
On the Emerging Role of the Taste Receptor Type 1 (T1R) Family of Nutrient-Sensors in the Musculoskeletal System


Abstract
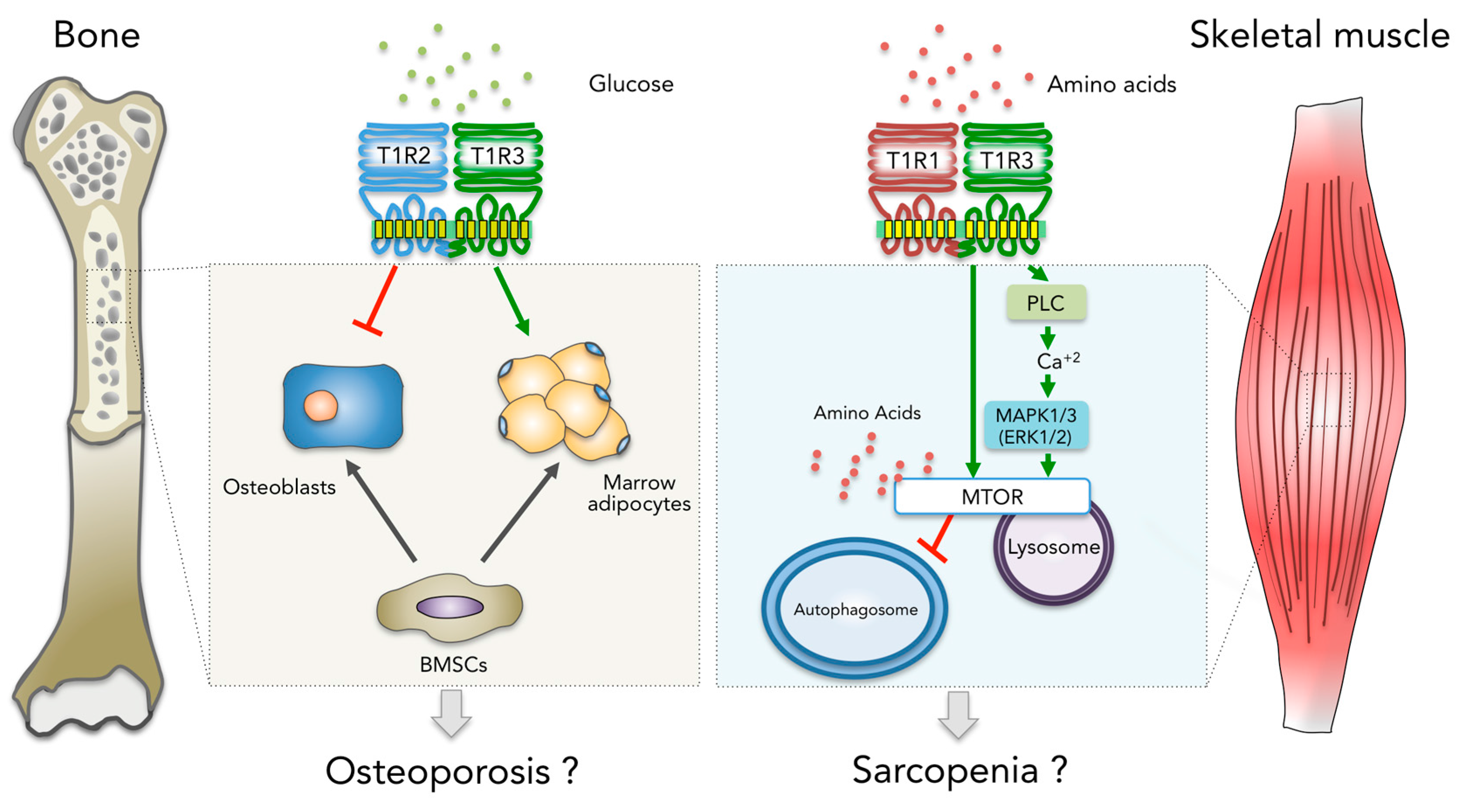
:1. Introduction
2. Sarcopenia and the Role of the T1R Family in Myogenesis
3. Osteoporosis and the Role of the T1R Family in Postnatal Bone Remodeling
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Loper, H.B.; La Sala, M.; Dotson, C.; Steinle, N. Taste perception, associated hormonal modulation, and nutrient intake. Nutr. Rev. 2015, 73, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Finger, T.E.; Kinnamon, S.C. Taste isn’t just for taste buds anymore. F1000 Biol. Rep. 2011, 3, 20. [Google Scholar] [CrossRef] [PubMed]
- Schild, L. The epithelial sodium channel and the control of sodium balance. BBA-Mol. Basis Dis. 2010, 1802, 1159–1165. [Google Scholar] [CrossRef] [PubMed]
- Matsunami, H.; Montmayeur, J.P.; Buck, L.B. A family of candidate taste receptors in human and mouse. Nature 2000, 404, 601–604. [Google Scholar] [PubMed]
- Hoon, M.A.; Adler, E.; Lindemeier, J.; Battey, J.F.; Ryba, N.J.; Zuker, C.S. Putative mammalian taste receptors: A class of taste-specific GPCRs with distinct topographic selectivity. Cell 1999, 96, 541–551. [Google Scholar] [CrossRef]
- Adler, E.; Hoon, M.A.; Mueller, K.L.; Chandrashekar, J.; Ryba, N.J.; Zuker, C.S. A novel family of mammalian taste receptors. Cell 2000, 100, 693–702. [Google Scholar] [CrossRef]
- Nelson, G.; Hoon, M.A.; Chandrashekar, J.; Zhang, Y.; Ryba, N.J.; Zuker, C.S. Mammalian sweet taste receptors. Cell 2001, 106, 381–390. [Google Scholar] [CrossRef]
- Nelson, G.; Chandrashekar, J.; Hoon, M.A.; Feng, L.; Zhao, G.; Ryba, N.J.; Zuker, C.S. An amino-acid taste receptor. Nature 2002, 416, 199–202. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.Q.; Zhang, Y.; Hoon, M.A.; Chandrashekar, J.; Erlenbach, I.; Ryba, N.J.; Zuker, C.S. The receptors for mammalian sweet and umami taste. Cell 2003, 115, 255–266. [Google Scholar] [CrossRef]
- Andres-Barquin, P.J.; Conte, C. Molecular basis of bitter taste: The T2R family of G protein-coupled receptors. Cell Biochem. Biophys. 2004, 41, 99–112. [Google Scholar] [CrossRef]
- Li, X.; Staszewski, L.; Xu, H.; Durick, K.; Zoller, M.; Adler, E. Human receptors for sweet and umami taste. Proc. Natl. Acad. Sci. USA 2002, 99, 4692–4696. [Google Scholar] [CrossRef] [PubMed]
- Hofer, D.; Puschel, B.; Drenckhahn, D. Taste receptor-like cells in the rat gut identified by expression of α-gustducin. Proc. Natl. Acad. Sci. USA 1996, 93, 6631–6634. [Google Scholar] [CrossRef] [PubMed]
- Oya, M.; Suzuki, H.; Watanabe, Y.; Sato, M.; Tsuboi, T. Amino acid taste receptor regulates insulin secretion in pancreatic β-cell line min6 cells. Genes Cells 2011, 16, 608–616. [Google Scholar] [CrossRef] [PubMed]
- Masubuchi, Y.; Nakagawa, Y.; Ma, J.; Sasaki, T.; Kitamura, T.; Yamamoto, Y.; Kurose, H.; Kojima, I.; Shibata, H. A novel regulatory function of sweet taste-sensing receptor in adipogenic differentiation of 3T3-L1 cells. PLoS ONE 2013, 8, e54500. [Google Scholar] [CrossRef] [PubMed]
- Foster, S.R.; Porrello, E.R.; Purdue, B.; Chan, H.W.; Voigt, A.; Frenzel, S.; Hannan, R.D.; Moritz, K.M.; Simmons, D.G.; Molenaar, P.; et al. Expression, regulation and putative nutrient-sensing function of taste GPCRs in the heart. PLoS ONE 2013, 8, e64579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, X.; Zhou, L.; Terwilliger, R.; Newton, S.S.; De Araujo, I.E. Sweet taste signaling functions as a hypothalamic glucose sensor. Front. Integr. Neurosci. 2009, 3, 12. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, T.; Wright, G.; Amrein, H. Nutrient sensors. Curr. Biol. 2013, 23, R369–R373. [Google Scholar] [CrossRef] [PubMed]
- Laffitte, A.; Neiers, F.; Briand, L. Functional roles of the sweet taste receptor in oral and extraoral tissues. Curr. Opin. Clin. Nutr. Metab. Care 2014, 17, 379–385. [Google Scholar] [CrossRef] [PubMed]
- Simon, B.R.; Learman, B.S.; Parlee, S.D.; Scheller, E.L.; Mori, H.; Cawthorn, W.P.; Ning, X.; Krishnan, V.; Ma, Y.L.; Tyrberg, B.; et al. Sweet taste receptor deficient mice have decreased adiposity and increased bone mass. PLoS ONE 2014, 9, e86454. [Google Scholar] [CrossRef] [PubMed]
- Kokabu, S.; Lowery, J.W.; Toyono, T.; Seta, Y.; Hitomi, S.; Sato, T.; Enoki, Y.; Okubo, M.; Fukushima, Y.; Yoda, T. Muscle regulatory factors regulate T1R3 taste receptor expression. Biochem. Biophys. Res. Commun. 2015, 468, 568–573. [Google Scholar] [CrossRef] [PubMed]
- Wauson, E.M.; Zaganjor, E.; Lee, A.Y.; Guerra, M.L.; Ghosh, A.B.; Bookout, A.L.; Chambers, C.P.; Jivan, A.; McGlynn, K.; Hutchison, M.R.; et al. The G protein-coupled taste receptor T1R1/T1R3 regulates mTORC1 and autophagy. Mol. Cell 2012, 47, 851–862. [Google Scholar] [CrossRef] [PubMed]
- Eaton, M.S.; Newby, J.B.; Plattes, M.; Foster, H.; Wauson, E.M.; Dewar, B.; Arthur, J.; Lowery, J.W. Loss of the nutrient sensor Tas1R3 leads to reduced bone resorption. In Proceedings of the American Society for Bone and Mineral Research Annual Meeting, Seattle, WA, USA, 9–12 October 2015.
- Yoshimura, N.; Nakamura, K. Epidemiology of locomotive organ disorders and symptoms: An estimation using the population-based cohorts in japan. Clinical Rev. Bone Miner. Metab. 2016, 14, 68–73. [Google Scholar] [CrossRef] [PubMed]
- Delmonico, M.J.; Harris, T.B.; Lee, J.S.; Visser, M.; Nevitt, M.; Kritchevsky, S.B.; Tylavsky, F.A.; Newman, A.B.; Health, A.; Body Composition, S. Alternative definitions of sarcopenia, lower extremity performance, and functional impairment with aging in older men and women. J. Am. Geriatr. Soc. 2007, 55, 769–774. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Jentoft, A.J.; Baeyens, J.P.; Bauer, J.M.; Boirie, Y.; Cederholm, T.; Landi, F.; Martin, F.C.; Michel, J.P.; Rolland, Y.; Schneider, S.M.; et al. Sarcopenia: European consensus on definition and diagnosis: Report of the european working group on sarcopenia in older people. Age Ageing 2010, 39, 412–423. [Google Scholar] [CrossRef] [PubMed]
- Janssen, I.; Shepard, D.S.; Katzmarzyk, P.T.; Roubenoff, R. The healthcare costs of sarcopenia in the united states. J Am. Geriatr. Soc. 2004, 52, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Chakkalakal, J.V.; Jones, K.M.; Basson, M.A.; Brack, A.S. The aged niche disrupts muscle stem cell quiescence. Nature 2012, 490, 355–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sousa-Victor, P.; Gutarra, S.; Garcia-Prat, L.; Rodriguez-Ubreva, J.; Ortet, L.; Ruiz-Bonilla, V.; Jardi, M.; Ballestar, E.; Gonzalez, S.; Serrano, A.L.; et al. Geriatric muscle stem cells switch reversible quiescence into senescence. Nature 2014, 506, 316–321. [Google Scholar] [CrossRef] [PubMed]
- Sousa-Victor, P.; Garcia-Prat, L.; Serrano, A.L.; Perdiguero, E.; Munoz-Canoves, P. Muscle stem cell aging: Regulation and rejuvenation. Trends. Endocrinol. Metab. 2015, 26, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Price, F.D.; von Maltzahn, J.; Bentzinger, C.F.; Dumont, N.A.; Yin, H.; Chang, N.C.; Wilson, D.H.; Frenette, J.; Rudnicki, M.A. Inhibition of jak-stat signaling stimulates adult satellite cell function. Nat. Med. 2014, 20, 1174–1181. [Google Scholar] [CrossRef] [PubMed]
- Tierney, M.T.; Aydogdu, T.; Sala, D.; Malecova, B.; Gatto, S.; Puri, P.L.; Latella, L.; Sacco, A. STAT3 signaling controls satellite cell expansion and skeletal muscle repair. Nat. Med. 2014, 20, 1182–1186. [Google Scholar] [CrossRef] [PubMed]
- Bernet, J.D.; Doles, J.D.; Hall, J.K.; Kelly Tanaka, K.; Carter, T.A.; Olwin, B.B. P38 MAPK signaling underlies a cell-autonomous loss of stem cell self-renewal in skeletal muscle of aged mice. Nat. Med. 2014, 20, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Cuervo, A.M.; Bergamini, E.; Brunk, U.T.; Droge, W.; Ffrench, M.; Terman, A. Autophagy and aging: The importance of maintaining “clean” cells. Autophagy 2005, 1, 131–140. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Klionsky, D.J. Regulation mechanisms and signaling pathways of autophagy. Annu. Rev. Genet. 2009, 43, 67–93. [Google Scholar] [CrossRef] [PubMed]
- Carnio, S.; LoVerso, F.; Baraibar, M.A.; Longa, E.; Khan, M.M.; Maffei, M.; Reischl, M.; Canepari, M.; Loefler, S.; Kern, H.; et al. Autophagy impairment in muscle induces neuromuscular junction degeneration and precocious aging. Cell Rep. 2014, 8, 1509–1521. [Google Scholar] [CrossRef] [PubMed]
- Rubinsztein, D.C.; Marino, G.; Kroemer, G. Autophagy and aging. Cell 2011, 146, 682–695. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Prat, L.; Martinez-Vicente, M.; Perdiguero, E.; Ortet, L.; Rodriguez-Ubreva, J.; Rebollo, E.; Ruiz-Bonilla, V.; Gutarra, S.; Ballestar, E.; Serrano, A.L.; et al. Autophagy maintains stemness by preventing senescence. Nature 2016, 529, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Jaenisch, R. Myogenin can substitute for Myf5 in promoting myogenesis but less efficiently. Development 1997, 124, 2507–2513. [Google Scholar] [PubMed]
- Zhu, Z.; Miller, J.B. Mrf4 can substitute for myogenin during early stages of myogenesis. Dev. Dyn. 1997, 209, 233–241. [Google Scholar] [CrossRef]
- Lassar, A.B.; Skapek, S.X.; Novitch, B. Regulatory mechanisms that coordinate skeletal muscle differentiation and cell cycle withdrawal. Curr. Opin. Cell Biol. 1994, 6, 788–794. [Google Scholar] [CrossRef]
- Pownall, M.E.; Gustafsson, M.K.; Emerson, C.P., Jr. Myogenic regulatory factors and the specification of muscle progenitors in vertebrate embryos. Annu. Rev. Cell Dev. Bi. 2002, 18, 747–783. [Google Scholar] [CrossRef] [PubMed]
- Yun, K.; Wold, B. Skeletal muscle determination and differentiation: Story of a core regulatory network and its context. Curr. Opin. Cell Biol. 1996, 8, 877–889. [Google Scholar] [CrossRef]
- Wauson, E.M.; Lorente-Rodriguez, A.; Cobb, M.H. Minireview: Nutrient sensing by G protein-coupled receptors. Mol. Endocrinol 2013, 27, 1188–1197. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, N.; Yoshikawa, N.; Ito, N.; Maruyama, T.; Suzuki, Y.; Takeda, S.; Nakae, J.; Tagata, Y.; Nishitani, S.; Takehana, K.; et al. Crosstalk between glucocorticoid receptor and nutritional sensor mtor in skeletal muscle. Cell Metab. 2011, 13, 170–182. [Google Scholar] [CrossRef] [PubMed]
- Edwards, M.H.; Dennison, E.M.; Aihie Sayer, A.; Fielding, R.; Cooper, C. Osteoporosis and sarcopenia in older age. Bone 2015, 80, 126–130. [Google Scholar] [CrossRef] [PubMed]
- Wright, N.C.; Looker, A.C.; Saag, K.G.; Curtis, J.R.; Delzell, E.S.; Randall, S.; Dawson-Hughes, B. The recent prevalence of osteoporosis and low bone mass in the united states based on bone mineral density at the femoral neck or lumbar spine. J. Bone Miner. Res. 2014, 29, 2520–2526. [Google Scholar] [CrossRef] [PubMed]
- Blume, S.W.; Curtis, J.R. Medical costs of osteoporosis in the elderly medicare population. Osteoporos. Int. 2011, 22, 1835–1844. [Google Scholar] [CrossRef] [PubMed]
- Kokabu, S.; Tsuchiya-Hirata, S.; Fukushima, H.; Sugiyama, G.; Lowery, J.W.; Katagiri, T.; Jimi, E. Inhibition of bone morphogenetic protein-induced osteoblast differentiation. J.Oral. Biosci. 2015, 57, 179–184. [Google Scholar] [CrossRef]
- Harvey, N.; Dennison, E.; Cooper, C. Osteoporosis: Impact on health and economics. Nat. Rev. Rheumatol. 2010, 6, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Wauson, E.M.; Zaganjor, E.; Cobb, M.H. Amino acid regulation of autophagy through the GPCR TAS1R1-TAS1R3. Autophagy 2013, 9, 418–419. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| No. | Transcription Factor (Description) | No. | Transcription Factor (Description) | |
|---|---|---|---|---|
| 1 | AP2 (activator protein 2) | 22 | LXR | |
| 2 | AP2α (activator protein 2α) | 23 | MAZ | |
| 3 | AP4 (activator protein 4) | 24 | MYOD (myoblast determining factor) | |
| 4 | ATF4 (activating transcription factor 4) | 25 | MYOGENIN | |
| 5 | CEBP (CCAAT/enhancer binding protein) | 26 | NGFIC | |
| 6 | CHCH (Churchill) | 27 | R (Epstein-Barr virus transcription factor R) | |
| 7 | CP2 | 28 | RFX (X-box binding protein RFX) | |
| 8 | CREB (cAMP-response element-binding protein) | 29 | SEF1 | |
| 9 | DEAF1 | 30 | SMAD | |
| 10 | E12 | 31 | SMAD4 | |
| 11 | E2A | 32 | SP1 | |
| 12 | E2F | 33 | SPZ1 | |
| 13 | E2F1 | 34 | SREBP1 | |
| 14 | EBOX | 35 | SRF (serum response factor) | |
| 15 | EGR | 36 | SRY (sex-determining region Y gene product) | |
| 16 | FOXO4 (fork head box O4) | 37 | STAF (Se-Cys tRNA gene transcription activating factor) | |
| 17 | HEB | 38 | STRA13 | |
| 18 | HEN1 | 39 | TEF1 | |
| 19 | HSF1 (heat shock factor 1) | 40 | UF1H3β | |
| 20 | KROX | 41 | USF (upstream stimulating factor) | |
| 21 | LBP1 | 42 | ZID (zinc finger with interaction domain) | |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kokabu, S.; Lowery, J.W.; Toyono, T.; Sato, T.; Yoda, T. On the Emerging Role of the Taste Receptor Type 1 (T1R) Family of Nutrient-Sensors in the Musculoskeletal System. Molecules 2017, 22, 469. https://doi.org/10.3390/molecules22030469
Kokabu S, Lowery JW, Toyono T, Sato T, Yoda T. On the Emerging Role of the Taste Receptor Type 1 (T1R) Family of Nutrient-Sensors in the Musculoskeletal System. Molecules. 2017; 22(3):469. https://doi.org/10.3390/molecules22030469
Chicago/Turabian StyleKokabu, Shoichiro, Jonathan W. Lowery, Takashi Toyono, Tsuyoshi Sato, and Tetsuya Yoda. 2017. "On the Emerging Role of the Taste Receptor Type 1 (T1R) Family of Nutrient-Sensors in the Musculoskeletal System" Molecules 22, no. 3: 469. https://doi.org/10.3390/molecules22030469
APA StyleKokabu, S., Lowery, J. W., Toyono, T., Sato, T., & Yoda, T. (2017). On the Emerging Role of the Taste Receptor Type 1 (T1R) Family of Nutrient-Sensors in the Musculoskeletal System. Molecules, 22(3), 469. https://doi.org/10.3390/molecules22030469