
A Novel Polysaccharide Conjugate from Bullacta exarata Induces G1-Phase Arrest and Apoptosis in Human Hepatocellular Carcinoma HepG2 Cells

Abstract
:
1. Introduction
2. Results and Discussion
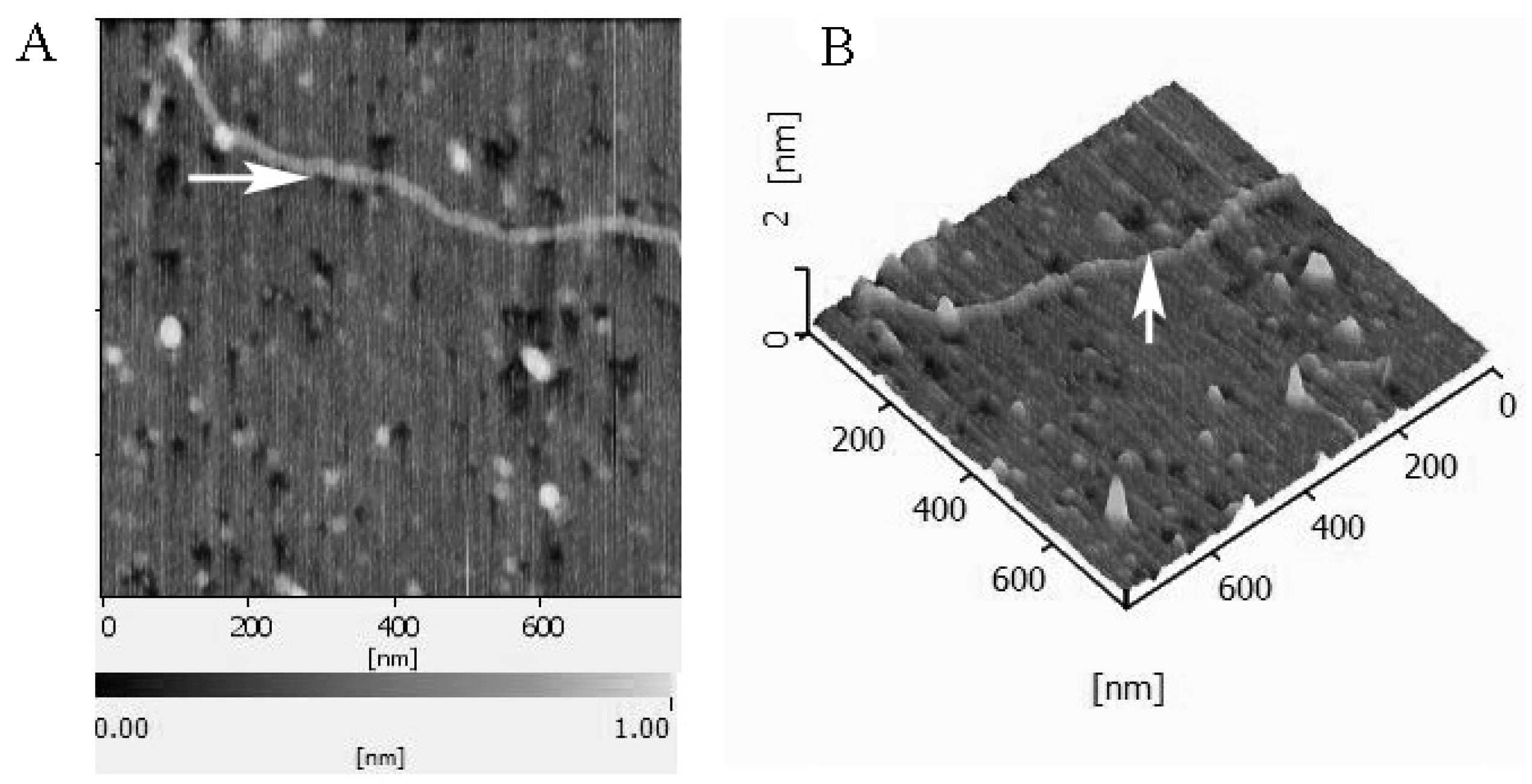
2.1. Composition Analyses and Image of BEPS-IA
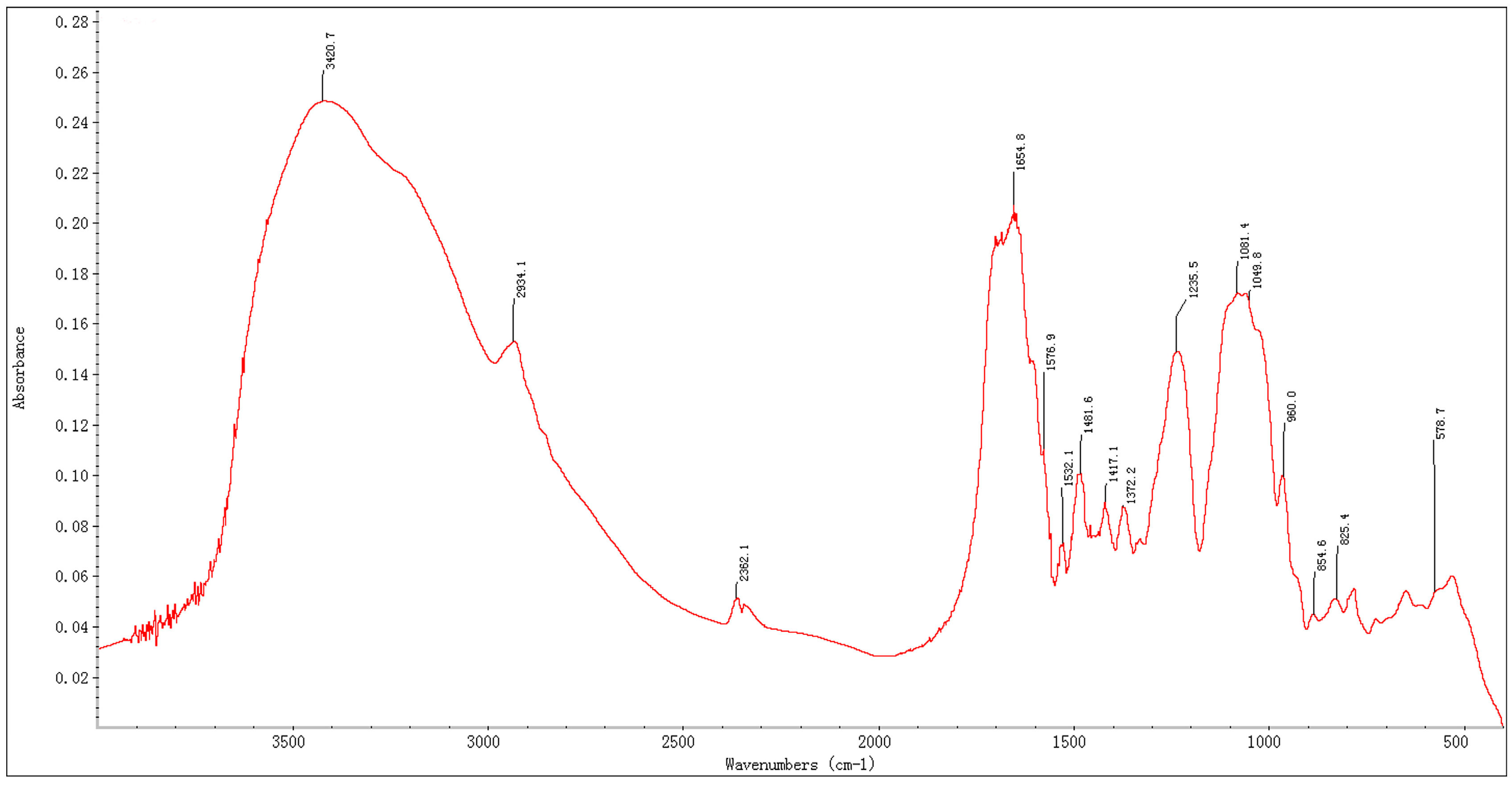
2.2. UV and IR Spectrum of BEPS-IA
2.3. Structure Data from NMR
2.4. Structure Data from Methylation Analysis
2.5. Anti-Proliferation and Cytotoxicity Assay
2.6. Effect of BEPS-IA on Cell Cycle Distribution of HepG2 Cells
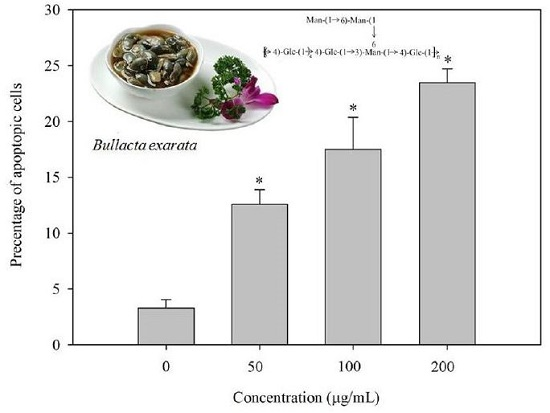
2.7. BEPS-IA Induced Apoptosis of HepG2 Cells In Vitro
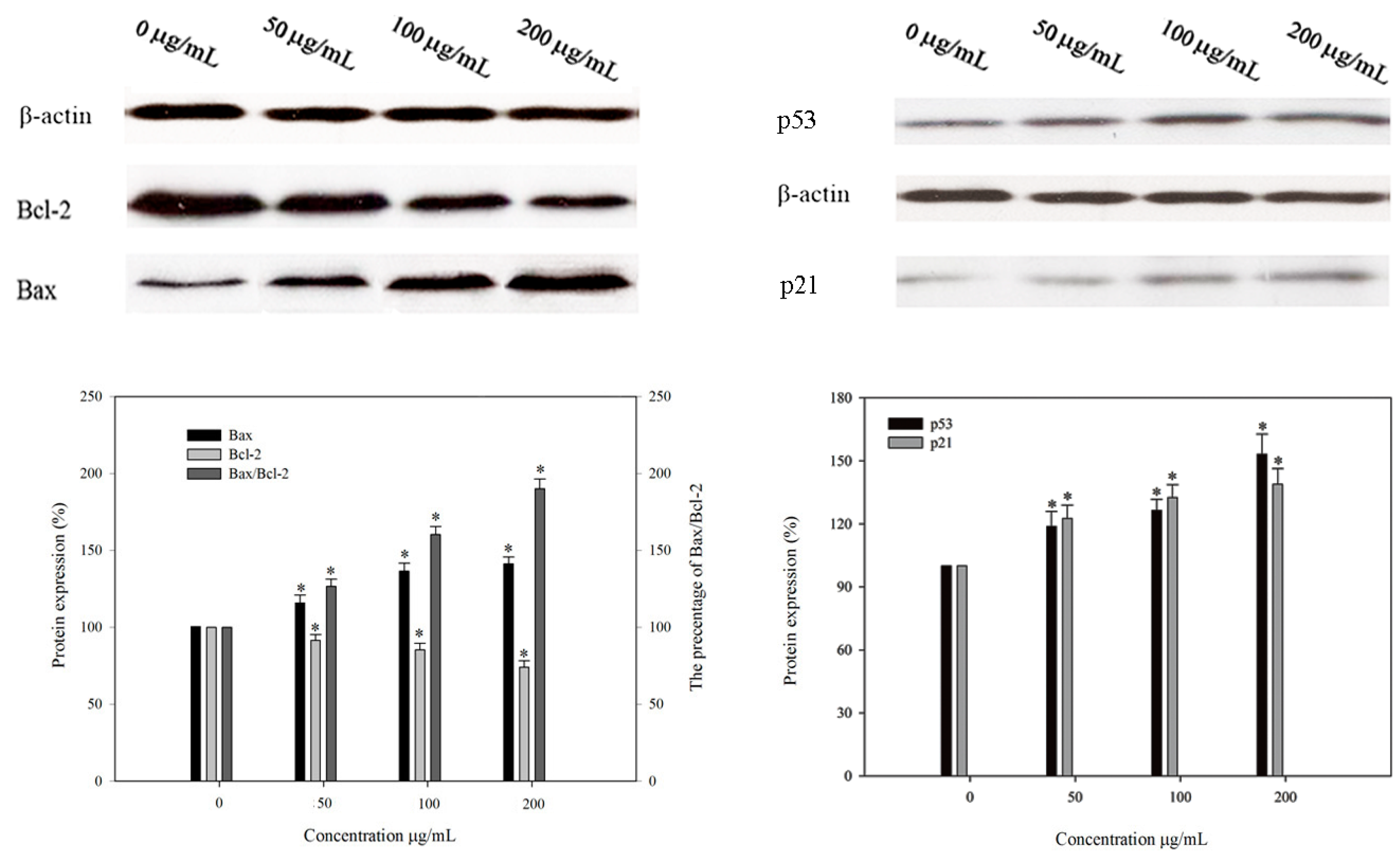
2.8. Effect of BEPS-IA on Protein Expression of p53, p21, Bcl-2 and Bax in HepG2 Cells
3. Materials and Methods
3.1. Materials and Reagents
3.2. Polysaccharide-Protein Complex BEPS-IA Extraction
3.3. Analytical Procedures
3.4. Methylation Analyses
3.5. FI-IR and NMR Analyses
3.6. Cell Culture
3.7. Cytotoxicity Measurements
3.8. Measurement of Cell Proliferation
3.9. TUNEL Assay
3.10. Cell Cycle Analysis
3.11. Western Blotting Analysis
3.12. Statistical Analyses
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| BEPS-IA | purified Bullacta exarata polysaccharides of molecular weight 127 kDa |
| AFM | Atomic Force Microscopy |
| NMR | Nuclear Magnetic Resonance |
| PMP | 1-phenyl-3-methyl-5-pyrazolone |
| CK | Control culture |
| IC50 | inhibitory concentration representing the concentration at which cell viability was reduced by 50% |
| HepG2 | human hepatocellular carcinoma cell |
| MEM | Medium Alpha Medium |
| Gal | galactose |
| Man | mannose |
| Glc | glucose |
| GlcUA | glucuronic acid |
| Arb | arabinose |
| Rha | rhamnose |
| Xyl | xylose |
| GalN | galactosamine |
| GlcN | glucosamine |
| Fuc | fucose |
References
- Zhang, D.; Wang, C.; Wu, H.; Xie, J.; Du, L.; Xia, Z.; Cai, J.; Huang, Z.; Wei, D. Three sulphated polysaccharides isolated from the mucilage of mud snail, Bullacta exarata philippi: Characterization and antitumour activity. Food Chem. 2013, 138, 306–314. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Liao, N.; Ye, X.; Hu, Y.; Wu, D.; Guo, X.; Zhong, J.; Wu, J.; Chen, S. Isolation and structural characterization of a novel antioxidant mannoglucan from a marine bubble snail, Bullacta exarata (Philippi). Mar. Drugs 2013, 11, 4464–4477. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Wu, H.; Xia, Z.; Wang, C.; Cai, J.; Huang, Z.; Du, L.; Sun, P.; Xie, J. Partial characterization, antioxidant and antitumor activities of three sulfated polysaccharides purified from Bullacta exarata. J. Funct. Foods 2012, 4, 784–792. [Google Scholar] [CrossRef]
- Yuan, C.; Cui, Q.; Xu, J.; Li, G. Biological activities and spectral analysis of clycosaminoglycan from Bullacta exarata. Guangdong Agric. Sci. 2013, 3, 056. [Google Scholar]
- Ma, J.; Huang, F.; Lin, H.; Wang, X. Isolation and purification of a peptide from Bullacta exarata and its impaction of apoptosis on prostate cancer cell. Mar. Drugs 2013, 11, 266–273. [Google Scholar] [CrossRef] [PubMed]
- Yuan, C.; Xu, J.; Cui, Q. Purification and analysis of chemical compositions on glycosaminoglycan from Bullacta exarata. In Proceedings of the 3rd International Conference of Bioinformatics, Natural Products and Traditional Medicine, Xi’an, China, 14–16 October 2011; pp. 23–27.
- Torre, L.A.; Bray, F.; Siegel, R.L.; Ferlay, J.; Lortet-Tieulent, J.; Jemal, A. Global cancer statistics, 2012. CA-Cancer J. Clin. 2015, 65, 87–108. [Google Scholar] [CrossRef] [PubMed]
- Darcy, D.G.; Malek, M.M.; Kobos, R.; Klimstra, D.S.; DeMatteo, R.; la Quaglia, M.P. Prognostic factors in fibrolamellar hepatocellular carcinoma in young people. J. Pediatr. Surg. 2015, 50, 153–156. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Zhu, H.; Li, X.; Liu, Z.; Zheng, W.; Chen, T.; Yu, B.; Wong, K.H. Induction of apoptosis and cell cycle arrest in A549 human lung adenocarcinoma cells by surface-capping selenium nanoparticles: An effect enhanced by polysaccharide-protein complexes from Polyporus rhinocerus. J. Agric. Food Chem. 2013, 61, 9859–9866. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Li, L.; Fang, J.C.; Wong, J.H.; Ng, T.B.; Jiang, Y.; Wang, C.R.; Zhang, N.Y.; Wen, T.Y.; Lv, P.Y.; et al. Isolation and identification of a novel polysaccharide-peptide complex with antioxidant, anti-proliferative and hypoglycaemic activities from the abalone mushroom. Biosci. Rep. 2012, 32, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Patel, Y.; Naraian, R.; Singh, V.K. Medicinal properties of Pleurotus species (oyster mushroom): A review. World J. Fungal Plant Biol. 2012, 3, 1–12. [Google Scholar]
- Yoshida, J.; Takamura, S.; Ishibashi, T.; Nishio, M. Antiproliferative and apoptosis-inducing effects of an antitumor glycoprotein from Streptococcus pyogenes. Anticancer Res. 1998, 19, 1865–1871. [Google Scholar]
- Tsai, P.F.; Ma, C.Y.; Wu, J.S.B. A novel glycoprotein from mushroom Hypsizygus marmoreus (Peck) Bigelow with growth inhibitory effect against human leukaemic U937 cells. Food Chem. 2013, 141, 1252–1258. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhou, W.; Zhang, Y. Immunoregulation and Lycium Barbarum. In Lycium Barbarum and Human Health; Springer: Dordrecht, The Netherlands, 2015; pp. 27–44. [Google Scholar]
- Batista, J.A.; Dias, E.G.; Brito, T.V.; Prudêncio, R.S.; Silva, R.O.; Ribeiro, R.A.; Souzab, M.H.; Paulad, R.C.; Feitosad, J.P.; Melo, M.R.; et al. Polysaccharide isolated from Agardhiella ramosissima: Chemical structure and anti-inflammation activity. Carbohyd. Polym. 2014, 99, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Wang, Y.; Li, X.; Yu, P. Purification and structural characterization of Chinese yam polysaccharide and its activities. Carbohyd. Polym. 2015, 117, 1021–1027. [Google Scholar] [CrossRef] [PubMed]
- Dore, C.M.P.G.; Alves, M.G.D.C.F.; Santos, M.D.G.L.; de Souza, L.A.R.; Baseia, I.G.; Leite, E.L. Antioxidant and anti-inflammatory properties of an extract rich in polysaccharides of the mushroom Polyporus dermoporus. Antioxidants 2014, 3, 730–744. [Google Scholar] [CrossRef] [PubMed]
- Karnchanatat, A.; Puthong, S.; Sihanonth, P.; Piapukiew, J.; Sangvanich, P. Antioxidation and antiproliferation properties of polysaccharide-protein complex extracted from Phaeogyroporus portentosus (Berk. Broome) McNabb. Afr. J. Microbiol. Res. 2013, 7, 1668–1680. [Google Scholar]
- Njei, B.; Rotman, Y.; Ditah, I.; Lim, J.K. Emerging trends in hepatocellular carcinoma incidence and mortality. Hepatology 2015, 61, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Chinembiri, T.N.; Du Plessis, L.H.; Gerber, M.; Hamman, J.H.; Du Plessis, J. Review of natural compounds for potential skin cancer treatment. Molecules 2014, 19, 11679–11721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guadagno, J.; Swan, P.; Shaikh, R.; Cregan, S.P. Microglia-derived IL-1β triggers p53-mediated cell cycle arrest and apoptosis in neural precursor cells. Cell Death Dis. 2015, 6, e1779. [Google Scholar] [CrossRef] [PubMed]
- Karimian, A.; Ahmadi, Y.; Yousefi, B. Multiple functions of p21 in cell cycle, apoptosis and transcriptional regulation after DNA damage. DNA Repair 2016, 42, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Al-Fatlawi, A.A.; Al-Fatlawi, A.A.; Irshad, M.; Zafaryab, M.; Rizvi, M.M.; Ahmad, A. Rice bran phytic acid induced apoptosis through regulation of Bcl-2/Bax and p53 genes in HepG2 human hepatocellular carcinoma cells. Asian Pac. J. Cancer Prev. 2014, 15, 3731–3736. [Google Scholar] [CrossRef] [PubMed]
- Czabotar, P.E.; Lessene, G.; Strasser, A.; Adams, J.M. Control of apoptosis by the BCL-2 protein family: Implications for physiology and therapy. Nat. Rev. Mol. Cell Biol. 2014, 15, 49–63. [Google Scholar] [CrossRef] [PubMed]
- Raemy, E.; Martinou, J.C. Involvement of cardiolipin in tBID-induced activation of BAX during apoptosis. Chem. Phys. Lipids 2014, 179, 70–74. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.L.; Zhu, W.; Wang, M.; Hu, M.M.; Chen, W.L.; Xu, X.J.; Lu, C.J. Polysaccharides from Smilax glabra inhibit the pro-inflammatory mediators via ERK1/2 and JNK pathways in LPS-induced RAW264. 7 cells. Carbohyd. Polym. 2015, 122, 428–436. [Google Scholar]
- Zong, A.; Cao, H.; Wang, F. Anticancer polysaccharides from natural resources: A review of recent research. Carbohyd. Polym. 2012, 90, 1395–1410. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.Y.; Bai, X.Y.; Wang, C.H. Traditional Chinese medicine: A treasured natural resource of anticancer drug research and development. Am. J. Chinese Med. 2014, 42, 543–559. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, S.S. Phenol-sulfuric acid method for total carbohydrates. In Food Analysis Laboratory Manual; Springer: New York, NY, USA, 2010; pp. 47–53. [Google Scholar]
- Ermakova, S.; Sokolova, R.; Kim, S.M.; Um, B.H.; Isakov, V.; Zvyagintseva, T. Fucoidans from brown seaweeds Sargassum hornery, Eclonia cava, Costaria costata: Structural characteristics and anticancer activity. Appl. Biochem. Biotechnol. 2011, 164, 841–850. [Google Scholar] [CrossRef] [PubMed]
- Zeng, F.; Ou, J.; Huang, Y.; Li, Q.; Xu, G.; Liu, Z.; Yang, S. Determination of 21 free amino acids in fruit juices by HPLC using a modification of the 6-aminoquinolyl-N-hydroxysuccinimidyl carbamate (AQC) method. Food Anal. Method 2015, 8, 428–437. [Google Scholar] [CrossRef]
- Zhu, Y.; Yang, Y.; Zhou, Z.; Li, G.; Jiang, M.; Zhang, C.; Chen, S. Direct determination of free tryptophan contents in soy sauces and its application as an index of soy sauce adulteration. Food Chem. 2010, 118, 159–162. [Google Scholar] [CrossRef]
- Zhang, T.; Zheng, Y.; Cosgrove, D.J. Spatial organization of cellulose microfibrils and matrix polysaccharides in primary plant cell walls as imaged by multichannel atomic force microscopy. Plant J. 2016, 85, 179–192. [Google Scholar] [CrossRef] [PubMed]
- Souza, R.O.; Assreuy, A.M.; Madeira, J.C.; Chagas, F.D.; Parreiras, L.A.; Santos, G.R.; Mourãoc, P.A.; Pereira, M.G. Purified polysaccharides of Geoffroea spinosa barks have anticoagulant and antithrombotic activities devoid of hemorrhagic risks. Carbohyd. Polym. 2015, 124, 208–215. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; Zhang, Y.; Mu, H.; Lv, Z.; Yang, Y.; Zhang, J. Structural elucidation and antioxidant activity of a novel polysaccharide (TAPB1) from Tremella aurantialba. Food Hydrocolloid 2015, 43, 459–464. [Google Scholar] [CrossRef]
- Nwokocha, L.M.; Williams, P.A. Rheological properties of a polysaccharide isolated from Adansonia digitata leaves. Food Hydrocoll. 2016, 58, 29–34. [Google Scholar] [CrossRef]
- Chen, J.; Tian, J.; Ge, H.; Liu, R.; Xiao, J. Effects of tetramethylpyrazine from Chinese black vinegar on antioxidant and hypolipidemia activities in HepG2 cells. Food Chem. Toxicol. 2016, in press. [Google Scholar] [CrossRef] [PubMed]
- Felice, D.L.; Sun, J.; Liu, R.H. A modified methylene blue assay for accurate cell counting. J. Funct. Foods 2009, 1, 109–118. [Google Scholar] [CrossRef]
- Zhu, Y.; Li, T.; Fu, X.; Abbasi, A.M.; Zheng, B.; Liu, R.H. Phenolics content, antioxidant and antiproliferative activities of dehulled highland barley (Hordeum vulgare L.). J. Funct. Foods 2015, 19, 439–450. [Google Scholar] [CrossRef]
- Jiang, X.; Li, T.; Liu, R.H. 2α-Hydroxyursolic acid inhibited cell proliferation and induced apoptosis in MDA-MB-231 human breast cancer cells through the p38/MAPK signal transduction pathway. J. Agric. Food Chem. 2016, 64, 1806–1816. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Not available.









{kind=link}
{kind=link}
{kind=link}
{kind=link}
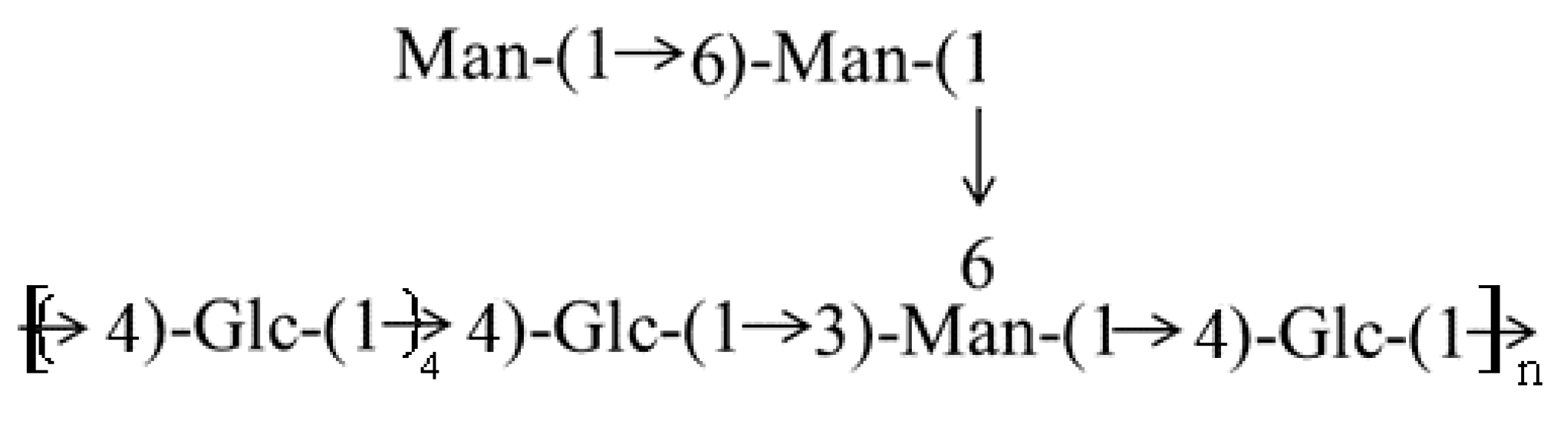{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amino Acid | Concentration (mg/g) a |
|---|---|
| Asparagine | 16.73 ± 1.37 |
| Serine | 5.33 ± 2.11 |
| Glutamic acid | 12.27 ± 3.79 |
| Glycine | 22.14 ± 0.68 |
| Arginine | 4.12 ± 1.61 |
| Threonine b | 1.04 ± 2.14 |
| Alanine | 9.26 ± 2.12 |
| Proline | 5.12 ± 2.01 |
| Cysteine | 4.28 ± 0.17 |
| Valine b | 8.95 ± 1.42 |
| Methionine b | 1.21 ± 2.04 |
| Lysine b | 2.41 ± 0.26 |
| Leucine b | 6.32 ± 0.24 |
| Total amino acids | 99.18 ± 6.43 |
| Proportion of the essential amino acids (%) | 20.09 ± 5.62 |
| Methylated Sugar | Molar Ratio | Mass Fragment (m/z) b | Type of Linkage |
|---|---|---|---|
| 2,3,6-Tri-O-Me-Glc a | 6.46 | 45, 87, 101, 113, 117, 161, 233 | →4)-Glcp-(1→ |
| 2,4,-Di-O-Me-Man | 1.13 | 71, 87, 99, 101, 189 | →3,6)-Manp-(1→ |
| 2,3,4,-Tri-O-Me-Man | 0.87 | 45, 71, 87, 101, 129, 161, 189 | →6)-Manp-(1→ |
| 2,3,4,6-Tetra-O-Me-Man | 0.91 | 28, 43, 71, 87, 101, 117, 129, 145, 161,205 | Manp-(1→ |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liao, N.; Sun, L.; Chen, J.; Zhong, J.; Zhang, Y.; Zhang, R. A Novel Polysaccharide Conjugate from Bullacta exarata Induces G1-Phase Arrest and Apoptosis in Human Hepatocellular Carcinoma HepG2 Cells. Molecules 2017, 22, 384. https://doi.org/10.3390/molecules22030384
Liao N, Sun L, Chen J, Zhong J, Zhang Y, Zhang R. A Novel Polysaccharide Conjugate from Bullacta exarata Induces G1-Phase Arrest and Apoptosis in Human Hepatocellular Carcinoma HepG2 Cells. Molecules. 2017; 22(3):384. https://doi.org/10.3390/molecules22030384
Chicago/Turabian StyleLiao, Ningbo, Liang Sun, Jiang Chen, Jianjun Zhong, Yanjun Zhang, and Ronghua Zhang. 2017. "A Novel Polysaccharide Conjugate from Bullacta exarata Induces G1-Phase Arrest and Apoptosis in Human Hepatocellular Carcinoma HepG2 Cells" Molecules 22, no. 3: 384. https://doi.org/10.3390/molecules22030384
APA StyleLiao, N., Sun, L., Chen, J., Zhong, J., Zhang, Y., & Zhang, R. (2017). A Novel Polysaccharide Conjugate from Bullacta exarata Induces G1-Phase Arrest and Apoptosis in Human Hepatocellular Carcinoma HepG2 Cells. Molecules, 22(3), 384. https://doi.org/10.3390/molecules22030384