
Sesquiterpene Variation in West Australian Sandalwood (Santalum spicatum)

Abstract
:1. Introduction
2. Results and Discussion
2.1. Variability of Total Sesquiterpene Content
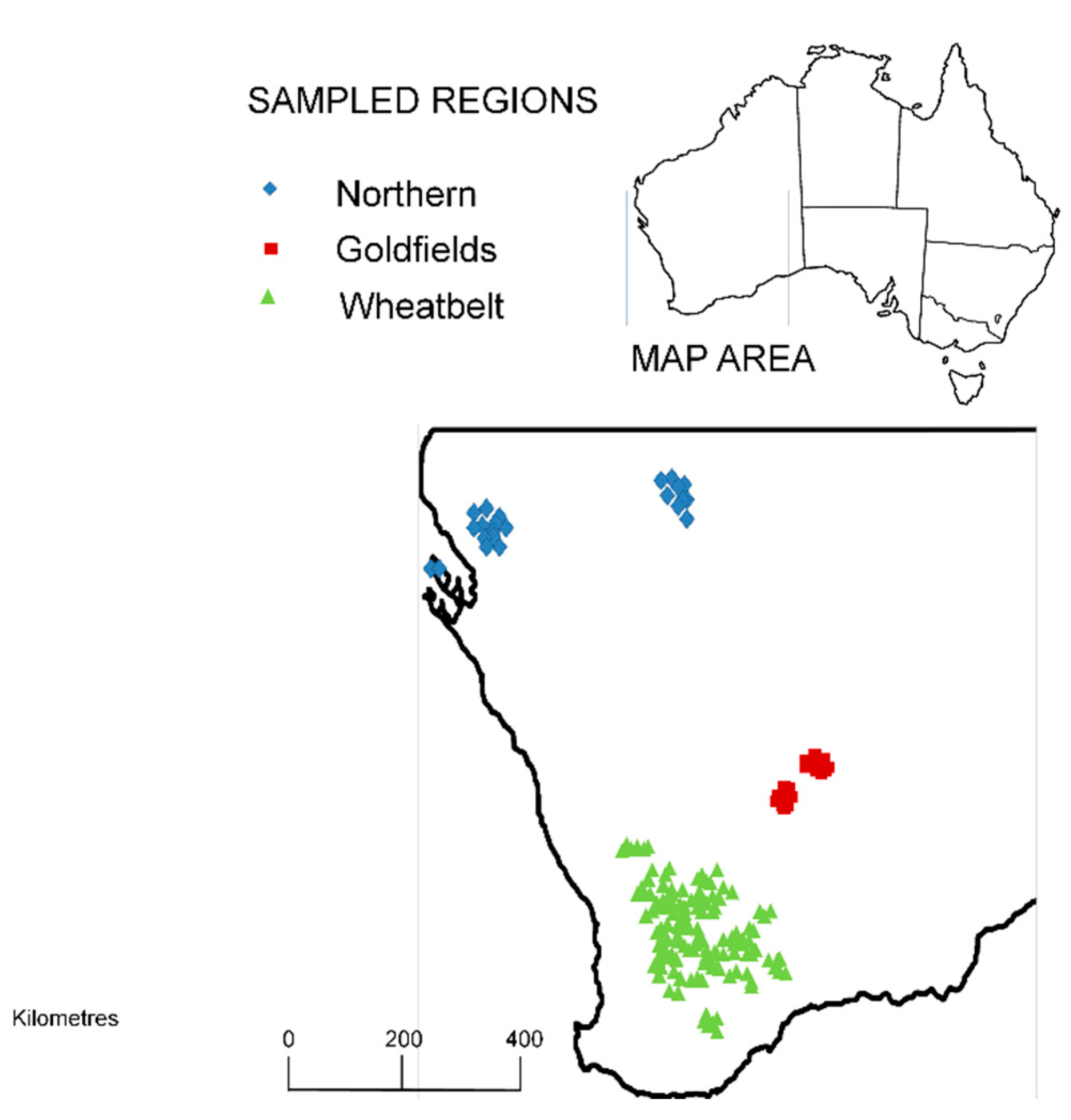
2.2. Sesquiterpene Profiles and Geographic Variation
2.3. Variation of Other Sesquiterpene Components
2.4. Statistical Analysis of Chemotypes
3. Materials and Methods
3.1. Substrates and Reagents
3.2. Plant Material Collection
3.3. S. spicatum Heartwood Extraction and GC-FID and GC-MS
3.4. Statistical Analysis
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Jones, C.G.; Ghisalberti, E.L.; Plummer, J.A.; Barbour, E.L. Quantitative co-occurrence of sesquiterpenes; a tool for elucidating their biosynthesis in Indian sandalwood, Santalum album. Phytochemistry 2006, 67, 2463–2468. [Google Scholar] [CrossRef] [PubMed]
- Moretta, P. Extraction and variation of the essential oil from Western Australian sandalwood (Santalum spicatum). Ph.D. Thesis, University of Western Australia, Perth, Australia, 2001. [Google Scholar]
- Page, T.; Southwell, I.; Russell, M.; Tate, H.; Tungon, J.; Sam, C.; Dickinson, G.; Robson, K.; Leakey, R.R.B. Geographic and phenotypic variation in heartwood and essential-oil characters in natural populations of Santalum austrocaledonicum in Vanuatu. Chem. Biodivers. 2010, 7, 1990–2006. [Google Scholar] [CrossRef] [PubMed]
- Howes, M.-J.R.; Simmonds, M.S.J.; Kite, G.C. Evaluation of the quality of sandalwood essential oils by gas chromatography-mass spectrometry. J. Chromatog. A 2004, 1028, 307–312. [Google Scholar] [CrossRef]
- The International Organisation for Standardisation. International Standard ISO 3518:2002 Oil of sandalwood (Santalum album L.). Available online: https://www.iso.org/standard/32037.html (accessed on 16 January 2013).
- Brand, J.E.; Pronk, G.M. Influence of age on sandalwood (Santalum spicatum) oil content within different wood grades from five plantations in Western Australia. Aust. For. 2011, 74, 141–148. [Google Scholar] [CrossRef]
- Hostynek, J.J.; Magee, P.S. Fragrance allergens: Classification and ranking by QSAR. Toxicol. In Vitro 1997, 11, 377–384. [Google Scholar] [CrossRef]
- Schnuch, A.; Uter, W.; Geier, J.; Lessmann, H.; Frosch, P.J. Contact allergy to farnesol in 2021 consecutively patch tested patients. Results of the IVDK. Contact Dermat. 2004, 50, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Adams, D.R.; Bhatnagar, S.P.; Cookson, R.C. Sesquiterpenes of Santalum album and Santalum spicatum. Phytochemistry 1975, 14, 1459–1460. [Google Scholar] [CrossRef]
- Baldovini, N.; Delasalle, C.; Joulain, D. Phytochemistry of the heartwood from fragrant Santalum species: A review. Flavour Frag. J. 2011, 26, 7–26. [Google Scholar] [CrossRef]
- Brophy, J.J.; Fookes, C.J.R.; Lassak, E.V. Constituents of Santalum spicatum (R.Br.) A. DC. wood oil. J. Essent. Oil Res. 1991, 3, 381–385. [Google Scholar] [CrossRef]
- Valder, C.; Neugebauer, M.; Meier, M.; Kohlenberg, B.; Hammerschmidt, F.-J.; Braun, N.A. Western Australian sandalwood oil—New constituents of Santalum spicatum (R.Br.) A. DC. (Santalaceae). J. Essent. Oil Res. 2003, 15, 178–186. [Google Scholar] [CrossRef]
- Jones, C.G.; Keeling, C.I.; Ghisalberti, E.L.; Barbour, E.L.; Plummer, J.A.; Bohlmann, J. Isolation of cDNAs and functional characterisation of two multi-product terpene synthase enzymes from sandalwood, Santalum album L. Arch. Biochem. Biophys. 2008, 477, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Byrne, M.; MacDonald, B.; Brand, J.E. Regional genetic differentiation in Western Australian sandalwood (Santalum spicatum) as revealed by nuclear RFLP analysis. Theor. Appl. Genet. 2003, 107, 1208–1214. [Google Scholar] [CrossRef] [PubMed]
- Byrne, M.; MacDonald, B.; Brand, J.E. Phylogeography and divergence in the chloroplast genome of Western Australian sandalwood (Santalum spicatum). Heredity 2003, 91, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Applegate, G.B.; Chamberlain, J.; Daruhi, G.; Feigelson, J.L.; Hamilton, L.; McKinnell, F.H.; Neil, P.E.; Rai, S.N.; Rodehn, P.; Statham, P.; et al. A state of knowledge synthesis and summary from the April 1990 symposium. In Proceedings of the Symposium on Sandalwood in the Pacific, Honolulu, HI, USA, 9–11 April 1990; Hamilton, L., Conrad, C.E., Eds.; US Forest Service: Honolulu, HI, USA, 1990; Volume PSW-122, pp. 1–11. [Google Scholar]
- Brand, J.E.; Fox, J.E.D.; Pronk, G.; Cornwell, C. Comparison of oil concentration and oil quality from Santalum spicatum and S. album plantations, 8–25 years old, with those from mature S. spicatum natural stands. Aust. For. 2007, 70, 235–241. [Google Scholar] [CrossRef]
- Egerton-Warburton, L.M.; Ghisalberti, E.L.; Considine, J.A. Infraspecific variability in the volatile leaf oils of Chamelaucium uncinatum (Myrtaceae). Biochem. Syst. Ecol. 1998, 26, 873–888. [Google Scholar] [CrossRef]
- Byrne, M.; Yeates, D.K.; Joseph, L.; Kearney, M.; Bowler, J.; Williams, M.A.J.; Cooper, S.; Donnellan, S.C.; Keogh, J.S.; Leys, R.; et al. Birth of a biome: Insights into the assembly and maintenance of the Australian arid zone biota. Mol. Ecol. 2008, 17, 4398–4417. [Google Scholar] [CrossRef] [PubMed]
- Loneragan, O.W. Historical Review of Sandalwood (Santalum spicatum) Research in Western Australia Management; Research Bulletin No.4.; Department of Conservation and Land Management: Perth, Australia, 1990; pp. 1–53.
- Steffen, W.; Burbidge, A.A.; Hughes, L.; Kitching, R.; Lindenmayer, D.; Musgrave, W.; Smith, M.S.; Werner, P.A. Australia’s Biodiversity and Climate Change; CSIRO Publishing: Clayton, Australia, 2009. [Google Scholar]
- Verghese, J.; Sunny, T.P.; Balakrishnan, K.V. (Z)-(+)-α-santalol and (Z)-(−)-β-santalol concentration, a new quality determinant of east Indian sandalwood oil. Flavour Frag. J. 1990, 5, 223–226. [Google Scholar] [CrossRef]
- Celedon, J.M.; Chiang, A.; Yuen, M.M.S.; Diaz-Chavez, M.L.; Madilao, L.L.; Finnegan, P.M.; Barbour, E.L.; Bohlmann, J. Heartwood specific transcriptome and metabolite signatures of tropical sandalwood (Santalum album) reveal the final step of (Z)-santalol fragrance biosynthesis. Plant J. 2016, 86, 289–299. [Google Scholar] [CrossRef] [PubMed]
- McComb, J.A.; Jones, M.G.K. Interspecific hybridisation between Santalum album and S. spicatum. In Sandal and its Products; Radomiljac, A., Ananthapadmanabho, H.S., Welbourn, R.M., Rao., K.S., Eds.; ACIAR: Canberra, Australia, 1998; Volume 84, pp. 36–41. [Google Scholar]
- Jones, C.G.; Moniodis, J.; Zulak, K.G.; Scaffidi, A.; Plummer, J.A.; Ghisalberti, E.L.; Barbour, E.L.; Bohlmann, J. Sandalwood fragrance biosynthesis involves sesquiterpene synthases of both the terpene synthase (TPS) a and TPS-b subfamilies, including santalene synthases. J. Biol. Chem. 2011, 286, 17445–17454. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Chavez, M.L.; Moniodis, J.; Madilao, L.L.; Jancsik, S.; Keeling, C.I.; Barbour, E.L.; Ghisalberti, E.L.; Plummer, J.A.; Jones, C.G.; Bohlmann, J. Biosynthesis of sandalwood Oil: Santalum album CYP76F cytochromes P450 produce santalols and bergamotol. PLoS ONE 2013, 8, e75053. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Tholl, D.; Bohlmann, J.; Pichersky, E. The family of terpene synthases in plants: A mid-size family of genes for specialized metabolism that is highly diversified throughout the kingdom. Plant J. 2011, 66, 212–229. [Google Scholar] [CrossRef] [PubMed]
- Hamberger, B.; Ohnishi, T.; Hamberger, B.; Séguin, A.; Bohlmann, J. Evolution of diterpene metabolism: Sitka spruce CYP720B4 catalyses multiple oxidations in resin acid biosynthesis of conifer defense against insects. Plant Physiol. Biochem. 2011, 157, 1677–1695. [Google Scholar] [CrossRef] [PubMed]
- Charles, D.J.; Simon, J.E.; Wood, K.V. Essential oil constituents of Ocimum micranthum Willd. J. Agric. Food Chem. 1990, 38, 120–122. [Google Scholar] [CrossRef]
- Vieira, R.F.; Grayer, R.J.; Paton, A.; Simon, J.E. Genetic diversity of Ocimum gratissimum L. based on volatile oil constituents, flavonoids and RAPD markers. Biochem. Syst. Ecol. 2001, 29, 287–304. [Google Scholar] [CrossRef]
- Clarke, K.R. Non-parametric multivariate analyses of changes in community structure. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar]
- Clarke, K.R.; Gorley, R.N. PRIMER v6: User Manual/Tutorial; PRIMER-E: Plymouth, UK, 2006; pp. 1–192. [Google Scholar]
- The R Project for Statistical Computing. Available online: URLhttp://www.r-project.org (accessed on 11 March 2017).
Sample Availability: Due to the limited amounts, samples of the plant materials, extracts and reference compounds are not available from the authors. Some of the reference compounds are commercially available. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| R.I. | Southwest of W.A. (Wheatbelt) | Southeast of W.A. (Goldfields) | North of W.A. (Carnarvon/Shark Bay) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Components | Average (%) | Min | Max | SD | Average (%) | Min | Max | SD | Average (%) | Min | Max | SD | |
| E,E-farnesol | 2361 | 14.4 | 2.0 | 46.2 | 8.5 | 19.4 | 9.3 | 36.5 | 5.6 | 9.1 | 2.1 | 20.1 | 5.4 |
| Total α- and β-Santalol | 12.5 | 0.9 | 54.3 | 10.9 | 10.3 | 0.8 | 23.5 | 7.0 | 33.0 | 8.7 | 55.6 | 11.6 | |
| Z-nuciferol, Z-ϒ-curcumen-12-ol | 2520 2525 | 11.2 | 3.1 | 23.3 | 4.6 | 8.9 | 3.6 | 15.4 | 3.0 | 5.9 | 0.7 | 19.0 | 4.7 |
| α-santalol | 2346 | 9.3 | 0.6 | 41.3 | 8.3 | 7.4 | 0.5 | 16.6 | 5.0 | 24.8 | 6.2 | 38.3 | 9.2 |
| Z-β-curcumen-12-ol | 2470 | 6.9 | 0.3 | 17.1 | 3.5 | 6.8 | 0.6 | 14.0 | 4.2 | 6.9 | 0.3 | 19.8 | 5.2 |
| β-santalol | 2429 | 3.2 | 0.3 | 13.0 | 2.7 | 2.9 | 0.3 | 7.0 | 2.0 | 8.2 | 2.5 | 13.4 | 2.8 |
| Z-α-trans-bergamotol | 2358 | 2.8 | 0.0 | 14.0 | 2.2 | 2.7 | 0.2 | 6.3 | 1.6 | 4.2 | 1.2 | 7.3 | 1.9 |
| α-bisabolol | 2220 | 1.8 | 0.0 | 18.4 | 3.3 | 9.7 | 2.2 | 19.0 | 4.9 | 1.5 | 0.1 | 6.0 | 1.7 |
| Z-lanceol | 2496 | 1.4 | 0.3 | 8.7 | 1.4 | 5.0 | 1.5 | 9.6 | 2.4 | 2.3 | 0.9 | 6.9 | 1.4 |
| dendrolasin | 1927 | 1.3 | 0.2 | 5.3 | 0.9 | 2.3 | 0.7 | 5.4 | 1.2 | 0.7 | 0.2 | 1.5 | 0.4 |
| epi-β-santalol | 2414 | 0.6 | 0.0 | 1.9 | 0.4 | 0.5 | 0.0 | 1.3 | 0.4 | 2.1 | 0.5 | 4.8 | 1.1 |
| α-santalene | 1560 | 0.5 | 0.0 | 2.1 | 0.3 | 0.2 | 0.0 | 0.5 | 0.1 | 1.2 | 0.5 | 2.3 | 0.5 |
| trans-nerolidol | 2033 | 0.4 | 0.0 | 1.7 | 0.3 | 0.6 | 0.2 | 1.5 | 0.4 | 0.7 | 0.1 | 2.2 | 0.6 |
| β-santalene | 1640 | 0.4 | 0.0 | 1.6 | 0.3 | 0.1 | 0.0 | 0.4 | 0.1 | 1.1 | 0.5 | 2.0 | 0.4 |
| E-β-farnesene | 1760 | 0.3 | 0.0 | 1.2 | 0.2 | 0.0 | 0.0 | 0.1 | 0.0 | 0.0 | 0.0 | 0.1 | 0.0 |
| epi-β-santalene | 1621 | 0.2 | 0.0 | 1.1 | 0.2 | 0.1 | 0.0 | 0.2 | 0.1 | 0.7 | 0.3 | 1.3 | 0.3 |
| α-trans-bergamotene | 1576 | 0.2 | 0.0 | 0.7 | 0.1 | 0.1 | 0.0 | 0.1 | 0.0 | 0.4 | 0.2 | 0.8 | 0.2 |
| Total | 54.8 | 24.6 | 81.9 | 10.3 | 66.7 | 51.2 | 76.0 | 5.7 | 69.9 | 56.7 | 78.5 | 6.3 | |
| Yield (%) | 3.22 | 0.01 | 6.41 | 1.08 | 2.40 | 0.58 | 4.61 | 0.94 | 3.58 | 1.48 | 5.79 | 1.15 | |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moniodis, J.; Jones, C.G.; Renton, M.; Plummer, J.A.; Barbour, E.L.; Ghisalberti, E.L.; Bohlmann, J. Sesquiterpene Variation in West Australian Sandalwood (Santalum spicatum). Molecules 2017, 22, 940. https://doi.org/10.3390/molecules22060940
Moniodis J, Jones CG, Renton M, Plummer JA, Barbour EL, Ghisalberti EL, Bohlmann J. Sesquiterpene Variation in West Australian Sandalwood (Santalum spicatum). Molecules. 2017; 22(6):940. https://doi.org/10.3390/molecules22060940
Chicago/Turabian StyleMoniodis, Jessie, Christopher G. Jones, Michael Renton, Julie A. Plummer, E. Liz Barbour, Emilio L. Ghisalberti, and Joerg Bohlmann. 2017. "Sesquiterpene Variation in West Australian Sandalwood (Santalum spicatum)" Molecules 22, no. 6: 940. https://doi.org/10.3390/molecules22060940
APA StyleMoniodis, J., Jones, C. G., Renton, M., Plummer, J. A., Barbour, E. L., Ghisalberti, E. L., & Bohlmann, J. (2017). Sesquiterpene Variation in West Australian Sandalwood (Santalum spicatum). Molecules, 22(6), 940. https://doi.org/10.3390/molecules22060940