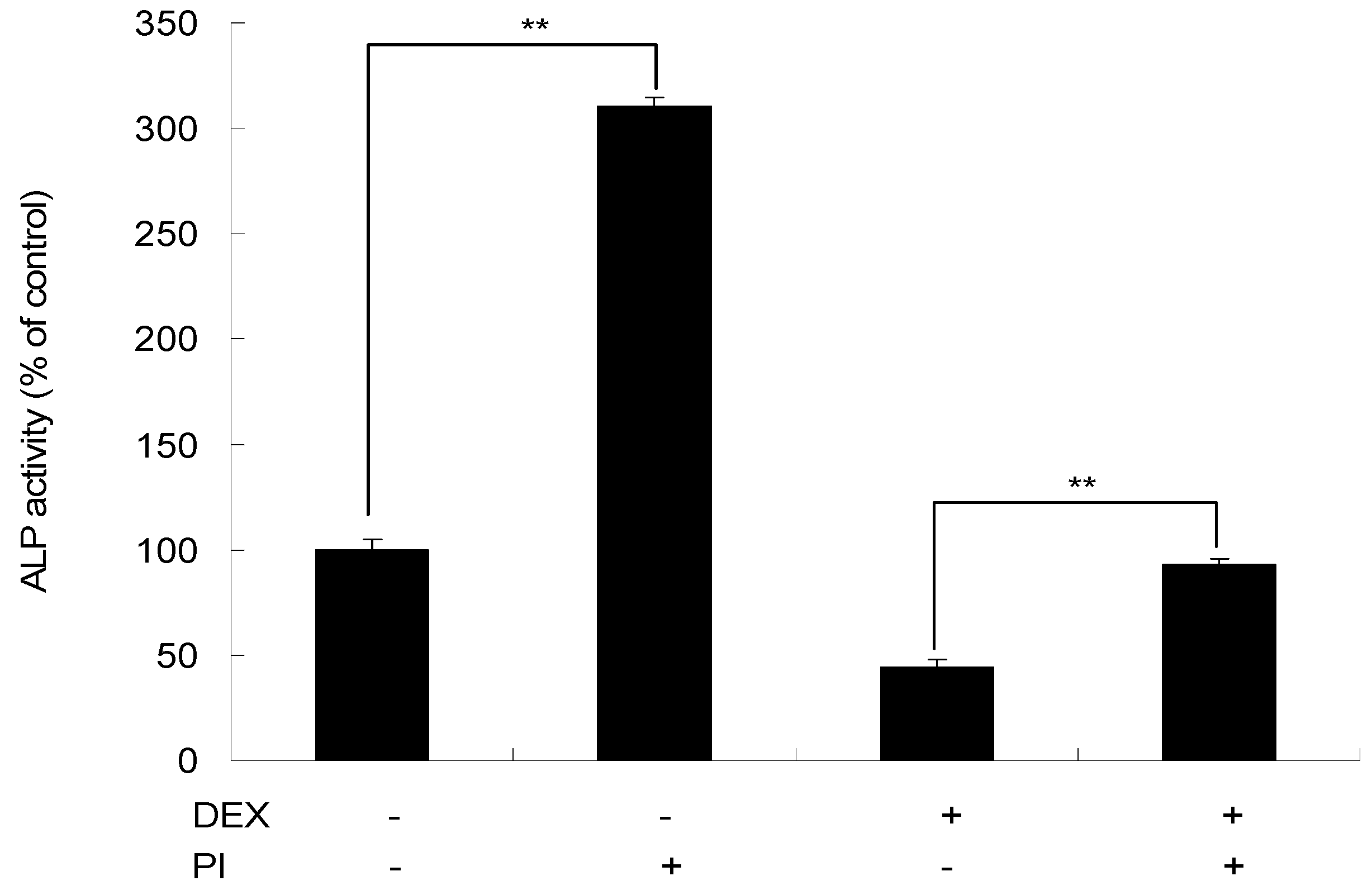{kind=link}
Abstract
:1. Introduction
2. Results
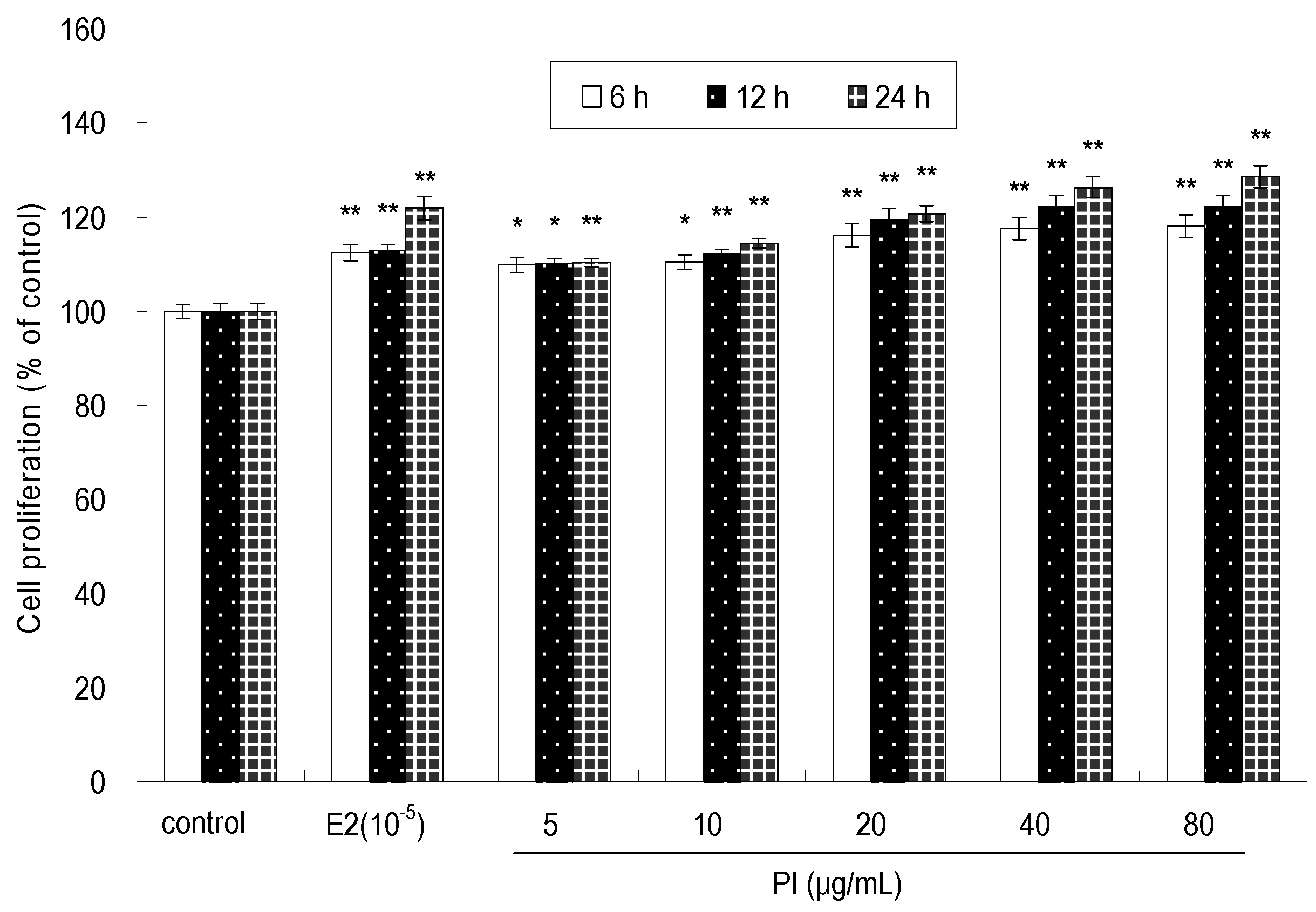
2.1. The Effect of PI on MC3T3-E1 Cell Proliferation
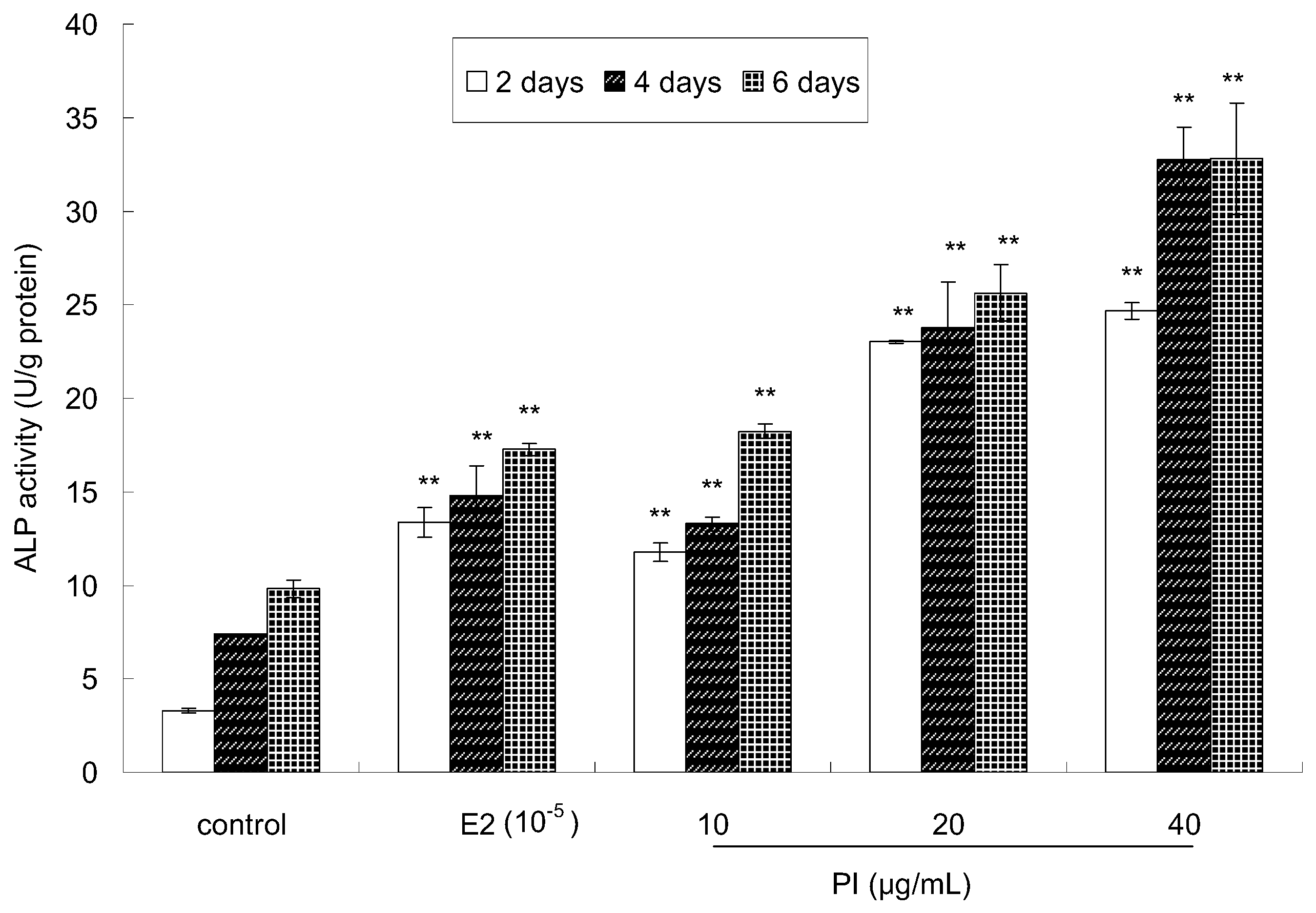
2.2. The Effect of PI on MC3T3-E1 Cell Differentiation
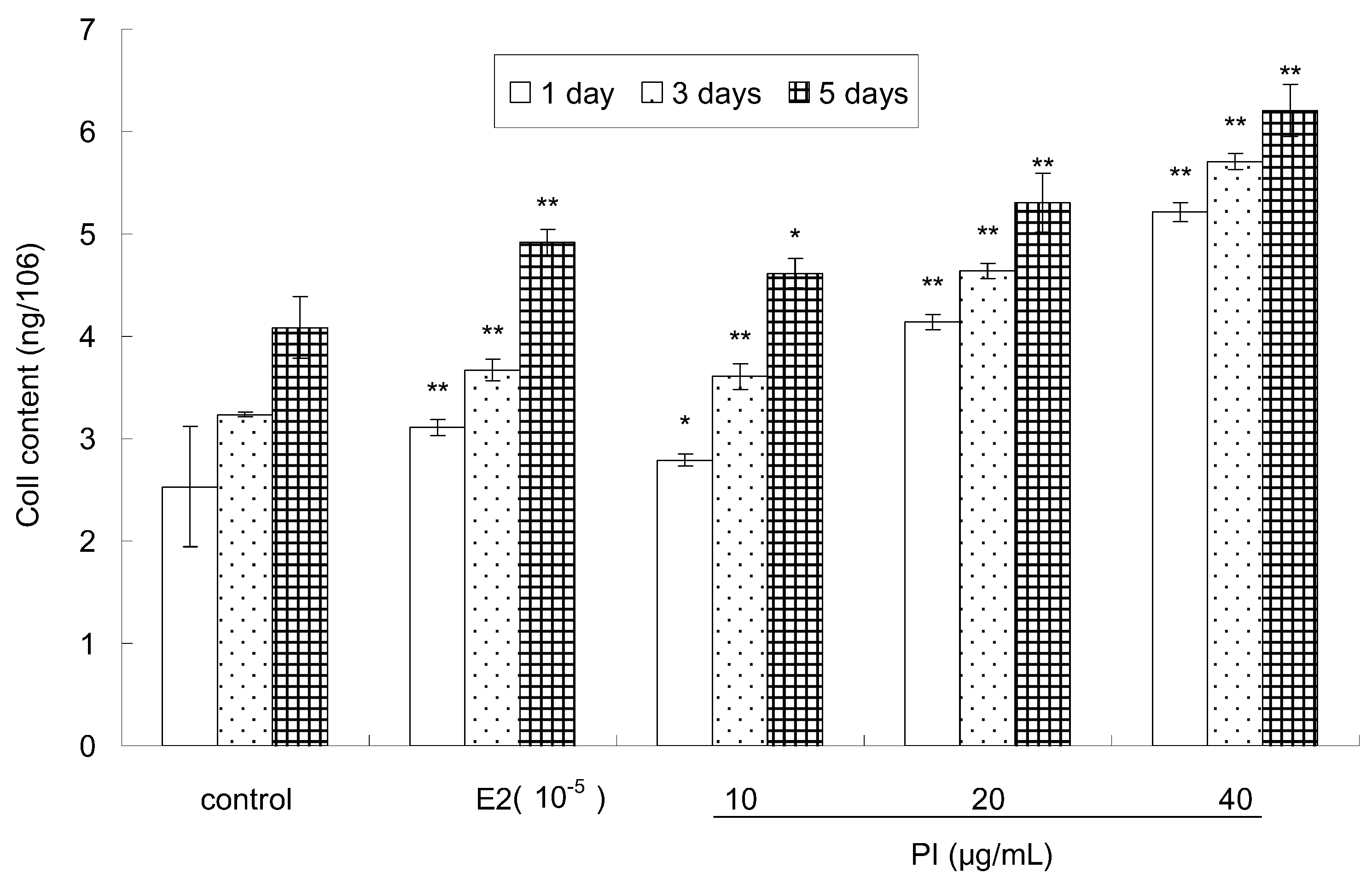
2.3. The Effect of PI on the Col I Content of MC3T3-E1 Cells
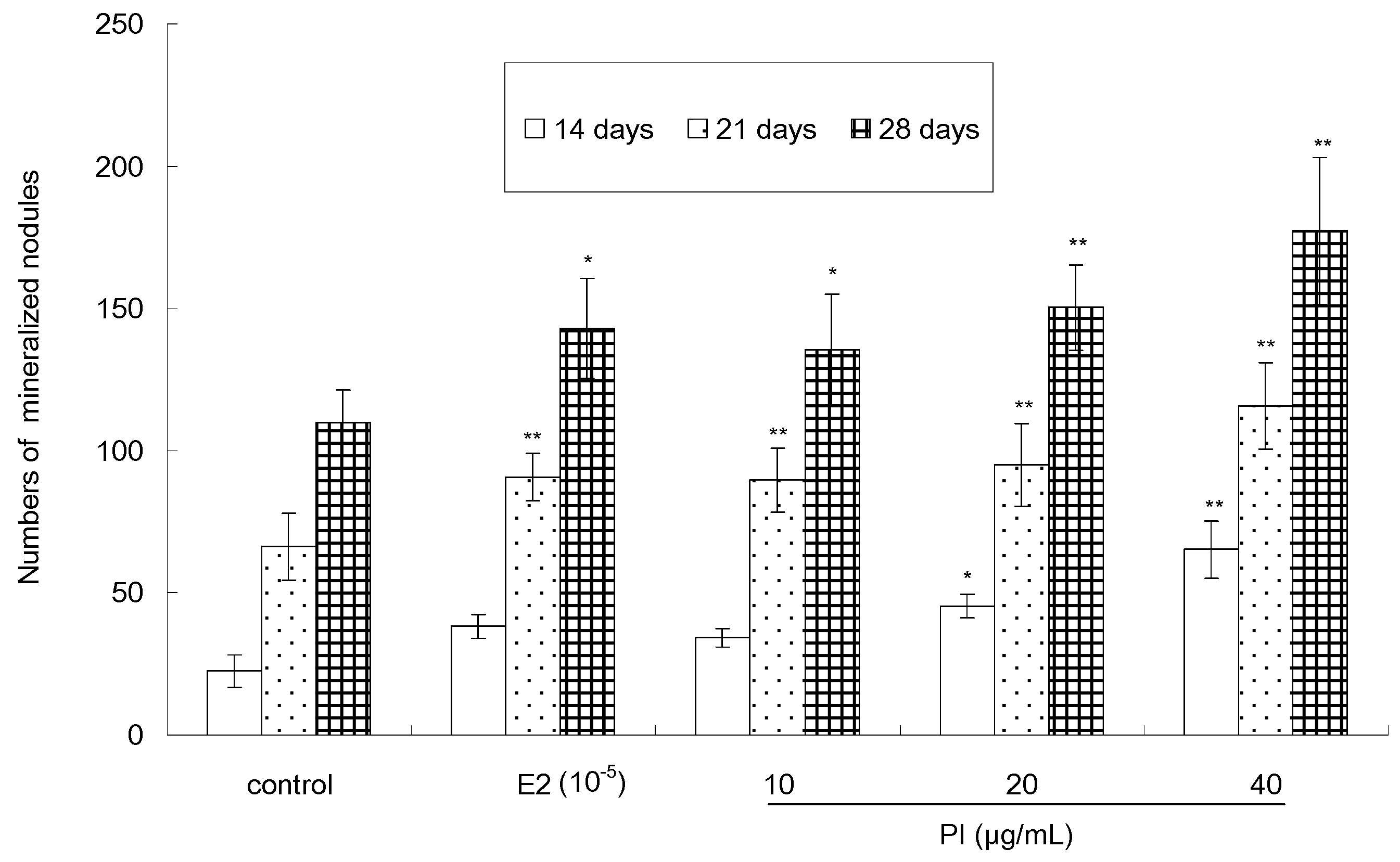
2.4. The Effect of PI on Osteoblast Mineralization of MC3T3-E1 Cells
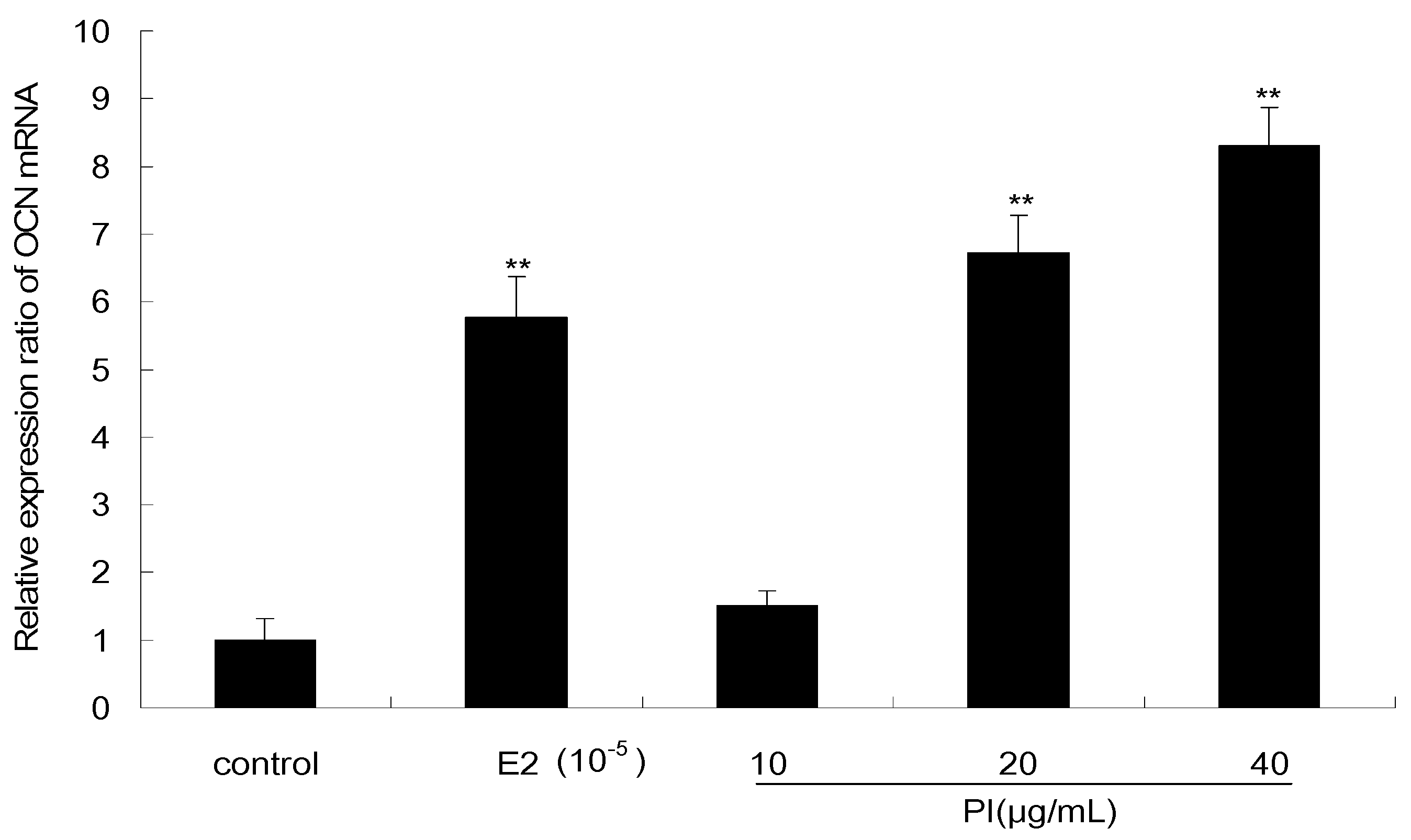
2.5. Expression Levels of OCN mRNA
2.6. The Effect of PI on DEX-Induced Inhibition of Osteogenetic Differentiation
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture
4.3. Cell Proliferation Assay
4.4. Cell Differentiation
4.5. Col I Content
4.6. Mineralized Assay
4.7. Quantitative Real-Time PCR
4.8. Effect of PI on DEX-Induced Inhibition of Osteogenetic Differentiation
4.9. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| PI | pinostrobin |
| Col I | Collagen I |
| ALP | alkaline phosphatase |
| OCN | osteocalcin |
| DEX | Dexamethasone |
References
- Kanis, J.A.; McCloskey, E.V.; Johansson, H.; Cooper, C.; Rizzoli, R.; Reginster, J.Y. European guidance for the diagnosis and management of osteoporosis in postmenopausal women. Osteoporos. Int. 2013, 24, 23–57. [Google Scholar] [CrossRef] [PubMed]
- Eastell, R.; O’Neill, T.W.; Hofbauer, L.C.; Langdahl, B.; Reid, I.R.; Gold, D.T.; Cummings, S.R. Postmenopausal osteoporosis. Nat. Rev. Dis. Prim. 2016, 2, 16069. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.L.; Li, F.B.; Ma, X.L.; Ma, J.X.; Zhao, B.; Zhang, Y.; Li, Y.J.; Lv, J.W.; Meng, X.M. The effects of combined treatment with naringin and treadmill exercise on osteoporosis in ovariectomized rats. Sci. Rep. 2015, 5, 13009. [Google Scholar] [CrossRef] [PubMed]
- Fadanelli, M.E.; Bone, H.G. Combining bisphosphonates with hormone therapy for postmenopausal osteoporosis. Treat. Endocrinol. 2004, 6, 361–369. [Google Scholar] [CrossRef]
- Maclaughlin, E.J.; Sleeper, R.B.; McNatty, D.; Raehl, C.L. Management of age-related osteoporosis and prevention of associated fractures. Ther. Clin. Risk Manag. 2006, 2, 281–295. [Google Scholar] [CrossRef] [PubMed]
- Rizzoli, R.; Reginster, J.Y.; Boonen, S.; Breart, G.; Diez-Perez, A.; Felsenberg, D.; Kaufman, J.M.; Kanis, J.A.; Cooper, C. Adverse reactions and drug-drug interactions in the management of women with postmenopausal osteoporosis. Calcif. Tissue Int. 2011, 89, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Vihtamaki, T.; Savilahti, R.; Tuimala, R. Why do postmenopausal women discontinue hormone replacement therapy? Maturitas 1999, 33, 99–105. [Google Scholar] [CrossRef]
- Kwak, E.J.; Lee, Y.S.; Choi, E.M. Effect of magnolol on the function of osteoblastic MC3T3-E1 cells. Mediat. Inflamm. 2012, 2012, 829650. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.M.; Kim, D.H.; Han, H.J.; Park, C.M.; Ganipisetti, S.R.; Valan Arasu, M.; Kim, Y.O.; Park, C.G.; Kim, B.Y.; Soung, N.K. Ginsenoside Re promotes osteoblast differentiation in mouse osteoblast precursor MC3T3-E1 Cells and a zebrafish model. Molecules 2016, 22, 42. [Google Scholar] [CrossRef] [PubMed]
- Mobasheri, A.; Shakibaei, M. Osteogenic effects of resveratrol in vitro: potential for the prevention and treatment of osteoporosis. Ann. N. Y. Acad. Sci. 2013, 1290, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Kumagai, M.; Mishima, T.; Watanabe, A.; Harada, T.; Yoshida, I.; Fujita, K.; Watai, M.; Tawata, S.; Nishikawa, K.; Morimoto, Y. 5,6-Dehydrokawain from Alpinia zerumbet promotes osteoblastic MC3T3-E1 cell differentiation. Biosci. Biotechnol. Biochem. 2016, 80, 1425–1432. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Zeng, Z.; Cai, G. Comparison of neoeriocitrin and naringin on proliferation and osteogenic differentiation in MC3T3-E1. Phytomedicine 2011, 18, 985–989. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.A.; Subhan, N.; Rahman, M.M.; Uddin, S.J.; Reza, H.M.; Sarker, S.D. Effect of citrus flavonoids, naringin and naringenin, on metabolic syndrome and their mechanisms of action. Adv. Nutr. 2014, 4, 404–417. [Google Scholar] [CrossRef] [PubMed]
- Wei, M.; Yang, Z.L.; Li, P.; Zhang, Y.B.; Sse, W.C. Anti-osteoporosis activity of naringin in the retinoic acid-induced osteoporosis model. Am. J. Chin. Med. 2007, 4, 663–667. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Jiang, Y.; Wooley, P.H.; Xu, Z.; Yang, S.Y. Naringin promotes osteoblast differentiation and effectively reverses ovariectomy-associated osteoporosis. J. Orthop. Sci. 2013, 3, 478–485. [Google Scholar]
- Wong, R.W.; Rabie, A.B. Effect of naringin on bone cells. J. Orthop. Res. 2006, 24, 2045–2050. [Google Scholar] [CrossRef] [PubMed]
- Christov, R.; Trusheva, B.; Popova, M.; Bankova, V.; Bertrand, M. Chemical composition of propolis from Canada, its antiradical activity and plant origin. Nat. Prod. Res. 2006, 20, 531–536. [Google Scholar] [CrossRef] [PubMed]
- Patel, N.K.; Bhutani, K.K. Pinostrobin and Cajanus lactone isolated from Cajanus cajan (L.) leaves inhibits TNF-alpha and IL-1 beta production: In vitro and in vivo experimentation. Phytomedicine 2014, 21, 946–953. [Google Scholar] [CrossRef] [PubMed]
- Patel, N.K.; Jaiswal, G.; Bhutani, K.K. A review on biological sources, chemistry and pharmacological activities of pinostrobin. Nat. Prod. Res. 2016, 30, 2017–2027. [Google Scholar] [CrossRef] [PubMed]
- Wu, N.; Kong, Y.; Zu, Y.; Fu, Y.; Liu, Z.; Meng, R.; Liu, X.; Efferth, T. Activity investigation of pinostrobin towards herpes simplex virus-1 as determined by atomic force microscopy. Phytomedicine 2011, 18, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Xian, Y.-F.; Ip, S.-P.; Lin, Z.-X.; Mao, Q.-Q.; Su, Z.-R.; Lai, X.-P. Protective effects of pinostrobin on beta-amyloid-induced neurotoxicity in PC12 cells. Cell. Mol. Neurobiol. 2012, 32, 1223–1230. [Google Scholar] [CrossRef] [PubMed]
- Le Bail, J.C.; Aubourg, L.; Habrioux, G. Effects of pinostrobin on estrogen metabolism and estrogen receptor transactivation. Cancer Lett. 2000, 156, 37–44. [Google Scholar] [CrossRef]
- Ashidi, J.S.; Houghton, P.J.; Hylands, P.J.; Efferth, T. Ethnobotanical survey and cytotoxicity testing of plants of South-western Nigeria used to treat cancer, with isolation of cytotoxic constituents from Cajanus cajan Millsp. leaves. J. Ethnopharmacol. 2010, 128, 501–512. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liu, C.; Sun, J.; Liu, D.; Wang, P. Effects of water extract of Cajanus cajan leaves on the osteogenic and adipogenic differentiation of mouse primary bone marrow stromal cells and the adipocytic trans-differentiation of mouse primary osteoblasts. Pharm. Biol. 2010, 48, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Wu, N.; Fu, K.; Fu, Y.-J.; Zu, Y.-G.; Chang, F.-R.; Chen, Y.-H.; Liu, X.-L.; Kong, Y.; Liu, W.; Gu, C.-B. Antioxidant activities of extracts and main components of pigeonpea [Cajanus cajan (L.) Millsp.] leaves. Molecules 2009, 14, 1032–1043. [Google Scholar] [CrossRef] [PubMed]
- Quarles, L.D.; Yohay, D.A.; Lever, L.W.; Caton, R.; Wenstrup, R.J. Distinct proliferative and differentiated stages of murine MC3T3-E1 cells in culture: an in vitro model of osteoblast development. J. Bone Miner. Res. 1992, 7, 683–692. [Google Scholar] [CrossRef] [PubMed]
- Willett, T.L.; Pasquale, J.; Grynpas, M.D. Collagen modifications in postmenopausal osteoporosis: advanced glycation endproducts may affect bone volume, structure and quality. Curr. Osteoporos. Rep. 2014, 12, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, M.; Weitzmann, M.N. The estrogen 17β-estradiol and phytoestrogen genistein mediate differential effects on osteoblas- tic NF-κB activity. Int. J. Mol. Med. 2009, 23, 297–301. [Google Scholar] [PubMed]
- Sugimoto, E.; Yamaguchi, M. Stimulatory effect of daidzein in osteoblastic MC3T3-E1 cells. Biochem. Pharmacol. 2000, 59, 471–475. [Google Scholar] [CrossRef]
- Yamaguchi, M. Isoflavone and bone metabolism: its cellular mechanism and preventive role in bone loss. J. Health Sci. 2002, 3, 209–222. [Google Scholar] [CrossRef]
- Pang, W.Y.; Wang, X.L.; Mok, S.K.; Lai, W.P.; Chow, H.K.; Leung, P.C.; Yao, X.S.; Wong, M.S. Naringin improves bone properties in ovariectomized mice and exerts oestrogen-like activities in rat osteoblast-like (UMR-106) cells. Br. J. Pharmacol. 2010, 159, 1693–1703. [Google Scholar] [CrossRef] [PubMed]
- Kahai, S.; Lee, S.C.; Seth, A.; Yang, B.B. Nephronectin promotes osteoblast differentiation via the epidermal growth factor-like repeats. FEBS Lett. 2010, 584, 233–238. [Google Scholar] [CrossRef] [PubMed]
- Owen, T.A.; Holthuis, J.; Markose, E.; van Wijnen, A.J.; Wolfe, S.A.; Grimes, S.R.; Lian, J.B.; Stein, G.S. Modifications of protein-DNA interactions in the proximal promoter of a cell-growth-regulated histone gene during onset and progression of osteoblast differentiation. Proc. Natl. Acad. Sci. USA 1990, 87, 5129–5133. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.P.; Delmas, P.D.; Malaval, L.; Edouard, C.; Chapuy, M.C.; Meunier, P.J. Serum bone Gla-protein: A specific marker for bone formation in postmenopausal osteoporosis. Lancet 1984, 1, 1091–1093. [Google Scholar] [CrossRef]
- Stein, G.S.; Lian, J.B.; Owen, T.A. Relationship of cell growth to the regulation of tissue-specific gene expression during osteoblast differentiation. FASEB J. 1990, 4, 3111–3123. [Google Scholar] [PubMed]
- Hu, Y.; Li, J.C.; Zhu, X.J.; Li, Y.H.; Zhang, S.; Chen, X.J.; Gao, Y.H.; Li, F.Y. 17β-estradiol-loaded peglyated upconversion nanoparticles as a bone-targeted drug nanocarrier. ACS Appl. Mater. Interfaces 2015, 29, 15803–15811. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.B.; Li, Y.H.; Kong, X.H.; Zhang, R.; Sun, Y.; Li, Q.; Li, C.; Liu, L.; Wang, J.; Mei, Q.B. The beneficial effect of Radix Dipsaci total saponins on bone metabolism in vitro and in vivo and the possible mechanisms of action. Osteoporos. Int. 2012, 23, 2649–2660. [Google Scholar] [CrossRef] [PubMed]
- Satija, N.K.; Gurudutta, G.U.; Sharma, S.; Afrin, F.; Gupta, P.; Verma, Y.K.; Singh, V.K.; Tripathi, R.P. Mesenchymal stem cells: Molecular targets for tissue engineering. Stem Cells Dev. 2007, 16, 7–23. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Lee, H.; Kang, K.S.; Chun, K.H.; Hwang, G.S. Protective effect of Korean Red Ginseng against glucocorticoid-induced osteoporosis in vitro and in vivo. J. Ginseng Res. 2015, 39, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Canalis, E. Mechanisms of glucocorticoid action in bone. Curr. Osteoporos. Rep. 2005, 3, 98–102. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Jiang, Z.L.; Chen, L.H.; Sun, Y.; Chen, S.Z.; Zhou, P.; Xia, A.X.; Jin, H.; Zhu, Y.W.; Chen, D.Y. Glucocorticoid-induced leucine zipper may play an important role in icariin by suppressing osteogenesis inhibition induced by glucocorticoids in osteoblasts. Biomed. Pharmacother. 2017, 90, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Charoensin, S.; Punvittayagul, C.; Pompimon, C.; Mevatee, U.; Wongpoomchai, R. Toxicological and clastogenic evaluation of pinocembrin and pinostrobin isolated from Boesenbergia pandurata in Wistar rats. Thai J. Toxicol. 2010, 1, 29–40. [Google Scholar]
- Sayre, C.L.; Davies, N.M. Quantification of three chiral flavonoids with reported bioactivity in selected licensed canadian natural health products and US marketed dietary supplements. J. Pharm. Pharm. Sci. 2013, 16, 272–278. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Not available. |








© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gu, C.; Fu, L.; Yuan, X.; Liu, Z. Promoting Effect of Pinostrobin on the Proliferation, Differentiation, and Mineralization of Murine Pre-osteoblastic MC3T3-E1 Cells. Molecules 2017, 22, 1735. https://doi.org/10.3390/molecules22101735
Gu C, Fu L, Yuan X, Liu Z. Promoting Effect of Pinostrobin on the Proliferation, Differentiation, and Mineralization of Murine Pre-osteoblastic MC3T3-E1 Cells. Molecules. 2017; 22(10):1735. https://doi.org/10.3390/molecules22101735
Chicago/Turabian StyleGu, Chengbo, Linan Fu, Xiaohan Yuan, and Zhiguo Liu. 2017. "Promoting Effect of Pinostrobin on the Proliferation, Differentiation, and Mineralization of Murine Pre-osteoblastic MC3T3-E1 Cells" Molecules 22, no. 10: 1735. https://doi.org/10.3390/molecules22101735
APA StyleGu, C., Fu, L., Yuan, X., & Liu, Z. (2017). Promoting Effect of Pinostrobin on the Proliferation, Differentiation, and Mineralization of Murine Pre-osteoblastic MC3T3-E1 Cells. Molecules, 22(10), 1735. https://doi.org/10.3390/molecules22101735