
Polyphenolic Compounds and Digestive Enzymes: In Vitro Non-Covalent Interactions

 , and
, and 
Abstract
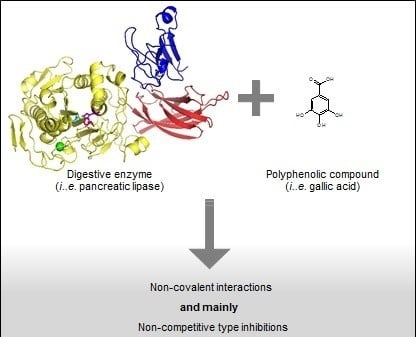
:
1. Introduction
2. Results
2.1. Digestive Enzymes
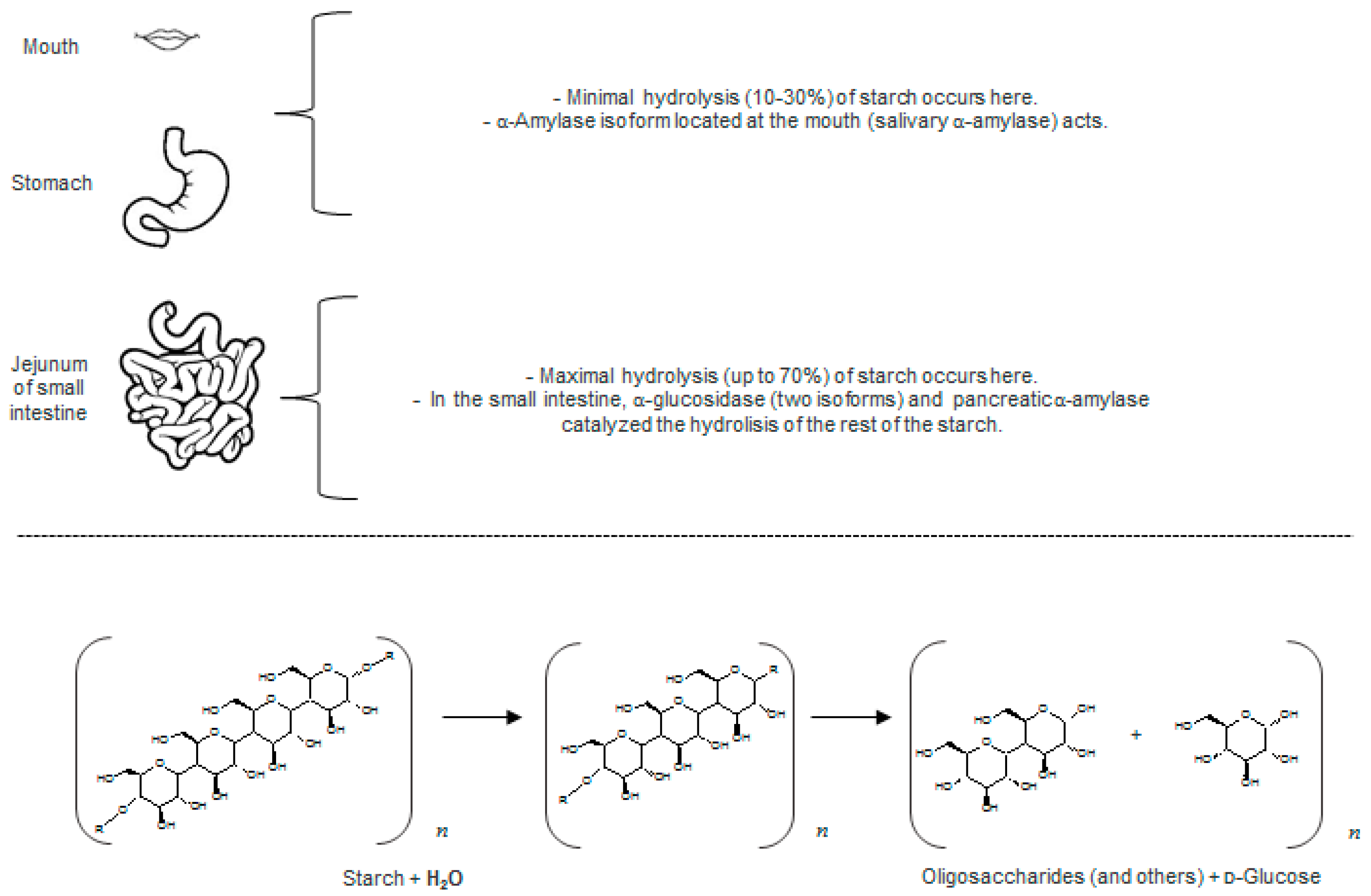
2.1.1. Glucosidase and Amylase Enzymes
2.1.2. Lipase Enzyme
2.1.3. Proteases Enzymes
2.2. Inhibition of Digestive Enzymes by Polyphenolic Compounds
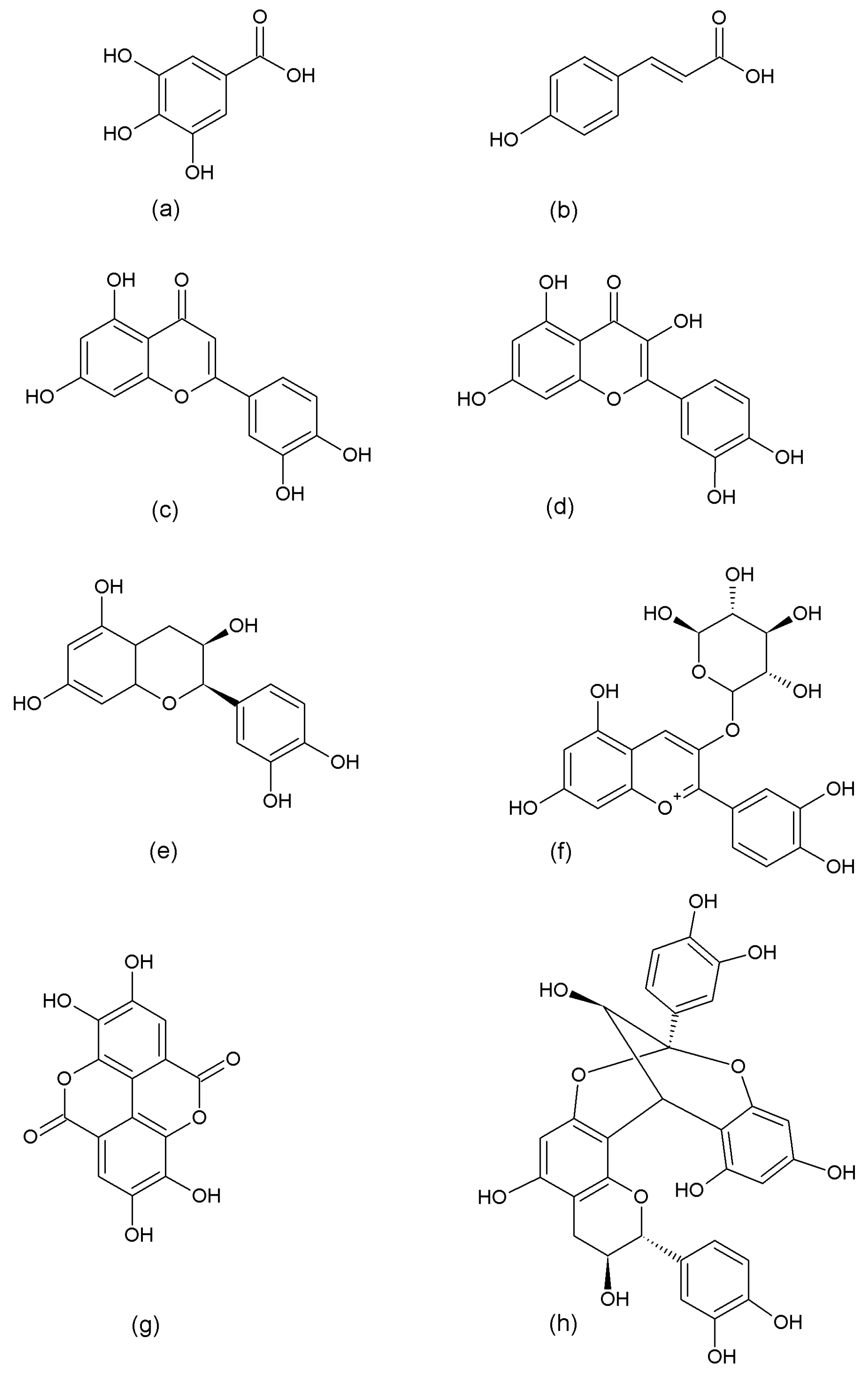
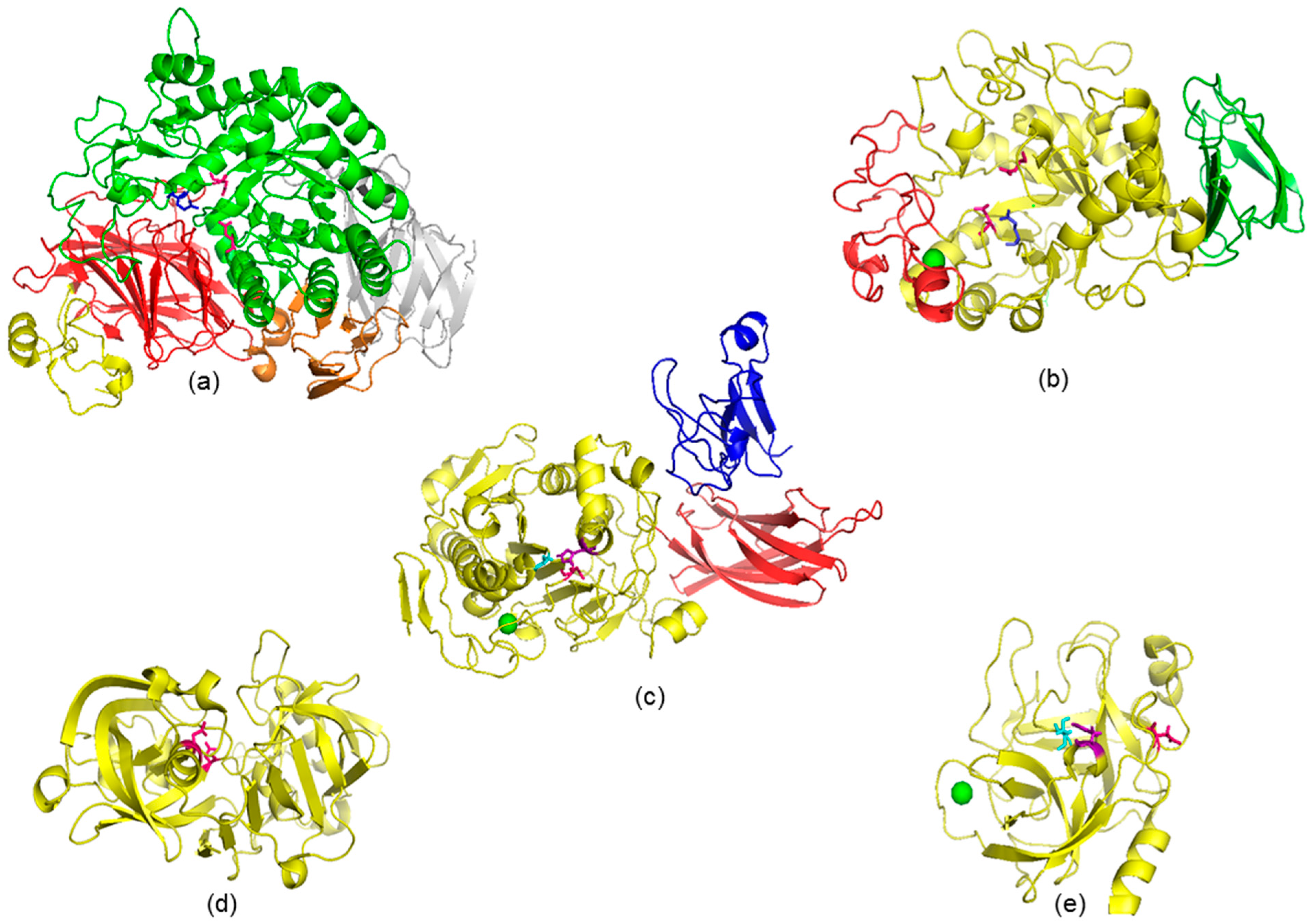
2.3. Structural and Thermodynamic Characterization of Polyphenolic Compounds–Protein Interactions
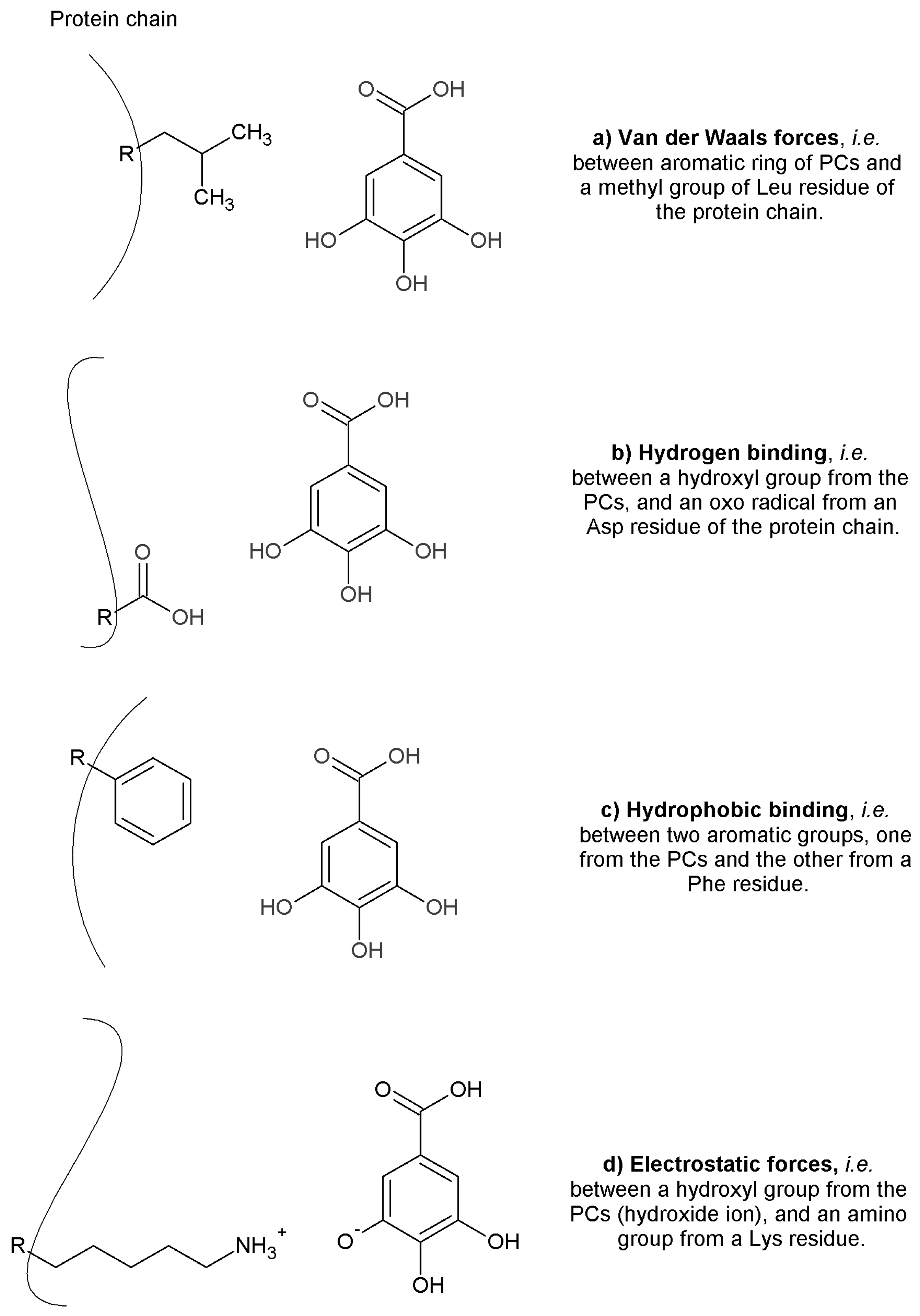
2.4. Mechanisms of Enzymatic Inhibition
2.5. Conditions Influence the Binding of Polyphenolic Compounds with Digestive Enzymes
3. Conclusions
Acknowledgments
Conflicts of Interest
References
- Huang, W.-Y.; Cai, Y.-Z.; Zhang, Y. Natural Phenolic Compounds From Medicinal Herbs and Dietary Plants: Potential Use for Cancer Prevention. Nutr. Cancer 2009, 62, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Acosta-Estrada, B.A.; Gutiérrez-Uribe, J.A.; Serna-Saldívar, S.O. Bound phenolics in foods, a review. Food Chem. 2014, 152, 46–55. [Google Scholar] [CrossRef] [PubMed]
- González-Castejón, M.; Rodriguez-Casado, A. Dietary phytochemicals and their potential effects on obesity: A review. Pharmacol. Res. 2011, 64, 438–455. [Google Scholar] [CrossRef] [PubMed]
- Buchholz, T.; Melzig, M.F. Polyphenolic Compounds as Pancreatic Lipase Inhibitors. Planta Med. 2015, 81, 771–783. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Luo, Q.; Sun, M.; Corke, H. Antioxidant activity and phenolic compounds of 112 traditional Chinese medicinal plants associated with anticancer. Life Sci. 2004, 74, 2157–2184. [Google Scholar] [CrossRef] [PubMed]
- Calabrone, L.; Larocca, M.; Marzocco, S.; Martelli, G.; Rossano, R. Total Phenols and Flavonoids Content, Antioxidant Capacity and Lipase Inhibition of Root and Leaf Horseradish (Armoracia rusticana) Extracts. Food Nutr. Sci. 2015, 06, 64–74. [Google Scholar] [CrossRef]
- Conte, A.; Pellegrini, S.; Tagliazucchi, D. Synergistic protection of PC12 cells from β-amyloid toxicity by resveratrol and catechin. Brain Res. Bull. 2003, 62, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Cruz, S.; Alvarez-Parrilla, E.; Laura, A.; Martínez-González, A.; Ornelas-Paz, J.; Mendoza-Wilson, A.; Gonzalez-Aguilar, G. Effect of different sanitizers on microbial, sensory and nutritional quality of fresh-cut jalapeno peppers. Am. J. Agric. Biol. Sci. 2010, 5, 331–341. [Google Scholar] [CrossRef]
- Sokmen, M.; Angelova, M.; Krumova, E.; Pashova, S.; Ivancheva, S.; Sokmen, A.; Serkedjieva, J. In vitro antioxidant activity of polyphenol extracts with antiviral properties from Geranium sanguineum L. Life Sci. 2005, 76, 2981–2993. [Google Scholar] [CrossRef] [PubMed]
- Riobó, S.P. Obesity and diabetes. Nutr. Hosp. 2013, 28 (Suppl. 5), 138–143. [Google Scholar]
- Wu, X.; He, W.; Zhang, H.; Li, Y.; Liu, Z.; He, Z. Acteoside: A lipase inhibitor from the Chinese tea Ligustrum purpurascens kudingcha. Food Chem. 2014, 142, 306–310. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, S.; Koehnlein, E.; Bracht, A.; Castoldi, R.; de Morais, G.; Baesso, M.; Peralta, R.; de Souza, C.; de Sá-Nakanishi, A.; Peralta, R. Inhibition of salivary and pancreatic α-amylases by a pinhão coat (Araucaria angustifolia) extract rich in condensed tannin. Food Res. Int. 2014, 56, 1–8. [Google Scholar] [CrossRef]
- Fasal, A.; Amin, I.; Norhaizan Mohd, E.; Chong Pei, P.; Sander, K. Hepatic genome-wide expression of lipid metabolism in diet-induced obesity rats treated with cocoa polyphenols. J. Funct. Foods 2015, 17, 969–978. [Google Scholar]
- Wang, S.; Sun, Z.; Dong, S.; Liu, Y.; Liu, Y. Molecular Interactions between (−)-Epigallocatechin Gallate Analogs and Pancreatic Lipase. PLoS ONE 2014, 9, e111143. [Google Scholar] [CrossRef] [PubMed]
- Jakobek, L. Interactions of polyphenols with carbohydrates, lipids and proteins. Food Chem. 2015, 175, 556–567. [Google Scholar] [CrossRef] [PubMed]
- Ozdal, T.; Capanoglu, E.; Altay, F. A review on protein-phenolic interactions and associated changes. Food Res. Int. 2013, 51, 954–970. [Google Scholar] [CrossRef]
- Birari, R.B.; Bhutani, K.K. Pancreatic lipase inhibitors from natural sources: Unexplored potential. Drug Discov. Today 2007, 12, 879–889. [Google Scholar] [CrossRef] [PubMed]
- McDougall, G.J.; Kulkarni, N.N.; Stewart, D. Berry polyphenols inhibit pancreatic lipase activity in vitro. Food Chem. 2009, 115, 193–199. [Google Scholar] [CrossRef]
- Miao, M.; Jiang, H.; Jiang, B.; Zhang, T.; Cui, S.W.; Jin, Z. Phytonutrients for controlling starch digestion: Evaluation of grape skin extract. Food Chem. 2014, 145, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Mnafgui, K.; Derbali, A.; Sayadi, S.; Gharsallah, N.; Elfeki, A.; Allouche, N. Anti-obesity and cardioprotective effects of cinnamic acid in high fat diet-induced obese rats. J. Food Sci. Technol. 2015, 52, 4378–4386. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Deng, Z.; Ramdath, D.D.; Tang, Y.; Chen, P.X.; Liu, R.; Liu, Q.; Tsao, R. Phenolic profiles of 20 Canadian lentil cultivars and their contribution to antioxidant activity and inhibitory effects on α-glucosidase and pancreatic lipase. Food Chem. 2015, 172, 862–872. [Google Scholar] [CrossRef] [PubMed]
- Costamagna, M.S.; Zampini, V.; Alberto, M.R.; Cuello, S.; Torres, S.; Pérez, J.; Quispe, C.; Schmeda-Hirschmann, G.; Isla, M.I. Polyphenols rich fraction from Geoffroea decorticans fruits flour affects key enzymes involved in metabolic syndrome, oxidative stress and inflammatory process. Food Chem. 2016, 190, 392–402. [Google Scholar] [CrossRef] [PubMed]
- Kushner, R.F. Weight loss strategies for treatment of obesity. Prog. Cardiovasc. Dis. 2014, 56, 465–472. [Google Scholar] [CrossRef] [PubMed]
- Bray, G.A. Drug Treatment of Obesity. Rev. Endocr. Metab. Disord. 2001, 2, 403–418. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Kong, F. Effects of tea polyphenols and different teas on pancreatic α-amylase activity in vitro. LWT-Food Sci. Technol. 2016, 66, 232–238. [Google Scholar] [CrossRef]
- Al-Asri, J.; Fazekas, E.; Lehoczki, G.; Perdih, A.; Görick, C.; Melzig, M.F.; Gyémánt, G.; Wolber, G.; Mortier, J. From carbohydrates to drug-like fragments: Rational development of novel α-amylase inhibitors. Bioorg. Med. Chem. 2015, 23, 6725–6732. [Google Scholar] [CrossRef] [PubMed]
- Sakulnarmrat, K.; Srzednicki, G.; Konczak, I. Composition and inhibitory activities towards digestive enzymes of polyphenolic-rich fractions of Davidson’s plum and quandong. LWT-Food Sci. Technol. 2014, 57, 366–375. [Google Scholar] [CrossRef]
- He, Q.; Lv, Y.; Yao, K. Effects of tea polyphenols on the activities of α-amylase, pepsin, trypsin and lipase. Food Chem. 2006, 101, 1178–1182. [Google Scholar] [CrossRef]
- Xiao, H.; Liu, B.; Mo, H.; Liang, G. Comparative evaluation of tannic acid inhibiting α-glucosidase and trypsin. Food Res. Int. 2015, 76 Pt 3, 605–610. [Google Scholar] [CrossRef]
- Gonzales, G.; Smagghe, G.; Grootaert, C.; Zotti, M.; Raes, K.; Camp, J. Flavonoid interactions during digestion, absorption, distribution and metabolism: A sequential structure—Activity/property relationship-based approach in the study of bioavailability and bioactivity. Drug Metab. Rev. 2015, 47, 175–190. [Google Scholar] [CrossRef] [PubMed]
- Hinsberger, A.; Sandhu, B.K. Digestion and absorption. Curr. Pediatr. Rev. 2004, 14, 605–611. [Google Scholar] [CrossRef]
- Helal, A.; Tagliazucchi, D.; Verzelloni, E.; Conte, A. Gastro-pancreatic release of phenolic compounds incorporated in a polyphenols-enriched cheese-curd. LWT-Food Sci. Technol. 2015, 60, 957–963. [Google Scholar] [CrossRef]
- Tenore, G.C.; Campiglia, P.; Giannetti, D.; Novellino, E. Simulated gastrointestinal digestion, intestinal permeation and plasma protein interaction of white, green, and black tea polyphenols. Food Chem. 2015, 169, 320–326. [Google Scholar] [CrossRef] [PubMed]
- Sergent, T.; Vanderstraeten, J.; Winand, J.; Beguin, P.; Schneider, Y.-J. Phenolic compounds and plant extracts as potential natural anti-obesity substances. Food Chem. 2012, 135, 68–73. [Google Scholar] [CrossRef]
- Qin, X.; Ren, L.; Yang, X.; Bai, F.; Wang, L.; Geng, P. Structures of human pancreatic α-amylase in complex with acarviostatins: Implications for drug design against type II diabetes. J. Struct. Biol. 2011, 174, 196–202. [Google Scholar] [CrossRef] [PubMed]
- Lin, A.; Lee, B.-H.; Chang, W.-J. Small intestine mucosal α-glucosidase: A missing feature of in vitro starch digestibility. Food Hydrocoll. 2016, 53, 163–171. [Google Scholar] [CrossRef]
- Bhandari, M.R.; Jong-Anurakkun, N.; Hong, G.; Kawabata, J. α-Glucosidase and α-amylase inhibitory activities of Nepalese medicinal herb Pakhanbhed (Bergenia ciliata, Haw). Food Chem. 2008, 106, 247–252. [Google Scholar] [CrossRef]
- Sim, L.; Quezada-Calvillo, R.; Sterchi, E.E.; Nichols, B.L.; Rose, D.R. Human Intestinal Maltase–Glucoamylase: Crystal Structure of the N-Terminal Catalytic Subunit and Basis of Inhibition and Substrate Specificity. J. Mol. Biol. 2008, 375, 782–792. [Google Scholar] [CrossRef] [PubMed]
- Behringer, V.; Borchers, C.; Deschner, T.; Möstl, E.; Selzer, D.; Hohmann, G. Measurements of Salivary Alpha Amylase and Salivary Cortisol in Hominoid Primates Reveal Within-Species Consistency and Between-Species Differences. PLoS ONE 2013, 8, e60773. [Google Scholar] [CrossRef] [PubMed]
- Kashtoh, H.; Muhammad, M.; Khan, J.; Rasheed, S.; Khan, A.; Perveen, S.; Javaid, K.; Atia tul, W.; Khan, K.; Choudhary, I. Dihydropyrano [2,3-c] pyrazole: Novel in vitro inhibitors of yeast α-glucosidase. Bioorg. Chem. 2016, 65, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Ferey-Roux, G.; Perrier, J.; Forest, E. The human pancreatic α-amylase isoforms: Isolation, structural studies and kinetics of inhibition by acarbose. Biochim. Biophys. Acta 1998, 1388, 10–20. [Google Scholar] [CrossRef]
- Liu, M.; Zhang, W.; Wei, J.; Lin, X. Synthesis and α-Glucosidase Inhibitory Mechanisms of Bis(2,3-dibromo-4,5-dihydroxybenzyl) Ether, a Potential Marine Bromophenol α-Glucosidase Inhibitor. Mar. Drugs 2011, 9, 1554–1565. [Google Scholar] [CrossRef] [PubMed]
- Zhuo, H.; Payan, F.; Qian, M. Crystal structure of the pig pancreatic α-amylase complexed with ρ-nitrophenyl-α-d-maltoside-flexibility in the active site. Protein J. 2004, 23, 379–387. [Google Scholar] [CrossRef] [PubMed]
- Buisson, G.; Duée, E.; Haser, R.; Payan, F. Three dimensional structure of porcine pancreatic alpha-amylase at 2.9 A resolution. Role of calcium in structure and activity. EMBO J. 1987, 6, 3909–3916. [Google Scholar] [PubMed]
- Mok, S.C.; Teh, A.; Saito, J.; Najimudin, N.; Alam, M. Crystal structure of a compact α-amylase from Geobacillus thermoleovorans. Enzyme Microb. Technol. 2013, 53, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, M. Human digestive and metabolic lipases—a brief review. J. Mol. Catal. B Enzym. 2003, 22, 369–376. [Google Scholar] [CrossRef]
- Shi, Y.; Burn, P. Lipid metabolic enzymes: Emerging drug targets for the treatment of obesity. Nat. Rev. Drug Discov. 2004, 3, 695–710. [Google Scholar] [CrossRef] [PubMed]
- Miled, N.; Canaan, S.; Dupuis, L.; Roussel, A.; Rivière, M. Digestive lipases: From three-dimensional structure to physiology. Biochimie 2000, 82, 973–976. [Google Scholar] [CrossRef]
- Egloff, M.P.; Sarda, L.; Verger, R.; Cambillau, C.; Tilbeurgh, H. Crystallographic study of the structure of colipase and of the interaction with pancreatic lipase. Protein Sci. 1995, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Van Tilbeurgh, H.; Bezzine, S.; Cambillau, C.; Verger, R.; Carrière, F. Colipase: Structure and interaction with pancreatic lipase. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 1999, 1441, 173–184. [Google Scholar] [CrossRef]
- Hermoso, J.; Pignol, D.; Kerferlec, B.; Crenon, I.; Chapus, C.; Fontecilla-Camps, J.C. Lipase Activation by Nonionic Detergents. J. Biol. Chem. 1996, 271, 18007–18016. [Google Scholar] [CrossRef] [PubMed]
- Ibarz, A.; Garvín, A.; Garza, S.; Pagán, J. Toxic effect of melanoidins from glucose–asparagine on trypsin activity. Food Chem. Toxicol. 2009, 47, 2071–2075. [Google Scholar] [CrossRef] [PubMed]
- Campos, L.; Sancho, J. The active site of pepsin is formed in the intermediate conformation dominant at mildly acidic pH. FEBS Lett. 2003, 538, 89–95. [Google Scholar] [CrossRef]
- Ma, W.; Tang, C.; Lai, L. Specificity of Trypsin and Chymotrypsin: Loop-Motion-Controlled Dynamic Correlation as a Determinant. Biophys. J. 2005, 89, 1183–1193. [Google Scholar] [CrossRef] [PubMed]
- Rawn, J.D. Bioquímica, 1st ed.; Interamericana-McGraw-Hill: Madrid, Spain, 1989; Volume I. [Google Scholar]
- Ahmad, V.; Lodhi, M.; Abbasi, M.; Choudhary, M. Kinetics study on a novel natural inhibitor of α-chymotrypsin. Fitoterapia 2008, 79, 505–508. [Google Scholar] [CrossRef] [PubMed]
- Dechakhamphu, A.; Wongchum, N. Screening for anti-pancreatic lipase properties of 28 traditional Thai medicinal herbs. Asian Pac. J. Trop. Biomed. 2015, 5, 1042–1045. [Google Scholar] [CrossRef]
- Dalar, A.; Konczak, I. Phenolic contents, antioxidant capacities and inhibitory activities against key metabolic syndrome relevant enzymes of herbal teas from Eastern Anatolia. Ind. Crops Prod. 2013, 44, 383–390. [Google Scholar] [CrossRef]
- Sinija, V.R.; Mishra, H.N. Green tea: health benefits. J. Nutr. Environ. Med. 2008, 17, 232–242. [Google Scholar] [CrossRef]
- Wang, H.; Du, Y.-J.; Song, H.-C. α-Glucosidase and α-amylase inhibitory activities of guava leaves. Food Chem. 2010, 123, 6–13. [Google Scholar] [CrossRef]
- Sakulnarmrat, K.; Konczak, I. Composition of native Australian herbs polyphenolic-rich fractions and in vitro inhibitory activities against key enzymes relevant to metabolic syndrome. Food Chem. 2012, 134, 1011–1019. [Google Scholar] [CrossRef] [PubMed]
- Worsztynowicz, P.; Napierała, M.; Białas, W.; Grajek, W. Pancreatic α-amylase and lipase inhibitory activity of polyphenolic compounds present in the extract of black chokeberry (Aronia melanocarpa L.). Process Biochem. 2014, 49, 1457–1463. [Google Scholar] [CrossRef]
- Podsedek, A.; Majewska, I.; Malgorzata, R.; Sosnowska, D.; Koziolkiewicz, M. In Vitro Inhibitory Effect on Digestive Enzymes and Antioxidant Potential of Commonly Consumed Fruits. J. Agric. Food Chem. 2014, 62, 4610–4617. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Wang, W.; Zhu, T.; Liang, T.; Lu, F.; He, W.; Zhang, H.; Liu, Z.; He, S.; Gao, K.; et al. Phenylpropanoid glycoside inhibition of pepsin, trypsin and α-chymotrypsin enzyme activity in Kudingcha leaves from Ligustrum purpurascens. Food Res. Int. 2013, 54, 1376–1382. [Google Scholar] [CrossRef]
- Li, Q.; Wei, Q.; Yuan, E.; Yang, J.; Ning, Z. Interaction between four flavonoids and trypsin: effect on the characteristics of trypsin and antioxidant activity of flavonoids. Int. J. Food Sci. Technol. 2014, 49, 1063–1069. [Google Scholar] [CrossRef]
- Zeng, H.-j.; Yang, R.; Liang, H.; Qu, L.-B. Molecular interactions of flavonoids to pepsin: Insights from spectroscopic and molecular docking studies. Spectrochim. Acta A Mol. Biomol. Spectros. 2015, 151, 576–590. [Google Scholar] [CrossRef] [PubMed]
- You, Q.; Chen, F.; Wang, X.; Jiang, Y.; Lin, S. Anti-diabetic activities of phenolic compounds in muscadine against alpha-glucosidase and pancreatic lipase. LWT-Food Sci. Technol. 2012, 46, 164–168. [Google Scholar] [CrossRef]
- Cai, X.; Yu, J.; Xu, L.; Liu, R.; Yang, J. The mechanism study in the interactions of sorghum procyanidins trimer with porcine pancreatic α-amylase. Food Chem. 2015, 174, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Narita, Y.; Inouye, K. Inhibitory effects of chlorogenic acids from green coffee beans and cinnamate derivatives on the activity of porcine pancreas α-amylase isozyme I. Food Chem. 2011, 127, 1532–1539. [Google Scholar] [CrossRef]
- Muthamma Milan, K.S.; Dholakia, H.; Tiku, P.K.; Vishveshwaraiah, P. Enhancement of digestive enzymatic activity by cumin (Cuminum cyminum L.) and role of spent cumin as a bionutrient. Food Chem. 2008, 110, 678–683. [Google Scholar] [CrossRef]
- Sosa, N.; Tibisay, P.; Araujo, L. Determinación del Efecto Inhibitorio de los Polifenoles presentes en la Fresa (Fragaria vesca L.) sobre la Enzima Alfa Amilasa. Rev. Fac. Farm. 2002, 43, 47–49. [Google Scholar]
- Hemalatha, P.; Bomzan, D.; Rao, B.V.; Sreerama, Y.N. Distribution of phenolic antioxidants in whole and milled fractions of quinoa and their inhibitory effects on α-amylase and α-glucosidase activities. Food Chem. 2016, 199, 330–338. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo-González, M.; Grosso, C.; Valentão, P.; Andrade, P.B. α-Glucosidase and α-amylase inhibitors from Myrcia spp.: A stronger alternative to acarbose? J. Pharm. Biomed. Anal. 2016, 118, 322–327. [Google Scholar] [CrossRef] [PubMed]
- Ademiluyi, A.O.; Oboh, G. Soybean phenolic-rich extracts inhibit key-enzymes linked to type 2 diabetes (α-amylase and α-glucosidase) and hypertension (angiotensin I converting enzyme) in vitro. Exp. Toxicol. Pathol. 2013, 65, 305–309. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Li, D.; Huang, B.; Chen, Y.; Lu, X.; Wang, Y. Inhibition of pancreatic lipase, α-glucosidase, α-amylase, and hypolipidemic effects of the total flavonoids from Nelumbo nucifera leaves. J. Ethnopharmacol. 2013, 149, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Nwosu, F.; Morris, J.; Lund, V.A.; Stewart, D.; Ross, H.A.; McDougall, G.J. Anti-proliferative and potential anti-diabetic effects of phenolic-rich extracts from edible marine algae. Food Chem. 2011, 126, 1006–1012. [Google Scholar] [CrossRef]
- Shobana, S.; Sreerama, Y.N.; Malleshi, N.G. Composition and enzyme inhibitory properties of finger millet (Eleusine coracana L.) seed coat phenolics: Mode of inhibition of α-glucosidase and pancreatic amylase. Food Chem. 2009, 115, 1268–1273. [Google Scholar] [CrossRef]
- Tadera, K.; Minami, Y.; Takamatsu, K. Inhibition of ALPHA-Glucosidase and ALPHA-Amylase by Flavonoids. J. Nutr. Sci. Vitaminol. 2006, 52, 149–153. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Chang, S.K.C.; Zhang, Y. Comparison of α-amylase, α-glucosidase and lipase inhibitory activity of the phenolic substances in two black legumes of different genera. Food Chem. 2017, 214, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Moreno, D.A.; Ilic, N.; Poulev, A.; Raskin, I. Effects of Arachis hypogaea nutshell extract on lipid metabolic enzymes and obesity parameters. Life Sci. 2005, 78, 2797–2803. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Cui, F.; Yin, F.; Zeng, X.; Sun, Y.; Li, Y. Caffeoylquinic acids competitively inhibit pancreatic lipase through binding to the catalytic triad. Int. J. Biol. Macromol. 2015, 80, 529–535. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Dong, S.; Zhang, R.; Shao, H.; Liu, Y. Effects of proanthocyanidins on porcine pancreatic lipase: Conformation, activity, kinetics and thermodynamics. Process Biochem. 2014, 49, 237–243. [Google Scholar] [CrossRef]
- Li, Y.-Q.; Yang, P.; Fei, G.; Zhang, Z.-W.; Wu, B. Probing the interaction between 3 flavonoids and pancreatic lipase by methods of fluorescence spectroscopy and enzymatic kinetics. Eur. Food Res. Technol. 2011, 233, 63–69. [Google Scholar] [CrossRef]
- Uchiyama, S.; Taniguchi, Y.; Saka, A.; Yoshida, A.; Yajima, H. Prevention of diet-induced obesity by dietary black tea polyphenols extract in vitro and in vivo. Nutrition 2011, 27, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Birari, R.B.; Gupta, S.; Gopi Mohan, C.; Bhutani, K.K. Antiobesity and lipid lowering effects of Glycyrrhiza chalcones: Experimental and computational studies. Phytomedicine 2011, 18, 795–801. [Google Scholar] [CrossRef] [PubMed]
- Gondoin, A.; Grussu, D.; Stewart, D.; McDougall, G.J. White and green tea polyphenols inhibit pancreatic lipase in vitro. Food Res. Int. 2010, 43, 1537–1544. [Google Scholar] [CrossRef]
- Sharma, N.; Sharma, V.K.; Seo, S.-Y. Screening of some medicinal plants for anti-lipase activity. J. Ethnopharmacol. 2005, 97, 453–456. [Google Scholar] [CrossRef] [PubMed]
- Lai, P.; Okazawa, A.; Izumi, Y.; Bamba, T.; Fukusaki, E.; Yoshikawa, M.; Kobayashi, A. Effect of gallic acid on peptides released by trypsin digestion of bovine α-casein. J. Biosci. Bioeng. 2013, 115, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, R.; Mateus, N.; Pianet, I.; Laguerre, M.; Freitas, V.D. Mechanisms of Tannin-Induced Trypsin Inhibition: A Molecular Approach. Langmuir 2011, 27, 13122–13129. [Google Scholar] [CrossRef] [PubMed]
- Akkarachiyasit, S.; Yibchok-Anun, S.; Wacharasindhu, S.; Adisakwattana, S. In Vitro Inhibitory Effects of Cyandin-3-rutinoside on Pancreatic α-Amylase and Its Combined Effect with Acarbose. Molecules 2011, 16, 2075–2083. [Google Scholar] [CrossRef] [PubMed]
- Kanakis, C.D.; Hasni, I.; Bourassa, P.; Tarantilis, P.A.; Polissiou, M.G.; Tajmir-Riahi, H.-A. Milk β-lactoglobulin complexes with tea polyphenols. Food Chem. 2011, 127, 1046–1055. [Google Scholar] [CrossRef] [PubMed]
- Jia, J.; Gao, X.; Hao, M.; Tang, L. Comparison of binding interaction between β-lactoglobulin and three common polyphenols using multi-spectroscopy and modeling methods. Food Chem. 2017, 228, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Vazquez-Flores, A.A.; Wong-Paz, J.E.; Lerma-Herrera, M.A.; Martinez-Gonzalez, A.I.; Olivas-Aguirre, F.J.; Aguilar, C.N.; Wall-Medrano, A.; Gonzalez-Aguilar, G.A.; Alvarez-Parrilla, E.; de la Rosa, L.A. Proanthocyanidins from the kernel and shell of pecan (Carya illinoinensis): Average degree of polymerization and effects on carbohydrate, lipid, and peptide hydrolysis in a simulated human digestive system. J. Funct. Foods 2017, 28, 227–234. [Google Scholar] [CrossRef]
- Wu, X.; Wu, H.; Liu, M.; Liu, Z.; Xu, H.; Lai, F. Analysis of binding interaction between (−)-epigallocatechin (EGC) and β-lactoglobulin by multi-spectroscopic method. Spectrochim. Acta A Mol. Biomol. Spectros. 2011, 82, 164–168. [Google Scholar] [CrossRef] [PubMed]
- Prigent, S.V.E.; Gruppen, H.; Visser, A.; van Koningsveld, G.A.; de Jong, G.A.H.; Voragen, A.G.J. Effects of non-covalent interactions with 5-O-caffeoylquinic acid (chlorogenic acid) on the heat denaturation and solubility of globular proteins. J. Agric. Food Chem. 2003, 51, 5088–5095. [Google Scholar] [CrossRef] [PubMed]
- Yuksel, Z.; Avci, E.; Erdem, Y.K. Characterization of binding interactions between green tea flavanoids and milk proteins. Food Chem. 2010, 121, 450–456. [Google Scholar] [CrossRef]
- Helal, A.; Tagliazucchi, D.; Verzelloni, E.; Conte, A. Bioaccessibility of polyphenols and cinnamaldehyde in cinnamon beverages subjected to in vitro gastro-pancreatic digestion. J. Funct. Foods 2014, 7, 506–516. [Google Scholar] [CrossRef]
- Lakowicz, J.R. Quenching of Fluorescence. In Principles of Fluorescence Spectroscopy, 2nd ed.; Kluwer Academic/Plenum Publishers: New York, NY, USA, 1999; pp. 237–265. [Google Scholar]
- Gonçalves, R.; Mateus, N.; Freitas, D.V. Inhibition of α-amylase activity by condensed tannins. Food Chem. 2011, 125, 665–672. [Google Scholar] [CrossRef]
- Skrt, M.; Benedik, E.; Podlipnik, C.; Ulrih, N.P. Interactions of different polyphenols with bovine serum albumin using fluorescence quenching and molecular docking. Food Chem. 2012, 135, 2418–2424. [Google Scholar] [CrossRef] [PubMed]
- Bourvellec, C.L.; Renard, C.M.G.C. Interactions between Polyphenols and Macromolecules: Quantification Methods and Mechanisms. Crit. Rev. Food Sci. Nutr. 2012, 52, 213–248. [Google Scholar] [CrossRef] [PubMed]
- Ross, P.D.; Subramanian, S. Thermodynamics of protein association reactions: Forces contributing to stability. Biochemistry 1981, 20, 3096–3102. [Google Scholar] [CrossRef] [PubMed]
- Trivella, D.B.B.; dos Reis, C.V.; Lima, L.M.T.R.; Foguel, D.; Polikarpov, I. Flavonoid interactions with human transthyretin: Combined structural and thermodynamic analysis. J. Struct. Biol. 2012, 180, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Budryn, G.; Pałecz, B.; Rachwał-Rosiak, D.; Oracz, J. Effect of inclusion of hydroxycinnamic and chlorogenic acids from green coffee bean in β-cyclodextrin on their interactions with whey, egg white and soy protein isolates. Food Chem. 2015, 168, 276–287. [Google Scholar] [CrossRef] [PubMed]
- Kokotos, G. Inhibition of digestive lipases by 2-oxo amide triacylglycerol analogues. J. Mol. Catal. B Enzym. 2003, 22, 255–269. [Google Scholar] [CrossRef]
- Housaindokht, M.; Bozorgmehr, M.; Hosseini, H.; Jalal, R.; Asoodeh, A.; Saberi, M.; Haratipour, Z.; Monhemi, H. Structural properties of the truncated and wild types of Taka-amylase: A molecular dynamics simulation and docking study. J. Mol. Catal. B Enzym. 2013, 95, 36–40. [Google Scholar] [CrossRef]
- Giang, T.M.; Gaucel, S.; Brestaz, P.; Anton, M.; Meynier, A.; Trelea, I.C.; Le Feunteun, S. Dynamic modeling of in vitro lipid digestion: Individual fatty acid release and bioaccessibility kinetics. Food Chem. 2016, 194, 1180–1188. [Google Scholar] [CrossRef] [PubMed]
- Thongkaew, C.; Gibis, M.; Hinrichs, J.; Weiss, J. Polyphenol interactions with whey protein isolate and whey protein isolate–pectin coacervates. Food Hydrocoll. 2014, 41, 103–112. [Google Scholar] [CrossRef]
- Hur, S.J.; Lim, B.O.; Decker, E.A.; McClements, D.J. In vitro human digestion models for food applications. Food Chem. 2011, 125, 1–12. [Google Scholar] [CrossRef]
- Unajak, S.; Meesawat, P.; Paemanee, A.; Areechon, N.; Engkagul, A.; Kovitvadhi, U.; Kovitvadhi, S.; Rungruangsak-Torrissen, K.; Choowongkomon, K. Characterisation of thermostable trypsin and determination of trypsin isozymes from intestine of Nile tilapia (Oreochromis niloticus L.). Food Chem. 2012, 134, 1533–1541. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme | PCs Extract | PCs Identified on the Extracts by HPLC | PCs Standard | Assessment Methods | Note | Reference | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| UVV | FLU | ITC | DSC | CD | MOD | Others | |||||||||
| % Inh | IC50 | KIN | AFF | THR | |||||||||||
| AG | ND | ND | Tannic acid | * | * | * | * | * | KIN calculated parameters (KM and Vmax); THR calculated parameters (ΔG, ΔH and ΔS); and AFF calculated parameters: Ksv, kq, Ka, n. | [29] | |||||
| Twenty Canadian lentil cultivars (Lens culinaris) | Twenty one mainly: p-hydroxybenzoic acid, syringic acid, epicatechin gallate, quercetin-3-xyloside, quercetin-3-glucoside, kaempferol-3-glucoside | Catechin, epicatechin, procyanidin B1, kaempferol, kaempferol-glucose, quercetin, quercetin-arabinose | * | [21] | |||||||||||
| Australian fruits (Davidson pruriens and Santalum acuminatum) | Ellagic acid, flavonoids (myricetin, quercetin, rutin), ellagitannins, and anthocyanins | ND | * | [27] | |||||||||||
| Muscadine (Vitis rotundifolia) | Twelve like catechin, ellagic acid, quercetin | ND | * | * | KIN including Ki. | [67] | |||||||||
| Tasmannian pepper (Drymis lanceolata) leaf, anise myrtle (Syzygium anisatum), lemon myrtle (Backhousia citriodora) | Ellagic acid, chlorogenic acid, flavonoids (catechin, myricetin, hesperetin, quercetin) | ND | * | * | [61] | ||||||||||
| PA | Green, black and oolong tea (Camellia sinensis) | ND | ND | * | * | [25] | |||||||||
| Sorghum | Procyanidins | ND | * | * | AFF (Ksv, kq, Ka). | [68] | |||||||||
| Pinhão coat (Araucaria angustifolia and A. mearnsii) | Condensed tannin | ND | * | * | KIN parameters; Measurement of post-prandial glycemic levels in healthy rats. | [12] | |||||||||
| Black chokeberry (Aronia melanocarpa L.) | Phenolic acids as chlorogenic acid, and anthocyanins as cyanidin-3-glucoside | ND | * | [62] | |||||||||||
| Green coffee (Coffea arabica) beans | Eight chlorogenic acids derivatives: three subgroups of caffeoylquinic acids, feruloylquinic acids, and dicaffeoylquinic acids | ND | * | * | KIN parameters. | [69] | |||||||||
| Cumin (Cuminum cyminum L.) | ND | ND | * | * | [70] | ||||||||||
| Nepalese medicinal herb Pakhanbhed (Bergenia ciliata) | (-)-3-o-galloylepicatechin and (-)-3-o-galloylcatechin | ND | * | [37] | |||||||||||
| Chinese green tea | Catechin | ND | * | * | * | KIN parameters. | [28] | ||||||||
| Strawberry (Fragaria vesca L.) | PCs fractions | ND | [71] | ||||||||||||
| AG, PA | Quinoa (Chenopodium quinoa) | Phenolic, flavonoid, and condensed tannins contents | ND | * | * | [72] | |||||||||
| Five species of Myrcia genus | Gallic acid, flavan-3-ols, and flavonols | ND | * | * | [73] | ||||||||||
| Chañar (Geoffrea decorticans) fruit | Caffeic acid, protocatechuic acid, vanillic acid, p-coumaric acid among others | Quercetin | * | [22] | |||||||||||
| Soybean (Glycine max) | Bound and free phenolic extract | ND | * | [74] | |||||||||||
| Nelumbo nucifera leaves | Flavonoids | ND | * | * | Measurement of total cholesterol, triacylglyceride and low-density lipoprotein cholesterol contents in high fat diet-fed rats, and others. | [75] | |||||||||
| Six herbal teas | From Plantago lanceolata, L.: chlorogenic acid, rutin, among others | Gallic acid, catechin, among others | * | * | [58] | ||||||||||
| Algae (Palmaria, Ascophyllum and Alaria) | Tannins | ND | * | [76] | |||||||||||
| Guava (Psidium guajava L.) leaves | Quercetin, kaempferol, guaijaverin, avicularin, myricetin, hyperin, and apigenin. | ND | * | * | [60] | ||||||||||
| Finger millet (Eleusine coracana L.) seed | Gallic acid, caffeic acid, kaempferol, among others. | ND | * | * | KIN parameters. | [77] | |||||||||
| ND | ND | Flavonoids | * | * | [78] | ||||||||||
| Black soybean (Glycine max) and black turtle beans (Phaseolus vulgaris) | ND | Phenolic acids (gallic acid, syringic acid and others), and flavonoids (catechin, quercetin-3-o-glucoside) | * | * | [79] | ||||||||||
| PL | Peanut (Arachis hypogaea L.) | ND | ND | * | * | Measurement of body weight, liver size, fecal lipid excretion and triacylglyceride content in high fat diet-fed rats. | [80] | ||||||||
| Chañar fruit | Caffeic acid, protocatechuic acid, vanillic acid, p-coumaric acid among others. | Quercetin | * | [22] | |||||||||||
| Twenty Canadian lentil cultivars | Twenty one as: p-hydroxybenzoic acid, and quercetin-3-o-glucoside. | Quercetin and quercetin-arabinoside | * | [21] | |||||||||||
| ND | ND | Cinnamic acid | * | * | Measurement of body weight, total cholesterol and triacylglyceride contents in high fat diet-fed rats, and others. | [20] | |||||||||
| ND | ND | 3-caffeoylquinic acid (CQA), 4,5-CQA, 3,4-CQA, 3,5-CQA, and 4,5-diCQA | * | * | [81] | ||||||||||
| Twenty eight traditional Thai medicinal herbs | ND | ND | * | * | [57] | ||||||||||
| Horseradish (Armoracia rusticana) | ND | ND | * | [6] | |||||||||||
| ND | ND | Acteoside | * | * | * | * | * | [11] | |||||||
| ND | ND | Proanthocyani-dins | * | * | * | * | * | * | KIN parameters; AFF parameters: Ksv, kq, Ka, fa; and formation of protein aggregates. Also a hydrodynamic radius analysis was performed. | [82] | |||||
| Black chokeberry | Phenolic acids and anthocyanins | ND | * | [62] | |||||||||||
| Australian fruits | Ellagic acid, ellagitannins, flavonoids and anthocyanins | ND | * | * | [27] | ||||||||||
| Nelumbo nucifera leaves | Flavonoids | ND | * | Measurement of lipid components such as triacylglyceride, total cholesterol, and others in high fat diet-fed rats. | [75] | ||||||||||
| Six herbal teas | From Plantago lanceolata, L.: chlorogenic acid, rutin, among others | Gallic acid, catechin, among others. | * | * | [58] | ||||||||||
| Muscadine | Twelve like catechin, ellagic acid, and quercetin | ND | * | * | KIN parameters and Ki. | [67] | |||||||||
| Green tea and grape seeds | ND | Epigallocatechin-3-gallate, kaempferol, and quercetin | * | * | [34] | ||||||||||
| Tasmannian pepper leaf, anise myrtle, lemon myrtle | Ellagic acid, chlorogenic acid, and flavonoids (i.e., catechin) | ND | * | * | [61] | ||||||||||
| Tartary buckwheat bran | ND | Quercetin, isoquercetin and rutin | * | * | * | * | [83] | ||||||||
| Black tea | Polymerized polyphenol fraction | Polymerized catechins such as theaflavin and theaflavin-3-gallate | * | * | Measurement of triacylglyceride content and body weight in high fat diet-fed rats. | [84] | |||||||||
| Root of Glycyrrhiza glabra | Twelve flavonoids | ND | * | * | * | Measurement of body weight, total cholesterol and triacylglyceride contents in high fat diet-fed rats. | [85] | ||||||||
| White and green tea | Flavan-3-ols for green tea, and 5-galloyl quinic acid, digalloyl glucose, trigalloyl glucose and strictinin for white tea | ND | * | * | [86] | ||||||||||
| Berries (blackcurrantrowan, blueberry, lingonberry, among others) | Tannins: ellagitannin and proanthocyanidin | ND | * | * | [18] | ||||||||||
| Cumin | ND | ND | * | [70] | |||||||||||
| Chinese green tea | Catechin | ND | * | [28] | |||||||||||
| Peanut | ND | ND | * | * | Measurement of body weight, liver size, fecal lipid excretion and triacylglyceride content in high fat diet-fed rats. | [80] | |||||||||
| Seventy five medicinal plants | ND | ND | * | * | A radioactive method was used. | [87] | |||||||||
| Black soybean (Glycine max) and black turtle beans (Phaseolus vulgaris) | Phenolic acids (gallic acid, syringic acid and others), and flavonoids (catechin, quercetin-3-o-glucoside) | * | * | [79] | |||||||||||
| PE | ND | ND | Ten flavonoids (principally baicalein) | * | * | * | * | AFF pameters: Ksv, kq. | [66] | ||||||
| TP | ND | ND | Tannic acid | * | * | * | * | AFF parameters: Ksv, kq, Ka, n. | [29] | ||||||
| ND | ND | Flavonoids as quercetin, luteolin, and kaempferol | * | * | AFF parameters: Ksv, kq. | [65] | |||||||||
| ND | ND | Gallic acid | * | [88] | |||||||||||
| ND | ND | Procyanidins (catechin) | * | * | * | * | Dynamic light scattering and nephelometry were used too. | [89] | |||||||
| PE, TP | Chinese green tea | Catechin | ND | * | [28] | ||||||||||
| PE, TP, CT | Tea (Ligustrum purpurascens) | Phenylpropanoid glycosides (like acteoside) | ND | * | * | * | * | * | AFF parameters: Ka and n. | [64] | |||||
| Enzyme | PCs | Inhibition Type | Type of Binding or Force | Binding Site in the Enzyme | Reference |
|---|---|---|---|---|---|
| AG | Tannic acid | Mixed-type | Hydrophobic and electrostatic | ND | [29] |
| Twelve PCs such as catechin, ellagic acid, and quercetin | Competitive | ND | ND | [67] | |
| PA | Teas (green, black and oolong tea) | Non-competitive | ND | ND | [25] |
| Procyanidins | ND | Hydrophobic | One site | [68] | |
| Condensed tannins | Mixed-type | ND | ND | [12] | |
| Eight Chlorogenic acids: three subgroups of caffeoylquinic acids, feruloylquinic acids, and dicaffeoylquinic acids | Mixed-type | Hydrogen | Non-catalytic sites | [69] | |
| Catechin | ND | Hydrogen and hydrophobic | ND | [28] | |
| AG, PA | Gallic acid, caffeic acid, kaempferol, and others | Non-competitive | ND | ND | [77] |
| PL | 3-caffeoylquinic acid (CQA) derivatives as 4,5-CQA, 3,4-CQA, 3,5-CQA, and 4,5-diCQA | Competitive | Hydrogen and hydrophobic | Catalytic triad | [81] |
| Acteoside | Non-competitive | Hydrogen | Non-catalytic sites | [11] | |
| Proanthocyanidins | Non-competitive | Weak | ND | [82] | |
| 12 like catechin, ellagic acid, as quercetin | Competitive | ND | ND | [67] | |
| Quercetin, isoquercetin and rutin | Non-competitive | Hydrophobic and van der Waals | One site | [83] | |
| 12 flavonoids such as isoliquiritigenin | Competitive | Hydrogen | Catalytic site | [85] | |
| Catechin | ND | Hydrogen and hydrophobic | ND | [28] | |
| PE | 10 Flavonoids (principally baicalein) | ND | Hydrophobic and electrostatic | One hydrophobic site or cavity | [66] |
| TP | Tannic acid | Mixed-type | Hydrophobic and electrostatic | ND | [29] |
| Gallic acid | ND | Hydrophobic | ND | [88] | |
| Procyanidins (catechin) | Competitive | Hydrogen bonds | Near to catalytic site | [99] | |
| PE, TP | Catechin | ND | Hydrogen and hydrophobic | ND | [28] |
| PE, TP, CT | Phenylpropanoid glycosides like acteoside | Non-competitive | Hydrogen, hydrophobic, van der Waals and electrostatic | Catalytic sites | [64] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martinez-Gonzalez, A.I.; Díaz-Sánchez, Á.G.; Rosa, L.A.d.l.; Vargas-Requena, C.L.; Bustos-Jaimes, I.; Alvarez-Parrilla, A.E. Polyphenolic Compounds and Digestive Enzymes: In Vitro Non-Covalent Interactions. Molecules 2017, 22, 669. https://doi.org/10.3390/molecules22040669
Martinez-Gonzalez AI, Díaz-Sánchez ÁG, Rosa LAdl, Vargas-Requena CL, Bustos-Jaimes I, Alvarez-Parrilla AE. Polyphenolic Compounds and Digestive Enzymes: In Vitro Non-Covalent Interactions. Molecules. 2017; 22(4):669. https://doi.org/10.3390/molecules22040669
Chicago/Turabian StyleMartinez-Gonzalez, Alejandra I., Ángel G. Díaz-Sánchez, Laura A. de la Rosa, Claudia L. Vargas-Requena, Ismael Bustos-Jaimes, and And Emilio Alvarez-Parrilla. 2017. "Polyphenolic Compounds and Digestive Enzymes: In Vitro Non-Covalent Interactions" Molecules 22, no. 4: 669. https://doi.org/10.3390/molecules22040669
APA StyleMartinez-Gonzalez, A. I., Díaz-Sánchez, Á. G., Rosa, L. A. d. l., Vargas-Requena, C. L., Bustos-Jaimes, I., & Alvarez-Parrilla, A. E. (2017). Polyphenolic Compounds and Digestive Enzymes: In Vitro Non-Covalent Interactions. Molecules, 22(4), 669. https://doi.org/10.3390/molecules22040669