Laxative Effects of Total Diterpenoids Extracted from the Roots of Euphorbia pekinensis Are Attributable to Alterations of Aquaporins in the Colon

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
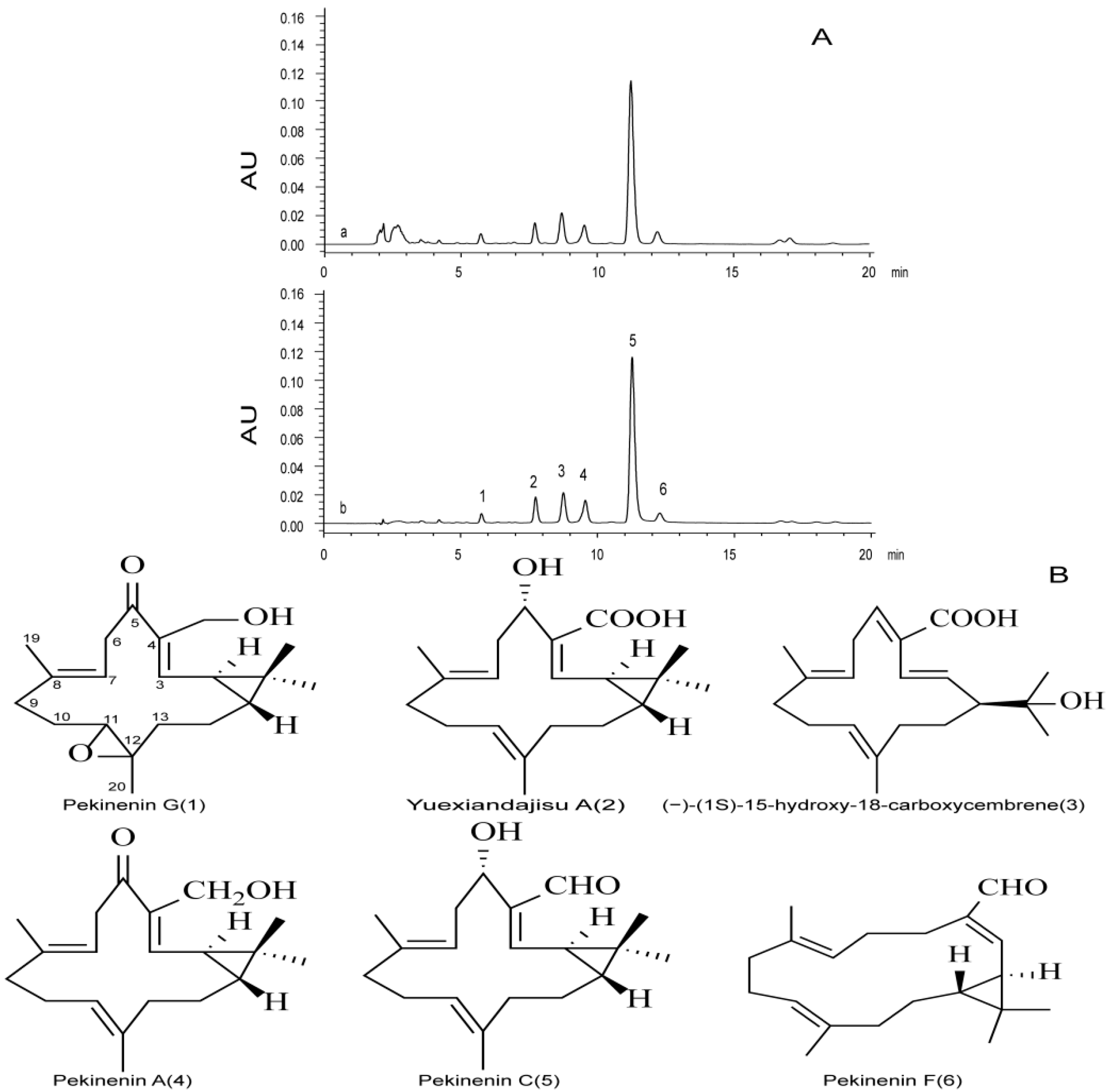
2.1. Analysis of TDEP
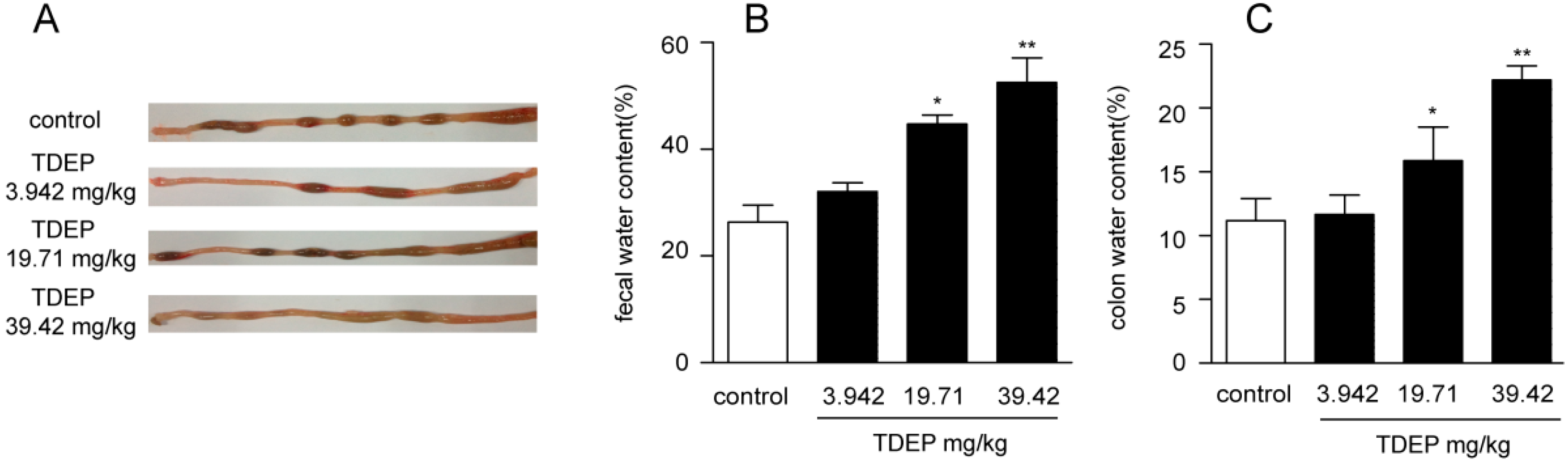
2.2. Effect of TDEP on Fecal and Intestinal Water Contents
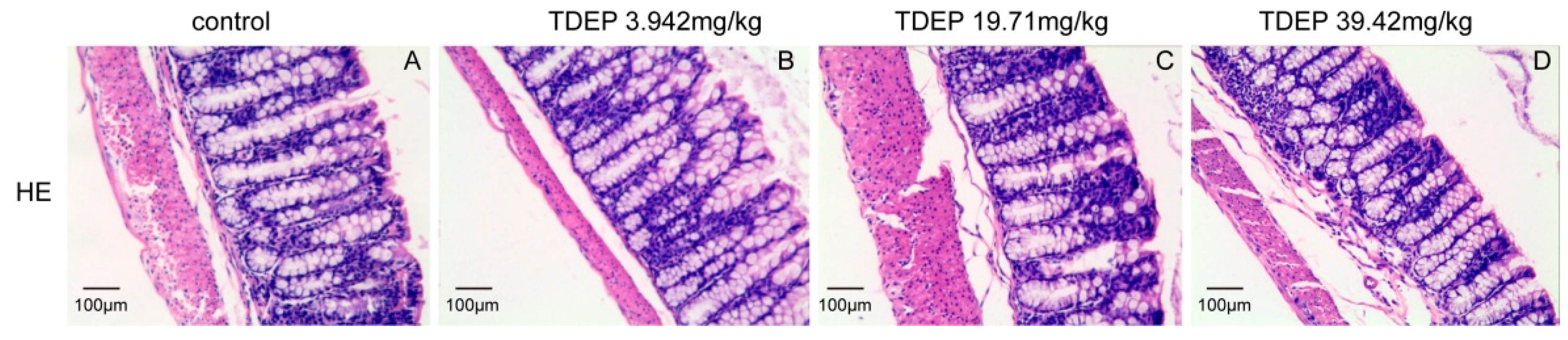
2.3. Histological Results
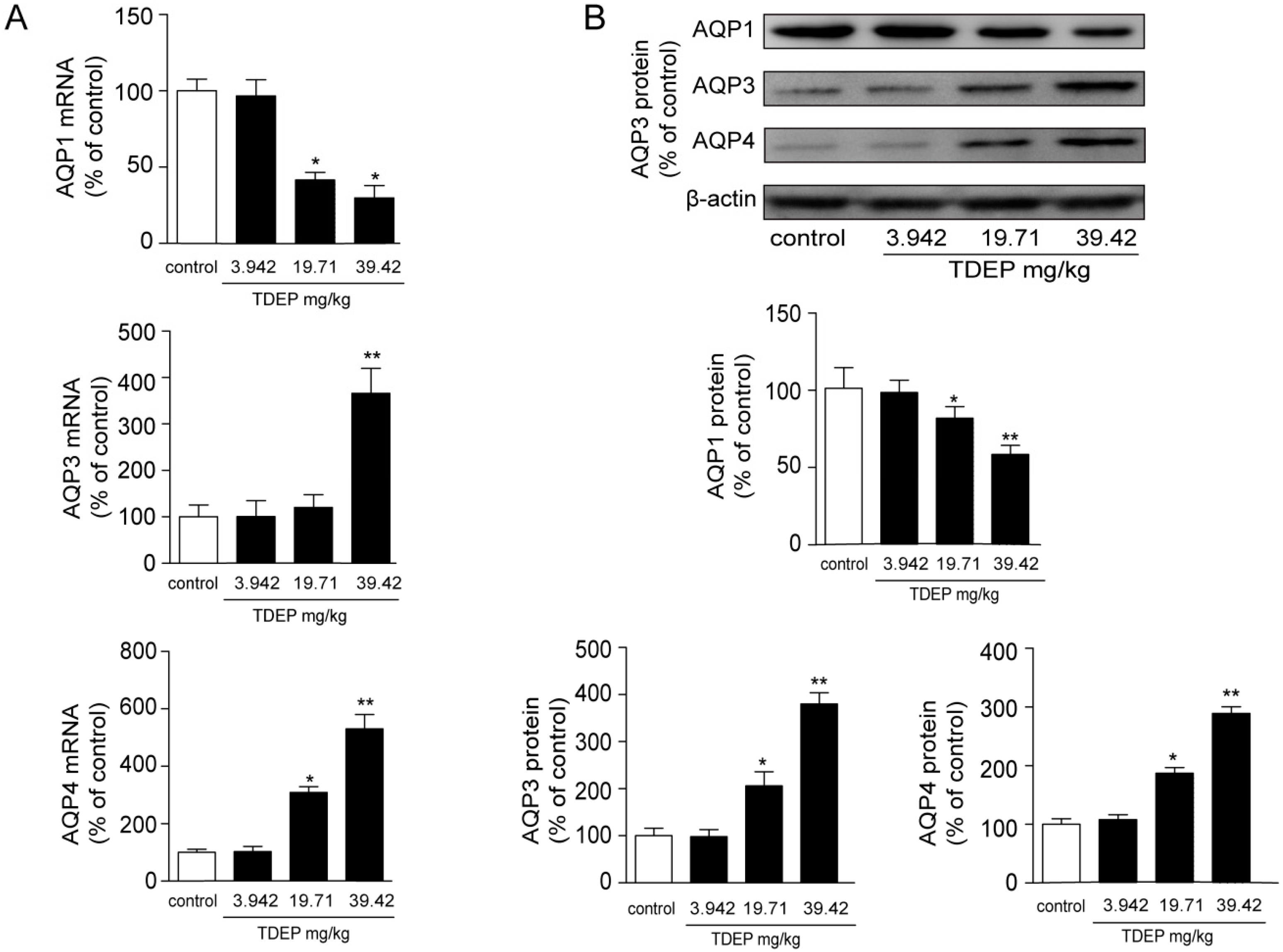
2.4. Effects of TDEP on mRNA and Protein Expression Levels of AQPs in the Colon
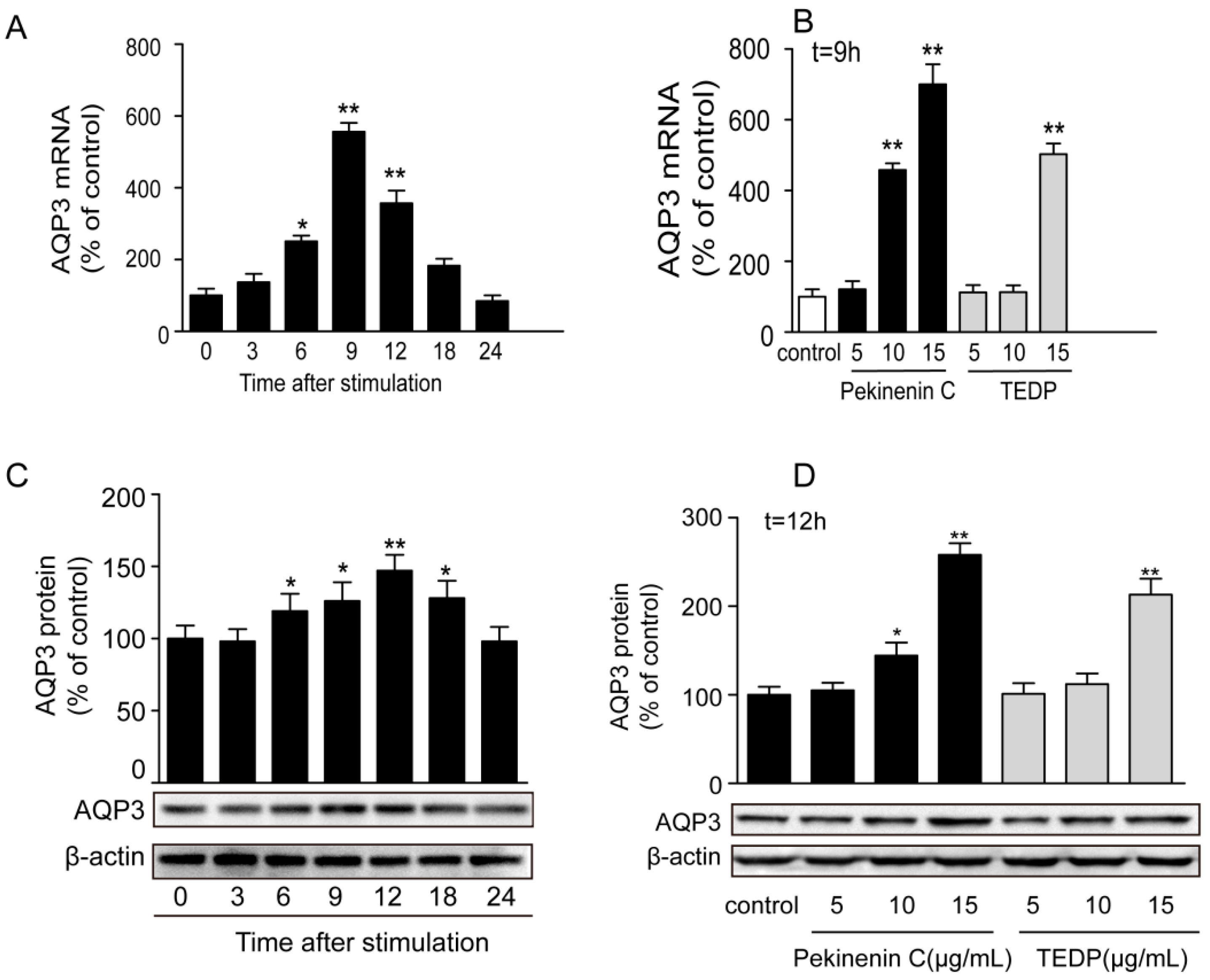
2.5. Pekinenin Cand TDEP Increased Gene and Protein Expressions of AQP3 in HT-29 Cells
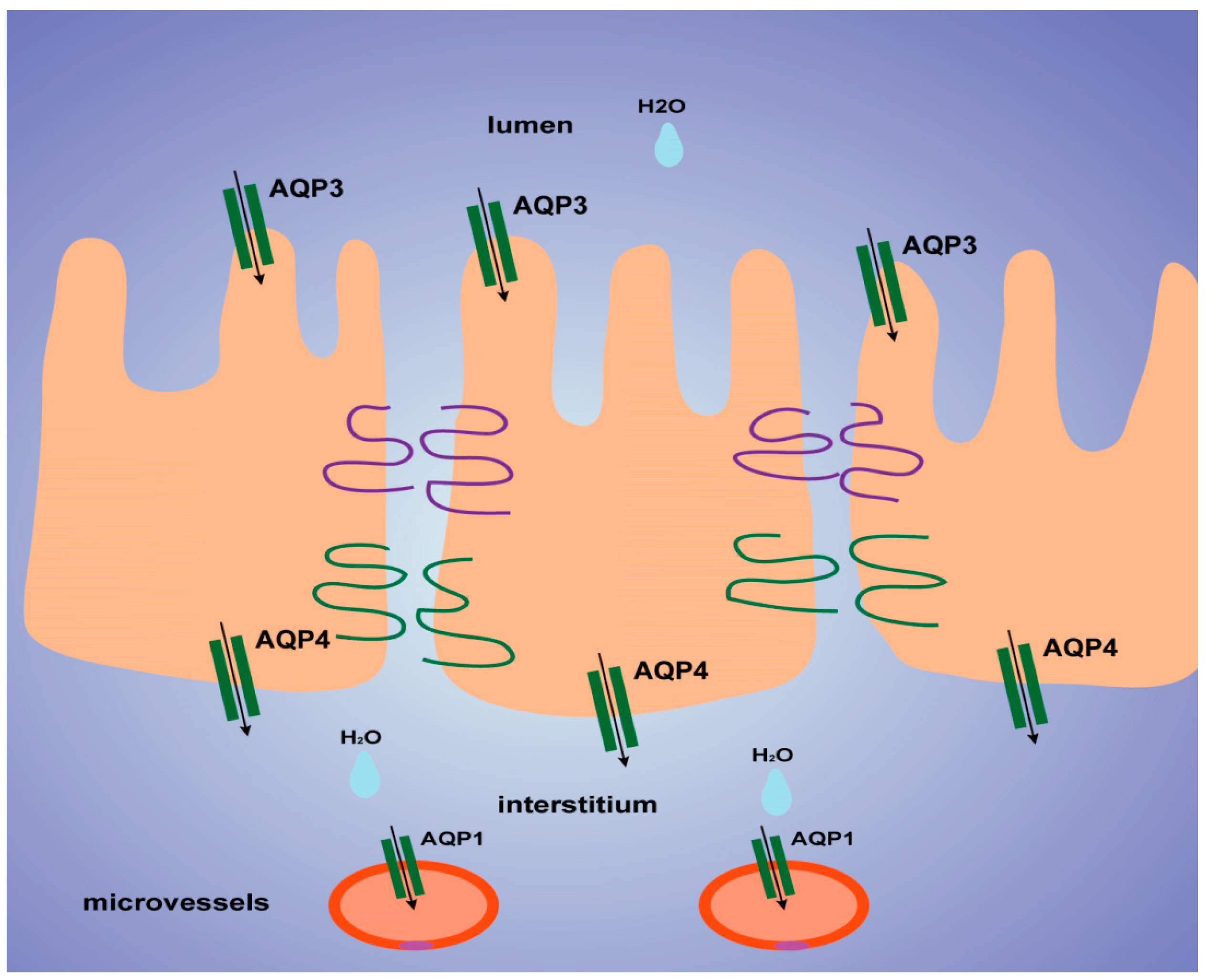
3. Discussion
4. Experimental Section
4.1. Preparation of TDEP
4.2. Determination of TDEP
4.3. Animals and Treatment
4.4. Feces Water Content
4.5. Colon Water Content
4.6. Histopathological Examination
4.7. Cell Culture
4.8. RNA Preparation and SYBR Green Real-Time RT-PCR
4.9. Preparation of Tissue and Cell Extraction for Western Blot
4.10. Electrophoresis and Immunoblotting
4.11. Statistical Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- King, L.S.; Kozono, D.; Agre, P. From structure to disease: The evolving tale of aquaporin biology. Nat. Rev. Mol. Cell Biol. 2004, 5, 687–698. [Google Scholar] [CrossRef] [PubMed]
- Laforenza, U. Water channel proteins in the gastrointestinal tract. Mol. Aspects Med. 2012, 33, 642–650. [Google Scholar] [CrossRef] [PubMed]
- Yde, J.; Keely, S.; Wu, Q.; Borg, J.F.; Lajczak, N.; O’Dwyer, A.; Dalsgaard, P.; Fenton, R.A.; Moeller, H.B. Characterization of AQPs in Mouse, Rat, and Human Colon and Their Selective Regulation by Bile Acids. Front. Nutr. 2016, 46, 3–20. [Google Scholar] [CrossRef] [PubMed]
- Ikarashi, N.; Mochiduki, T.; Takasaki, A.; Ushiki, T.; Baba, K.; Ishii, M.; Kudo, T.; Ito, K.; Toda, T.; Ochiai, W.; et al. A mechanism by which the osmotic laxative magnesium sulphate increases the intestinal aquaporin 3 expression in HT-29 cells. Life Sci. 2011, 88, 194–200. [Google Scholar] [CrossRef] [PubMed]
- Jiangsu College of New Medicine. The Dictionary of the Traditional Chinese Medicine; Shanghai Press of Science Technology: Shanghai, China, 1977; pp. 108–110. [Google Scholar]
- Tian, R.; Lu, Y.; Chen, D. Four new diterpenoids from the roots of Euphorbia pekinensis. Chem. Biodivers. 2016, 13, 1404–1409. [Google Scholar] [CrossRef] [PubMed]
- Liang, Q.; Dai, C.; Jiang, J.; Tang, Y.; Duan, J. A new cytotoxic casbane diterpene from Euphorbia pekinensis. Fitoterapia 2009, 80, 514–516. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Yu, H.; Wu, H.; Wang, X.; Pan, Y.; Chen, Y.; Liu, L.; Jin, Y.; Zhang, C. A new casbane diterpene from Euphorbia pekinensis. Nat. Prod. Res. 2015, 29, 1456–1460. [Google Scholar] [CrossRef] [PubMed]
- Tao, W.; Duan, J.; Tang, Y.; Yang, N.; Li, J.; Qian, Y. Casbane diterpenoids from the roots of Euphorbia pekinensis. Phytochemistry 2013, 94, 249–253. [Google Scholar] [CrossRef] [PubMed]
- Shao, F.; Bu, R.; Zhang, C.; Chen, C.; Huang, J.; Wang, J. Two new casbane diterpenoids from the roots of Euphorbia pekinensis. J. Asian Nat. Prod. Res. 2011, 13, 805–810. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Cheng, F.; Yao, W.; Bao, B.; Zhang, K.; Zhang, L.; Ding, A. Toxicity of Pekinenin C from Euphorbia Pekinensis Radix on Rat Small Intestinal Crypt Epithelial Cell and Its Apoptotic Mechanism. Int. J. Mol. Sci. 2016, 17, 850. [Google Scholar] [CrossRef] [PubMed]
- Turner, J.R. Intestinal mucosal barrier function in health and disease. Nat. Rev. Immunol. 2009, 9, 799–809. [Google Scholar] [CrossRef] [PubMed]
- Loo, D.D.F.; Wright, E.M.; Zeuthen, T. Water pumps. J. Physiol. 2002, 542, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Fischbarg, J. Fluid transport across leaky epithelia: Central role of the tight junction and supporting role of aquaporins. Physiol. Rev. 2010, 90, 1271–1290. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Chen, Z.; Jiang, Z. Expression, Distribution and Role of Aquaporin Water Channels in Human and Animal Stomach and Intestines. Int. J. Mol. Sci. 2016, 17, 1399. [Google Scholar] [CrossRef] [PubMed]
- Pelagalli, A.; Squillacioti, C.; Mirabella, N.; Meli, R. Aquaporins in Health and Disease: An Overview Focusing on the Gut of Different Species. Int. J. Mol. Sci. 2016, 17, 1213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koyama, Y.; Yamamoto, T.; Tani, T.; Nihei, K.; Kondo, D.; Funaki, H.; Yaoita, E.; Kawasaki, K.; Sato, N.; Hatakeyama, K.; et al. Expression and localization of aquaporins in rat gastrointestinal tract. Am. J. Physiol. 1999, 276, C621–C627. [Google Scholar] [PubMed]
- Wang, K.S.; Ma, T.; Filiz, F.; Verkman, A.S.; Bastidas, J.A. Colon water transport in transgenic mice lacking aquaporin-4 water channels. Am. J. Physiol. Gastrointest. Liver Physiol. 2000, 279, 463–470. [Google Scholar]
- Ikarashi, N.; Ushiki, T.; Mochizuki, T.; Toda, T.; Kudo, T.; Baba, K.; Ishii, M.; Ito, K.; Ochiai, W.; Sugiyama, K. Effects of Magnesium Sulphate Administration on Aquaporin 3 in Rat. Gastrointest. Tract. Biol. Phar. Bull. 2011, 34, 238–242. [Google Scholar] [CrossRef]
- Ikarashi, N.; Kon, R.; Iizasa, T.; Suzuki, N.; Hiruma, R.; Suenaga, K.; Toda, T.; Ishii, M.; Hoshino, M.; Ochiai, W.; et al. Inhibition of Aquaporin-3 Water Channel in the Colon Induces Diarrhea. Biol. Phar. Bull. 2012, 35, 957–962. [Google Scholar] [CrossRef]
- Xu, Z.H.; Sun, J.; Xu, R.S.; Qin, G.W. Casbane diterpenoids from Euphorbia ebracteolata. Phytochemistry 1998, 49, 149–151. [Google Scholar] [CrossRef]
- Chen, H.; Jia, Z.J. Diterpenes and Sequiterpene from two Euphorbia species-Structure elucidation of Euphorhylonal A and Euphoranin E. Indian J. Chem. 1996, 35B, 1308–1310. [Google Scholar]
- Ikarashi, N.; Kon, R.; Sugiyama, K. Aquaporins in the Colon as a New Therapeutic Target in Diarrhea and Constipation. Int. J. Mol. Sci. 2016, 17, 1172. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the compounds are available from the authors.







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward (5—3) | Reverse (5—3) | Amplicon Size (bp) |
|---|---|---|---|
| Mouse AQP1 | CTGCTGGCGATTGACTACAC | TCCACACCTTCATGCGGTC | 190 |
| Mouse AQP3 | GTGATGTTTGGCTGTGGCTC | TTCAAGTGGGCACCAGACAC | 184 |
| Mouse AQP4 | AAGGCGGTGGGGTAAGTG | TGAGCCACCCCAGTTTATGG | 187 |
| Mouse β-actin | GACCCAGATCATGTTTGAGAC | GTAGCCACGCTCGGTCAG | 168 |
| Human AQP3 | CTGGTGGTCCTGGTCATTG | CACATTCTCTTCCTCGTTGG | 163 |
| Human β-actin | GACCCAGATCATGTTTGAGAC | GTAGCCACGCTCGGTCAG | 179 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, K.; Liu, L.; Huang, J.; Yu, H.; Wu, H.; Duan, Y.; Cui, X.; Zhang, X.; Liu, L.; Wang, W. Laxative Effects of Total Diterpenoids Extracted from the Roots of Euphorbia pekinensis Are Attributable to Alterations of Aquaporins in the Colon. Molecules 2017, 22, 465. https://doi.org/10.3390/molecules22030465
Wang K, Liu L, Huang J, Yu H, Wu H, Duan Y, Cui X, Zhang X, Liu L, Wang W. Laxative Effects of Total Diterpenoids Extracted from the Roots of Euphorbia pekinensis Are Attributable to Alterations of Aquaporins in the Colon. Molecules. 2017; 22(3):465. https://doi.org/10.3390/molecules22030465
Chicago/Turabian StyleWang, Kuilong, Lian Liu, Jianyu Huang, Hongli Yu, Hao Wu, Yu Duan, Xiaobing Cui, Xingde Zhang, Liping Liu, and Wei Wang. 2017. "Laxative Effects of Total Diterpenoids Extracted from the Roots of Euphorbia pekinensis Are Attributable to Alterations of Aquaporins in the Colon" Molecules 22, no. 3: 465. https://doi.org/10.3390/molecules22030465
APA StyleWang, K., Liu, L., Huang, J., Yu, H., Wu, H., Duan, Y., Cui, X., Zhang, X., Liu, L., & Wang, W. (2017). Laxative Effects of Total Diterpenoids Extracted from the Roots of Euphorbia pekinensis Are Attributable to Alterations of Aquaporins in the Colon. Molecules, 22(3), 465. https://doi.org/10.3390/molecules22030465