
Isoprenoids Production from Lipid-Extracted Microalgal Biomass Residues Using Engineered E. coli

Abstract
:1. Introduction
2. Results and Discussion
2.1. Chemical Composition of LMBRs
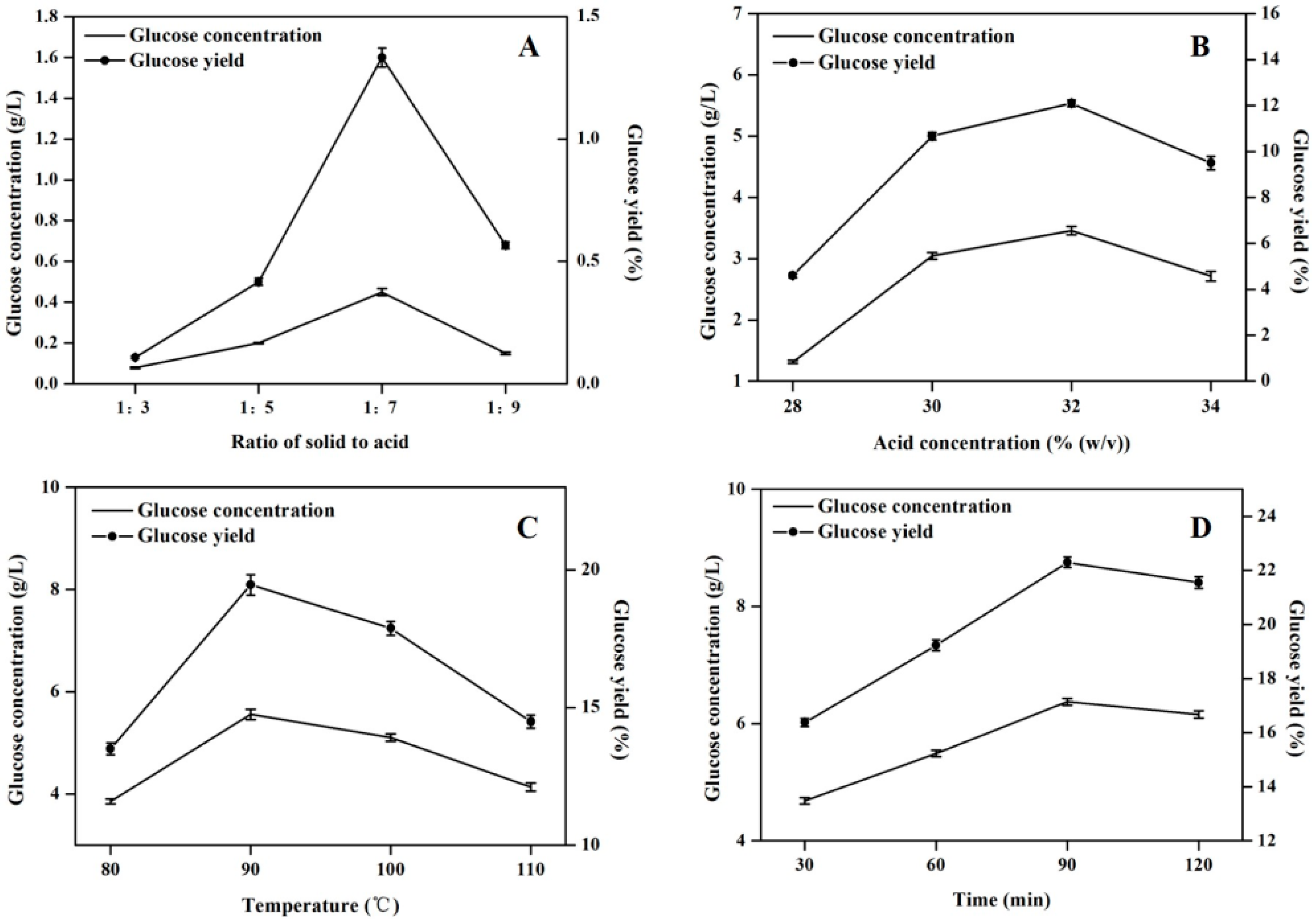
2.2. Optimization of Acid Hydrolysis Condition
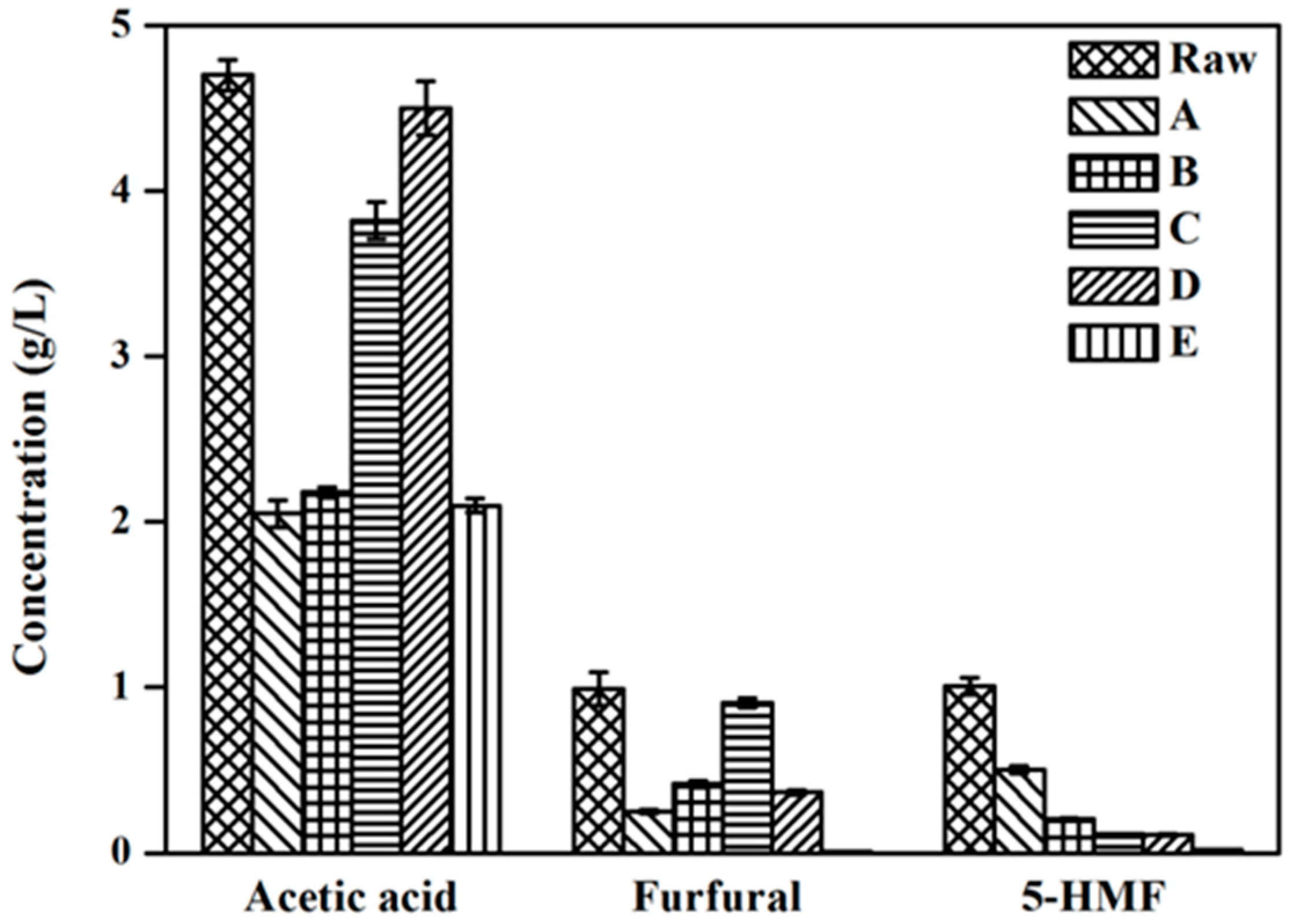
2.3. Inhibitor Changes with Different Detoxification Methods
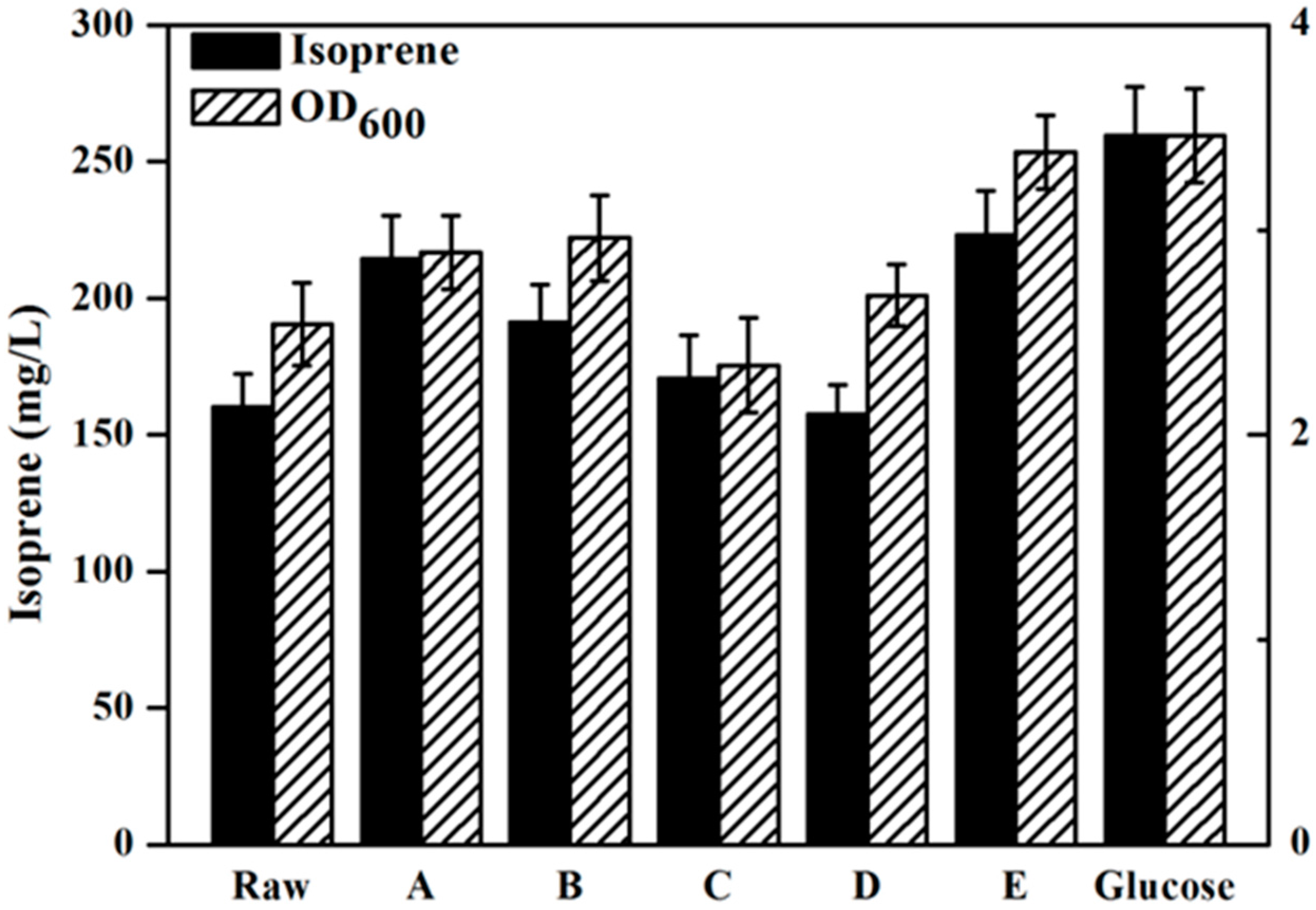
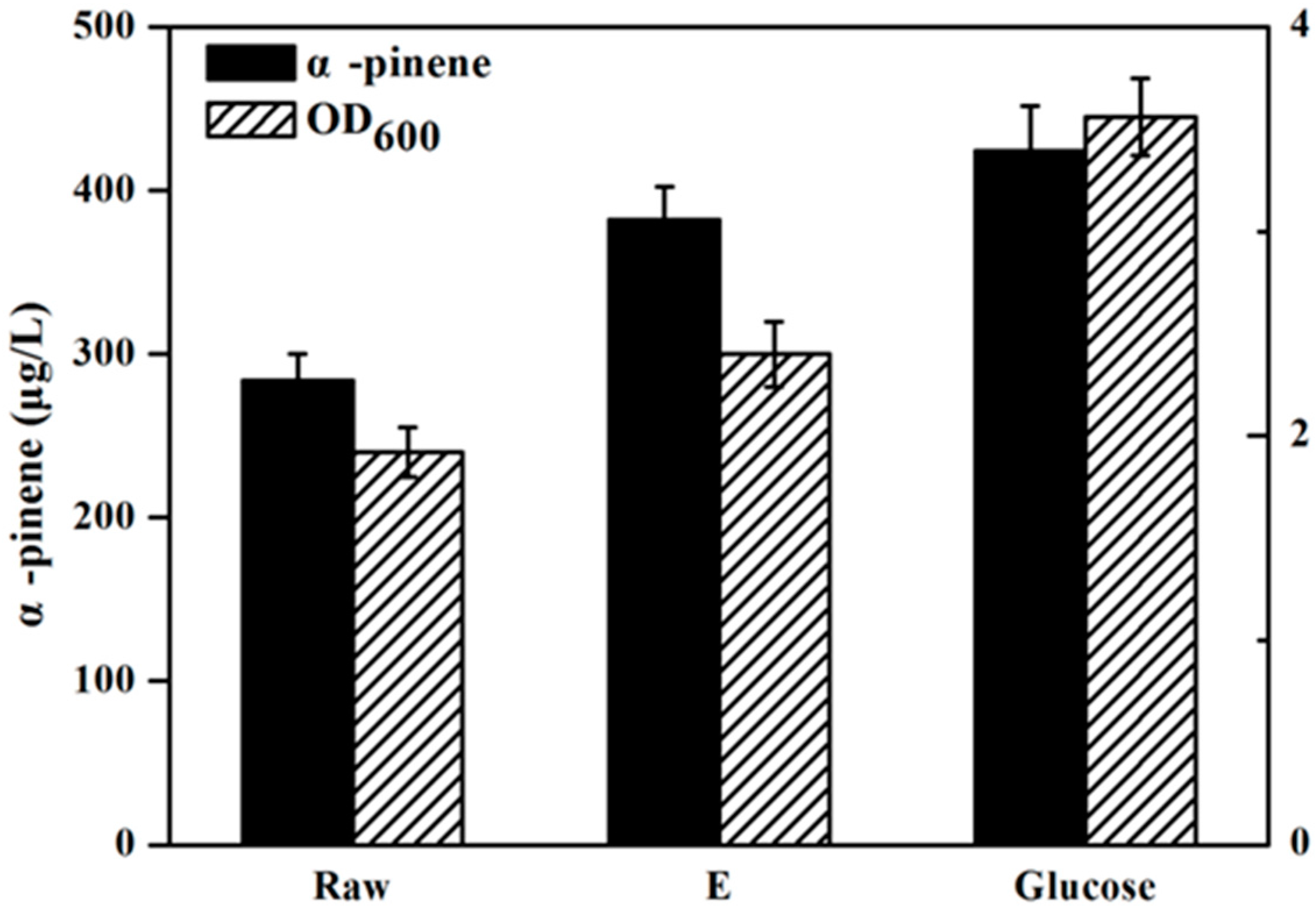
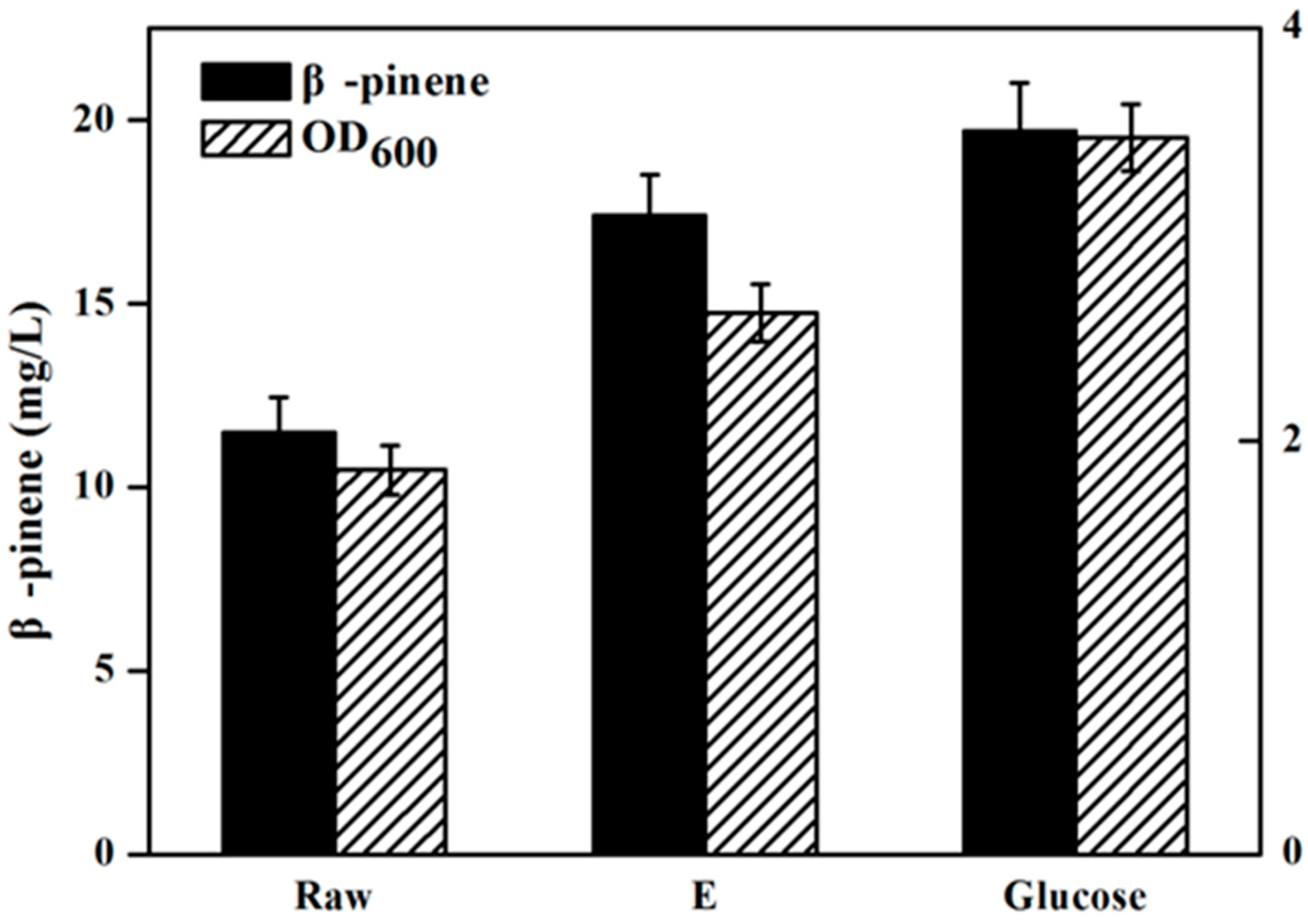
2.4. Effect of Detoxification on Isoprenoids Production
3. Methods and Materials
3.1. Materials
3.2. Compositions Analysis
3.3. Optimization of the Acid Hydrolysis Process
3.4. Inhibitor and Sugar Analysis of the Hydrolysates
3.5. Detoxification
- The hydrolysate was adjusted to pH 10 with sodium hydroxide, and then the solution was readjusted with sulfuric acid to pH 5. Anhydrous sodium sulphite (1 g/L) was added to the solution which was heated to 100 °C for 15 min. Then, 1% (w/v) activated carbon was mixed into the solution and incubated at 40 °C with shaking at 200 rpm for 1 h.
- The hydrolysate was neutralized with calcium hydroxide. After that, 1% (w/v) activated carbon was added into the solution and incubated at 40 °C with shaking at 200 rpm for 1 h.
- Sodium hydroxide was used to regulate the pH of the hydrolysate to 5.0, and then 1% (w/v) activated carbon was added into the solution and incubated 40 °C with shaking at 200 rpm for 1 h.
- Anion exchange resin (D301, Tianjin, China) was added to the hydrolysate with the loading concentration of 20% (w/v) until the pH reached 5.5. The mixed solution was kept at 24 °C, with shaking 200 rpm for 1 h.
- The pH of the hydrolysate was initially adjusted to 7.0 with calcium hydroxide, after that, the pH was readjusted to 5.5 with phosphoric acid.
3.6. Biosynthesis and Analysis of Isoprenoids Produced Using the Engineered E. coli Strains
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Zhao, Y.; Yang, J.; Qin, B.; Li, Y.; Sun, Y.; Su, S.; Xian, M. Biosynthesis of isoprene in Escherichia coli via methylerythritol phosphate (MEP) pathway. Appl. Microbiol. Biotechnol. 2011, 90, 1915–1922. [Google Scholar] [CrossRef] [PubMed]
- Alianell, G.A.; Derwitsch, F.; Wells, D.; Taylor, T. Isoprene Compositions and Methods of Use. U.S. Patent US20100099932 A1, 22 April 2010. [Google Scholar]
- Kesselmeier, J.; Staudt, M. Biogenic volatile organic compounds (VOC): An overview on emission, physiology and ecology. J. Atmos. Chem. 1999, 33, 23–88. [Google Scholar] [CrossRef]
- Yang, J.; Xian, M.; Su, S.; Zhao, G.; Nie, Q.; Jiang, X.; Zheng, Y.; Liu, W. Enhancing production of bio-isoprene using hybrid MVA pathway and isoprene synthase in E. coli. PLoS ONE 2012, 7, e33509. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Nie, Q.; Ren, M.; Feng, H.; Jiang, X.; Zheng, Y.; Liu, M.; Zhang, H.; Xian, M. Metabolic engineering of Escherichia coli for the biosynthesis of alpha-pinene. Biotechnol. Biofuels 2013, 6, 60. [Google Scholar] [CrossRef] [PubMed]
- Larsson, S.; Reimann, A.; Nilvebrant, N.-O.; Jönsson, L.J. Comparison of different methods for the detoxification of lignocellulose hydrolyzates of spruce. Appl. Biochem. Biotechnol. 1999, 77, 91–103. [Google Scholar] [CrossRef]
- Agblevor, F.; Fu, J.; Hames, B.; McMillan, J. Identification of microbial inhibitory functional groups in corn stover hydrolysate by carbon-13 nuclear magnetic resonance spectroscopy. Appl. Biochem. Biotechnol. 2004, 119, 97–120. [Google Scholar] [CrossRef]
- Canilha, L.; de Almeida e Silva, J.B.; Solenzal, A.I.N. Eucalyptus hydrolysate detoxification with activated charcoal adsorption or ion-exchange resins for xylitol production. Process Biochem. 2004, 39, 1909–1912. [Google Scholar] [CrossRef]
- Martinez, A.; Rodriguez, M.E.; York, S.W.; Preston, J.F.; Ingram, L.O. Effects of Ca(OH)2 treatments (overliming) on the composition and toxicity of bagasse hemicellulose hydrolysates. Biotechnol. Bioeng. 2000, 69, 526–536. [Google Scholar] [CrossRef]
- Mpabanga, T.P.; Chandel, A.K.; da Silva, S.S.; Singh, O.V. Detoxification Strategies Applied to Lignocellulosic Hydrolysates for Improved Xylitol Production. In D-Xylitol; Springer: Cham, Switzerland, 2012; pp. 63–82. [Google Scholar]
- Chen, L.; Liu, T.; Zhang, W.; Chen, X.; Wang, J. Biodiesel production from algae oil high in free fatty acids by two-step catalytic conversion. Bioresour. Technol. 2012, 111, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.; Ragauskas, A. Pretreatment and lignocellulosic chemistry. Bioenergy Res. 2012, 5, 1043–1066. [Google Scholar] [CrossRef]
- Harun, R.; Danquah, M.K. Influence of acid pre-treatment on microalgal biomass for bioethanol production. Process Biochem. 2011, 46, 304–309. [Google Scholar] [CrossRef]
- Wi, S.G.; Cho, E.J.; Lee, D.-S.; Lee, S.J.; Lee, Y.J.; Bae, H.-J. Lignocellulose conversion for biofuel: A new pretreatment greatly improves downstream biocatalytic hydrolysis of various lignocellulosic materials. Biotechnol. Biofuels 2015, 8, 228. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.H.; Kim, I.J.; Kim, H.K.; Kim, K.H. Dilute acid pretreatment of lignocellulose for whole slurry ethanol fermentation. Bioresour. Technol. 2013, 132, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Parawira, W.; Tekere, M. Biotechnological strategies to overcome inhibitors in lignocellulose hydrolysates for ethanol production: Review. Crit. Rev. Biotechnol. 2011, 31, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Jönsson, L.J.; Alriksson, B.; Nilvebrant, N.-O. Bioconversion of lignocellulose: Inhibitors and detoxification. Biotechnol. Biofuels 2013, 6, 16. [Google Scholar] [CrossRef] [PubMed]
- Chandel, A.K.; Singh, O.V.; da Silva, S.S. Detoxification of Lignocellulosic Hydrolysates for Improved Bioethanol Production. In Biofuel Production-Recent Developments and Prospects; INTECH Open Access Publisher: Rijeka, Croatia, 2011. [Google Scholar]
- Palmqvist, E.; Almeida, J.S.; Hahn-Hägerdal, B. Influence of furfural on anaerobic glycolytic kinetics of Saccharomyces cerevisiae in batch culture. Biotechnol. Bioeng. 1999, 62, 447–454. [Google Scholar] [CrossRef]
- Taherzadeh, M.J.; Gustafsson, L.; Niklasson, C.; Lidén, G. Conversion of furfural in aerobic and anaerobic batch fermentation of glucose by Saccharomyces cerevisiae. J. Biosci. Bioeng. 1999, 87, 169–174. [Google Scholar] [CrossRef]
- Palmqvist, E.; Hahn-Hägerdal, B. Fermentation of lignocellulosic hydrolysates. II: Inhibitors and mechanisms of inhibition. Bioresour. Technol. 2000, 74, 25–33. [Google Scholar] [CrossRef]
- Mills, T.Y.; Sandoval, N.R.; Gill, R.T. Cellulosic hydrolysate toxicity and tolerance mechanisms in Escherichia coli. Biotechnol. Biofuels 2009, 2, 1. [Google Scholar] [CrossRef] [PubMed]
- Chandel, A.K.; Da Silva, S.S.; Singh, O.V. Detoxification of lignocellulose hydrolysates: Biochemical and metabolic engineering toward white biotechnology. Bioenergy Res. 2013, 6, 388–401. [Google Scholar] [CrossRef]
- Sluiter, A.D.; Hames, B.; Ruiz, R.O.; Scarlata, C.; Sluiter, J.; Templeton, D.W.; Crocker, D. Determination of structural carbohydrates and lignin in biomass. Lab. Anal. Proced. 2008; NREL/TP; 510-42618. 1617. [Google Scholar]
- Ho, S.-H.; Huang, S.-W.; Chen, C.-Y.; Hasunuma, T.; Kondo, A.; Chang, J.-S. Bioethanol production using carbohydrate-rich microalgae biomass as feedstock. Bioresour. Technol. 2013, 135, 191–198. [Google Scholar] [CrossRef] [PubMed]
- luiter A, H.B.; Ruiz, R.; Scarlata, C.; Sluiter, J.; Templeton, D. Determination of Ash in Biomass Laboratory Analytical Procedure (LAP); National Renewable Energy Laboratory: Golden, CO, USA, 2005.
- Standard, T. Solvent Extractives of Wood and Pulp. In TAPPI T 204 cm-97; TAPPI Press: Atlanta, GA, USA, 1997. [Google Scholar]
- Hongru Feng, J.Y.; Li, Q.I.N.; Mo, X.I.A.N. Expression of β-pinene synthase (QH6) in Escherichia coli for the biosynthesis of β-pinene. Chin. J. Bioprocess Eng. 2015, 13, 28–34. [Google Scholar]
- Yang, J.; Zhao, G.; Sun, Y.; Zheng, Y.; Jiang, X.; Liu, W.; Xian, M. Bio-isoprene production using exogenous MVA pathway and isoprene synthase in Escherichia coli. Bioresour. Technol. 2012, 104, 642–647. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds holocellulose, water, lipid, ash, protein are available from the authors. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Optimization Factor | Range of Optimization Factor | Constant Factors |
|---|---|---|
| Ratio of solid to acid | 1:3, 1:5, 1:7, 1:9 | 20% (w/v) concentration of sulfuric acid solution, at 80 °C for 30 min |
| Concentration of sulfuric acid | 28% (w/v), 30% (w/v), 32% (w/v), 34% (w/v) | 1:7 ratio of solid to acid solution, at 80 °C for 30 min |
| temperature | 80 °C, 90 °C, 100 °C,110 °C | 1:7 ratio of solid to acid solution and 32% (w/v) concentration of sulfuric acid solution for 30 min |
| Time | 30 min, 60 min, 90 min, 120 min | 1:7 ratio of solid to acid solution and 32% (w/v) concentration of sulfuric acid solution, at 90 °C |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, S.; Yang, J. Isoprenoids Production from Lipid-Extracted Microalgal Biomass Residues Using Engineered E. coli. Molecules 2017, 22, 960. https://doi.org/10.3390/molecules22060960
Wang S, Yang J. Isoprenoids Production from Lipid-Extracted Microalgal Biomass Residues Using Engineered E. coli. Molecules. 2017; 22(6):960. https://doi.org/10.3390/molecules22060960
Chicago/Turabian StyleWang, Sumeng, and Jianming Yang. 2017. "Isoprenoids Production from Lipid-Extracted Microalgal Biomass Residues Using Engineered E. coli" Molecules 22, no. 6: 960. https://doi.org/10.3390/molecules22060960
APA StyleWang, S., & Yang, J. (2017). Isoprenoids Production from Lipid-Extracted Microalgal Biomass Residues Using Engineered E. coli. Molecules, 22(6), 960. https://doi.org/10.3390/molecules22060960