Recent Advances in the Inhibition of p38 MAPK as a Potential Strategy for the Treatment of Alzheimer’s Disease

Abstract
:1. Introduction
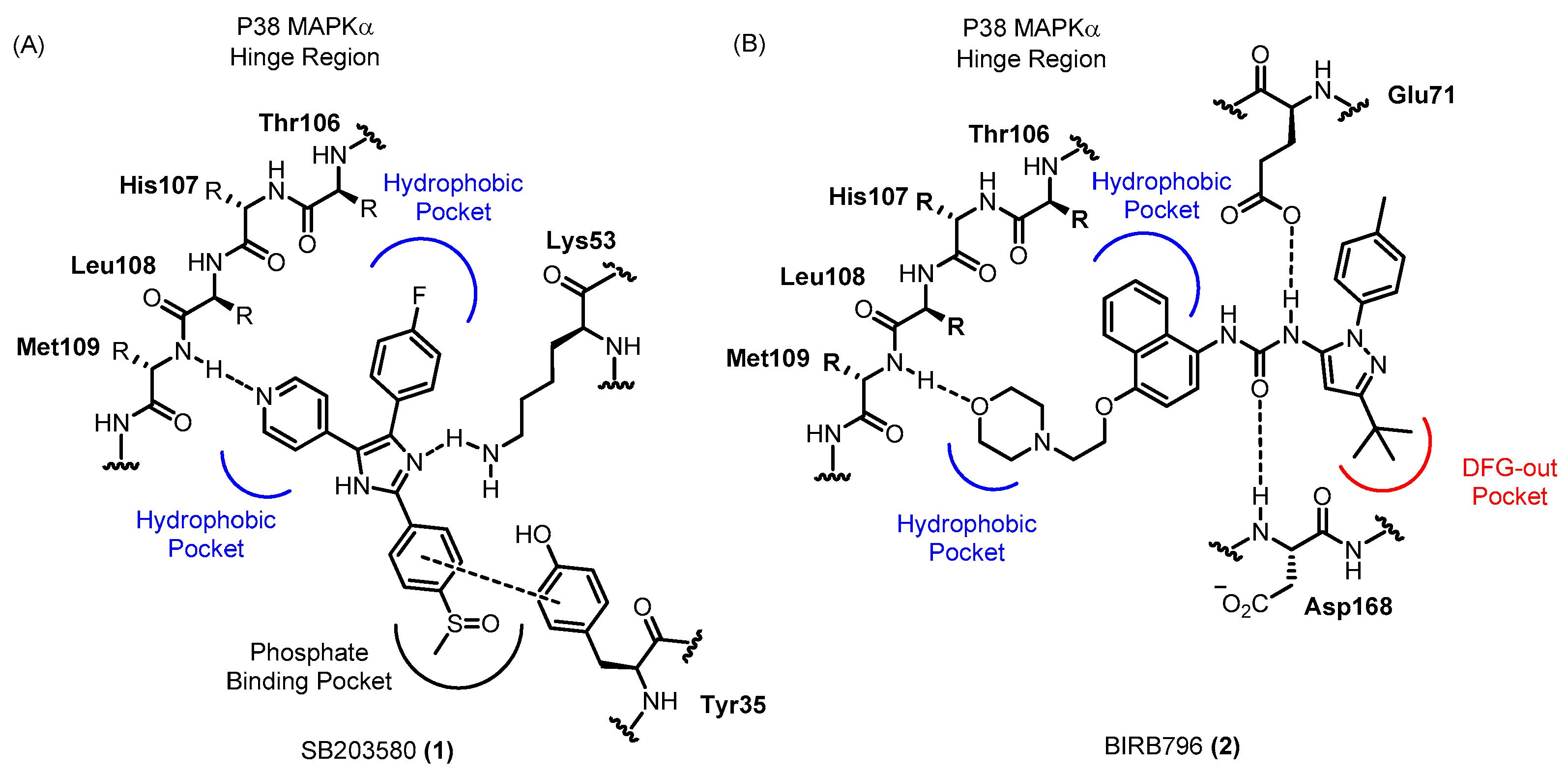
1.1. P38 Mitogen-Activated Protein Kinase (P38 MAPK) and Its Inhibitors
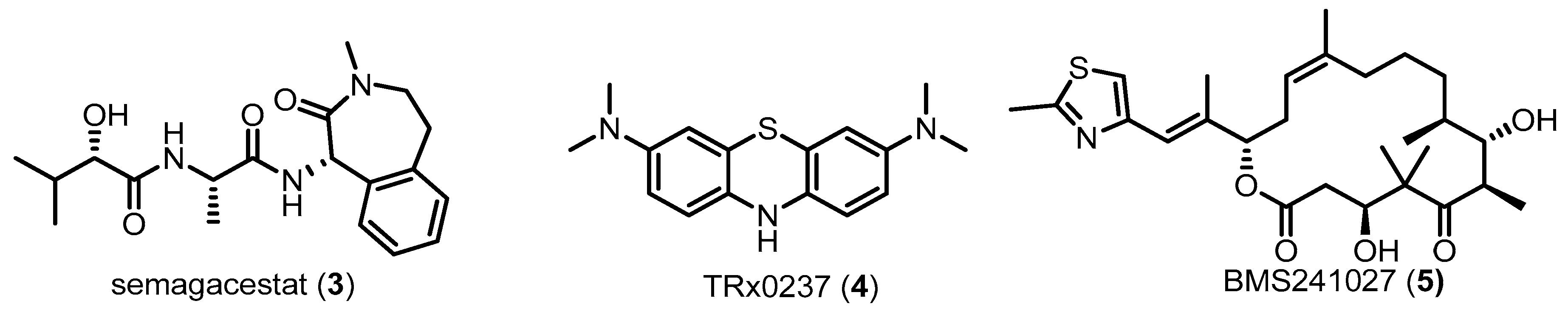
1.2. Alzheimer’s Disease (AD) and Recent Drug Candidates Targeting AD Pathologies
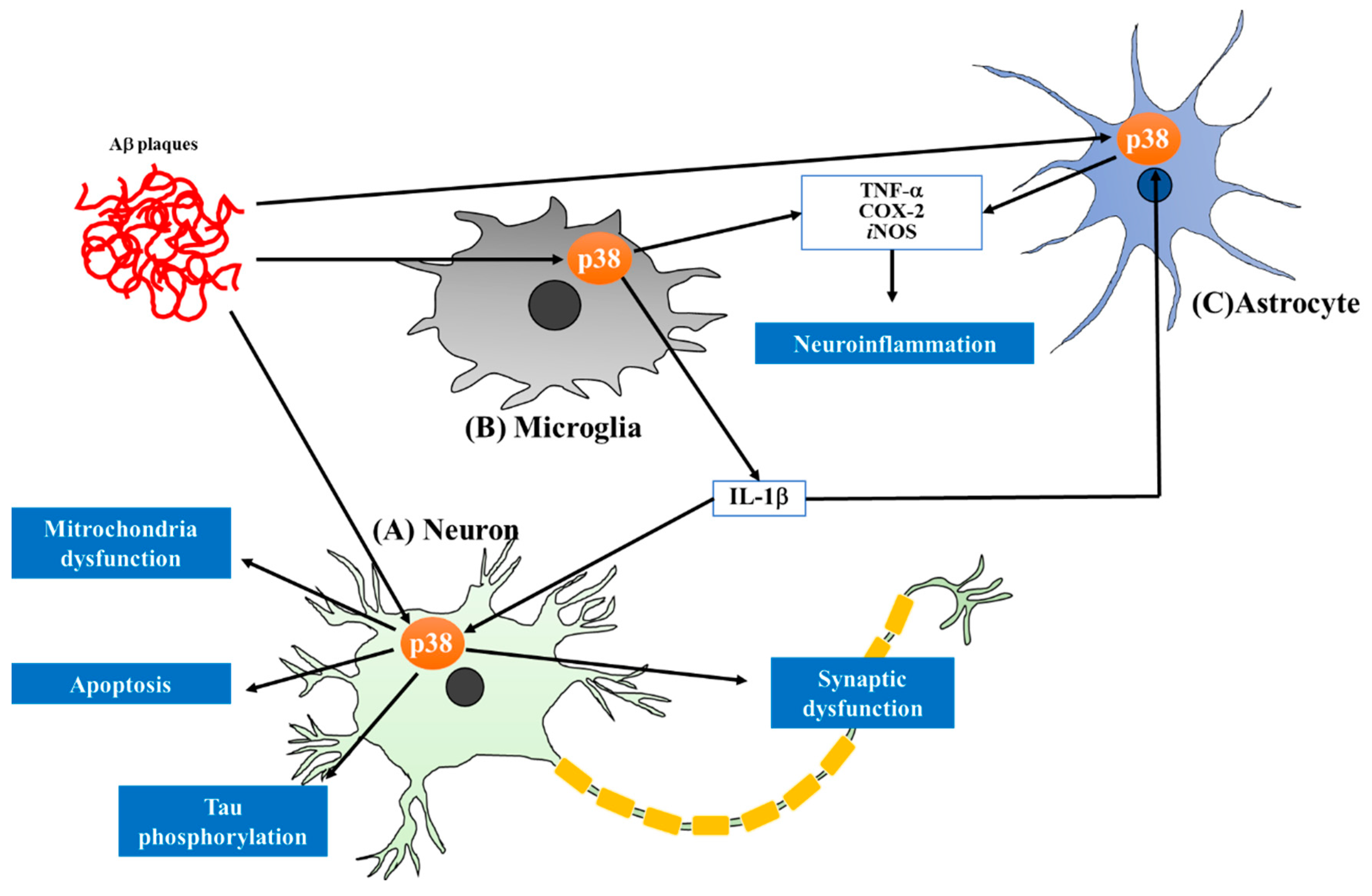
2. Recent Advances in the Study of p38 MAPK and Its Inhibition in AD Pathology
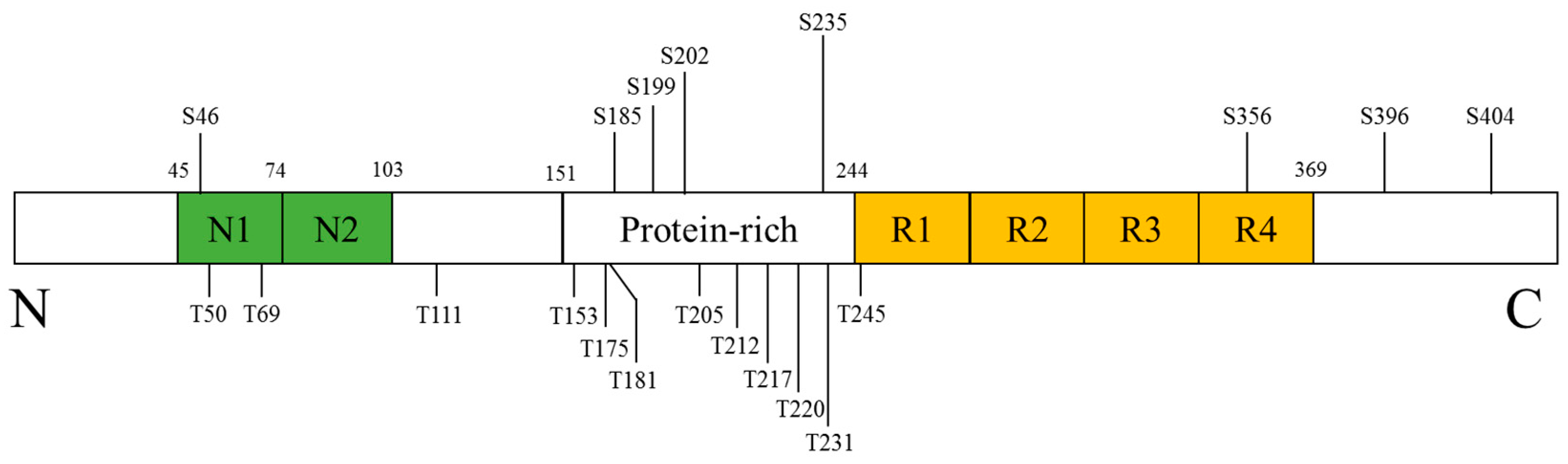
2.1. Targeting the Tau Pathology via p38 Inhibition in Neuronal Cells
2.2. Neuroprotective Effect of p38 Inhibition against Aβ-Induced Neuronal Damage
2.3. Reduction of Neuroinflammation by Inhibiting p38MAPK Pathway in Microglia and Astrocytes
2.4. Improvement of Synaptic Plasticity by p38 Inhibition
3. Conclusions and Perspectives
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| AD | Alzheimer’s disease |
| FDA | Food and Drug Administration |
| AchE | acetylcholinesterase |
| NMDAR | N-methyl-d-aspartate receptor |
| MAPK | mitogen-activated protein kinase |
| ROS | reactive oxygen species |
| JNK | c-jun N-terminal kinase |
| ERK | extracellular signal-regulated kinase |
| TNF-α | tumor necrosis factor-α |
| IL-1 | interleukin-1 |
| LPS | lipopolysaccharide |
| ATP | adenosine triphosphate |
| CNS | central nervous systems |
| Aβ | amyloid-β |
| NFTs | neurofibrillary tangles |
| APP | amyloid precursor protein |
| PS | presenilin |
| NO | nitric oxide |
| NSAIDs | nonsteroidal anti-inflammatory drugs |
| iNOS | inducible nitric oxide synthase |
| NF-kB | nuclear factor kappa-light-chain-enhancer of activated B cells |
| LTP | long-term potentiation |
| LTD | long-term depression |
| mGluR | metabotropic glutamate receptor |
| AMPAR | α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptor |
| BBB | blood-brain barrier |
References
- Prince, M.; Bryce, R.; Albanese, E.; Wimo, A.; Ribeiro, W.; Ferri, C.P. The global prevalence of dementia: A systematic review and metaanalysis. Alzheimers Dement. 2013, 9, 63–75.e2. [Google Scholar] [CrossRef] [PubMed]
- Mangialasche, F.; Solomon, A.; Winblad, B.; Mecocci, P.; Kivipelto, M. Alzheimer’s disease: Clinical trials and drug development. Lancet Neurol. 2010, 9, 702–716. [Google Scholar] [CrossRef]
- Van Cauwenberghe, C.; Van Broeckhoven, C.; Sleegers, K. The genetic landscape of alzheimer disease: Clinical implications and perspectives. Genet. Med. 2016, 18, 421–430. [Google Scholar] [CrossRef] [PubMed]
- Grant, S.K. Therapeutic protein kinase inhibitors. Cell. Mol. Life Sci. 2009, 66, 1163–1177. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.Y.; Mei, Z.Q.; Wu, J.W.; Wang, Z.X. Enzymatic activity and substrate specificity of mitogen-activated protein kinase p38α in different phosphorylation states. J. Biol. Chem. 2008, 283, 26591–26601. [Google Scholar] [CrossRef] [PubMed]
- Sun, A.; Liu, M.; Nguyen, X.V.; Bing, G. P38 map kinase is activated at early stages in Alzheimer’s disease brain. Exp. Neurol. 2003, 183, 394–405. [Google Scholar] [CrossRef]
- Hensley, K.; Floyd, R.A.; Zheng, N.Y.; Nael, R.; Robinson, K.A.; Nguyen, X.; Pye, Q.N.; Stewart, C.A.; Geddes, J.; Markesbery, W.R.; et al. P38 kinase is activated in the Alzheimer’s disease brain. J. Neurochem. 1999, 72, 2053–2058. [Google Scholar] [CrossRef] [PubMed]
- Schnoder, L.; Hao, W.; Qin, Y.; Liu, S.; Tomic, I.; Liu, X.; Fassbender, K.; Liu, Y. Deficiency of neuronal p38α MAPK attenuates amyloid pathology in Alzheimer disease mouse and cell models through facilitating lysosomal degradation of BACE1. J. Biol. Chem. 2016, 291, 2067–2079. [Google Scholar] [CrossRef] [PubMed]
- Munoz, L.; Ammit, A.J. Targeting p38 mapk pathway for the treatment of Alzheimer’s disease. Neuropharmacology 2010, 58, 561–568. [Google Scholar] [CrossRef] [PubMed]
- Yokota, T.; Wang, Y. P38 map kinases in the heart. Gene 2016, 575, 369–376. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, S.; Sugiura, H.; Tanaka, H.; Takigami, S.; Yamagata, K. P38 MAP kinase inhibitors as potential therapeutic drugs for neural diseases. Cent. Nerv. Syst. Agents Med. Chem. 2011, 11, 45–59. [Google Scholar] [CrossRef] [PubMed]
- Marber, M.S.; Rose, B.; Wang, Y. The p38 mitogen-activated protein kinase pathway—A potential target for intervention in infarction, hypertrophy, and heart failure. J. Mol. Cell. Cardiol. 2011, 51, 485–490. [Google Scholar] [CrossRef] [PubMed]
- Denise Martin, E.; De Nicola, G.F.; Marber, M.S. New therapeutic targets in cardiology: P38α mitogen-activated protein kinase for ischemic heart disease. Circulation 2012, 126, 357–368. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Boehm, J.; Lee, J.C. P38 map kinases: Key signalling molecules as therapeutic targets for inflammatory diseases. Nat. Rev. Drug Discov. 2003, 2, 717–726. [Google Scholar] [CrossRef] [PubMed]
- Cuenda, A.; Rousseau, S. P38 MAP-kinases pathway regulation, function and role in human diseases. Biochim. Biophys. Acta 2007, 1773, 1358–1375. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.S.; Diener, K.; Manthey, C.L.; Wang, S.; Rosenzweig, B.; Bray, J.; Delaney, J.; Cole, C.N.; Chan-Hui, P.Y.; Mantlo, N.; et al. Molecular cloning and characterization of a novel p38 mitogen-activated protein kinase. J. Biol. Chem. 1997, 272, 23668–23674. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.B.; Cheng, T.T.; Chindalore, V.; Damjanov, N.; Burgos-Vargas, R.; Delora, P.; Zimany, K.; Travers, H.; Caulfield, J.P. Evaluation of the efficacy and safety of pamapimod, a p38 MAP kinase inhibitor, in a double-blind, methotrexate-controlled study of patients with active rheumatoid arthritis. Arthritis Rheum. 2009, 60, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Wang, M.; Zhang, J.; Xu, R. P38 MAPK: A potential target of chronic pain. Curr. Med. Chem. 2014, 21, 4405–4418. [Google Scholar] [CrossRef] [PubMed]
- Arthur, J.S.; Ley, S.C. Mitogen-activated protein kinases in innate immunity. Nat. Rev. Immunol. 2013, 13, 679–692. [Google Scholar] [CrossRef] [PubMed]
- Astolfi, A.; Iraci, N.; Manfroni, G.; Barreca, M.L.; Cecchetti, V. A comprehensive structural overview of p38α MAPK in complex with type I inhibitors. ChemMedChem 2015, 10, 957–969. [Google Scholar] [CrossRef] [PubMed]
- Cohen, P. Targeting protein kinases for the development of anti-inflammatory drugs. Curr. Opin. Cell Biol. 2009, 21, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.R.; Dominguez, C. Map kinase p38 inhibitors: Clinical results and an intimate look at their interactions with p38α protein. Curr. Med. Chem. 2005, 12, 2979–2994. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Shen, B.; Lin, A. Novel strategies for inhibition of the p38 MAPK pathway. Trends Pharmacol. Sci. 2007, 28, 286–295. [Google Scholar] [CrossRef] [PubMed]
- Aouadi, M.; Bost, F.; Caron, L.; Laurent, K.; Le Marchand Brustel, Y.; Binetruy, B. P38 mitogen-activated protein kinase activity commits embryonic stem cells to either neurogenesis or cardiomyogenesis. Stem Cells 2006, 24, 1399–1406. [Google Scholar] [CrossRef] [PubMed]
- Poolos, N.P.; Bullis, J.B.; Roth, M.K. Modulation of h-channels in hippocampal pyramidal neurons by p38 mitogen-activated protein kinase. J. Neurosci. 2006, 26, 7995–8003. [Google Scholar] [CrossRef] [PubMed]
- Zhong, P.; Liu, W.; Gu, Z.; Yan, Z. Serotonin facilitates long-term depression induction in prefrontal cortex via p38 Mapk/RAB5-mediated enhancement of ampa receptor internalization. J. Physiol. 2008, 586, 4465–4479. [Google Scholar] [CrossRef] [PubMed]
- Bachstetter, A.D.; Xing, B.; de Almeida, L.; Dimayuga, E.R.; Watterson, D.M.; Van Eldik, L.J. Microglial p38α MAPK is a key regulator of proinflammatory cytokine up-regulation induced by toll-like receptor (TLR) ligands or β-amyloid (Aβ). J. Neuroinflamm. 2011, 8, 79. [Google Scholar] [CrossRef] [PubMed]
- Ashabi, G.; Alamdary, S.Z.; Ramin, M.; Khodagholi, F. Reduction of hippocampal apoptosis by intracerebroventricular administration of extracellular signal-regulated protein kinase and/or p38 inhibitors in amyloid β rat model of Alzheimer’s disease: Involvement of nuclear-related factor-2 and nuclear factor-kappaB. Basic Clin. Pharmacol. Toxicol. 2013, 112, 145–155. [Google Scholar] [PubMed]
- Xuan, A.; Long, D.; Li, J.; Ji, W.; Zhang, M.; Hong, L.; Liu, J. Hydrogen sulfide attenuates spatial memory impairment and hippocampal neuroinflammation in β-amyloid rat model of Alzheimer’s disease. J. Neuroinflamm. 2012, 9, 202. [Google Scholar] [CrossRef] [PubMed]
- Burns, A.; Iliffe, S. Alzheimer’s disease. BMJ 2009, 338, b158. [Google Scholar] [CrossRef] [PubMed]
- Forman, M.S.; Trojanowski, J.Q.; Lee, V.M. Neurodegenerative diseases: A decade of discoveries paves the way for therapeutic breakthroughs. Nat. Med. 2004, 10, 1055–1063. [Google Scholar] [CrossRef] [PubMed]
- Kurz, A.; Perneczky, R. Novel insights for the treatment of Alzheimer’s disease. Prog. Neuropsychopharmacol. Biol. Psychiatry 2011, 35, 373–379. [Google Scholar] [CrossRef] [PubMed]
- Karran, E.; De Strooper, B. The amyloid cascade hypothesis: Are we poised for success or failure? J. Neurochem. 2016, 139 (Suppl. 2), 237–252. [Google Scholar] [CrossRef] [PubMed]
- Gandy, S.; DeKosky, S.T. Toward the treatment and prevention of Alzheimer’s disease: Rational strategies and recent progress. Annu. Rev. Med. 2013, 64, 367–383. [Google Scholar] [CrossRef] [PubMed]
- Benilova, I.; De Strooper, B. An overlooked neurotoxic species in Alzheimer’s disease. Nat. Neurosci. 2011, 14, 949–950. [Google Scholar] [CrossRef] [PubMed]
- Saito, T.; Suemoto, T.; Brouwers, N.; Sleegers, K.; Funamoto, S.; Mihira, N.; Matsuba, Y.; Yamada, K.; Nilsson, P.; Takano, J.; et al. Potent amyloidogenicity and pathogenicity of Aβ43. Nat. Neurosci. 2011, 14, 1023–1032. [Google Scholar] [CrossRef] [PubMed]
- Holtta, M.; Dean, R.A.; Siemers, E.; Mawuenyega, K.G.; Sigurdson, W.; May, P.C.; Holtzman, D.M.; Portelius, E.; Zetterberg, H.; Bateman, R.J.; et al. A single dose of the gamma-secretase inhibitor semagacestat alters the cerebrospinal fluid peptidome in humans. Alzheimers Res. Ther. 2016, 8, 11. [Google Scholar] [CrossRef] [PubMed]
- Ivanoiu, A.; Pariente, J.; Booth, K.; Lobello, K.; Luscan, G.; Hua, L.; Lucas, P.; Styren, S.; Yang, L.; Li, D.; et al. Long-term safety and tolerability of bapineuzumab in patients with Alzheimer’s disease in two phase 3 extension studies. Alzheimers Res. Ther. 2016, 8, 24. [Google Scholar] [CrossRef] [PubMed]
- Liu-Seifert, H.; Siemers, E.; Price, K.; Han, B.; Selzler, K.J.; Henley, D.; Sundell, K.; Aisen, P.; Cummings, J.; Raskin, J.; et al. Cognitive impairment precedes and predicts functional impairment in mild Alzheimer’s disease. J. Alzheimers Dis. 2015, 47, 205–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, M.; Maeda, S.; Vossel, K.; Mucke, L. The many faces of tau. Neuron 2011, 70, 410–426. [Google Scholar] [CrossRef] [PubMed]
- Braak, H.; Braak, E. Neuropathological stageing of Alzheimer-related changes. Acta Neuropathol. 1991, 82, 239–259. [Google Scholar] [CrossRef] [PubMed]
- Krstic, D.; Knuesel, I. Deciphering the mechanism underlying late-onset Alzheimer disease. Nat. Rev. Neurol. 2013, 9, 25–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forner, S.; Baglietto-Vargas, D.; Martini, A.C.; Trujillo-Estrada, L.; LaFerla, F.M. Synaptic impairment in Alzheimer’s disease: A dysregulated symphony. Trends Neurosci. 2017, 40, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Wischik, C.; Staff, R. Challenges in the conduct of disease-modifying trials in ad: Practical experience from a phase 2 trial of tau-aggregation inhibitor therapy. J. Nutr. Health Aging 2009, 13, 367–369. [Google Scholar] [CrossRef] [PubMed]
- Holmes, C.; Cunningham, C.; Zotova, E.; Woolford, J.; Dean, C.; Kerr, S.; Culliford, D.; Perry, V.H. Systemic inflammation and disease progression in Alzheimer disease. Neurology 2009, 73, 768–774. [Google Scholar] [CrossRef] [PubMed]
- Mohammadzadeh Honarvar, N.; Saedisomeolia, A.; Abdolahi, M.; Shayeganrad, A.; Taheri Sangsari, G.; Hassanzadeh Rad, B.; Muench, G. Molecular anti-inflammatory mechanisms of retinoids and carotenoids in Alzheimer’s disease: A review of current evidence. J. Mol. Neurosci. 2017, 61, 289–304. [Google Scholar] [CrossRef] [PubMed]
- Tan, Z.S.; Beiser, A.S.; Vasan, R.S.; Roubenoff, R.; Dinarello, C.A.; Harris, T.B.; Benjamin, E.J.; Au, R.; Kiel, D.P.; Wolf, P.A.; et al. Inflammatory markers and the risk of Alzheimer disease: The framingham study. Neurology 2007, 68, 1902–1908. [Google Scholar] [CrossRef] [PubMed]
- Tuppo, E.E.; Arias, H.R. The role of inflammation in Alzheimer’s disease. Int. J. Biochem. Cell Biol. 2005, 37, 289–305. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Singh, A.; Ekavali. A review on Alzheimer’s disease pathophysiology and its management: An update. Pharmacol. Rep. 2015, 67, 195–203. [Google Scholar] [CrossRef] [PubMed]
- McGeer, P.L.; Schulzer, M.; McGeer, E.G. Arthritis and anti-inflammatory agents as possible protective factors for Alzheimer’s disease: A review of 17 epidemiologic studies. Neurology 1996, 47, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Scharf, S.; Mander, A.; Ugoni, A.; Vajda, F.; Christophidis, N. A double-blind, placebo-controlled trial of diclofenac/misoprostol in Alzheimer’s disease. Neurology 1999, 53, 197–201. [Google Scholar] [CrossRef] [PubMed]
- Koch, H.J.; Szecsey, A. A randomized controlled trial of prednisone in Alzheimer’s disease. Neurology 2000, 55, 1067. [Google Scholar] [CrossRef] [PubMed]
- Hoozemans, J.J.; O’Banion, M.K. The role of COX-1 and COX-2 in Alzheimer’s disease pathology and the therapeutic potentials of non-steroidal anti-inflammatory drugs. Curr. Drug Targets CNS Neurol. Disord. 2005, 4, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Robinson, K.A.; Stewart, C.A.; Pye, Q.N.; Nguyen, X.; Kenney, L.; Salzman, S.; Floyd, R.A.; Hensley, K. Redox-sensitive protein phosphatase activity regulates the phosphorylation state of p38 protein kinase in primary astrocyte culture. J. Neurosci. Res. 1999, 55, 724–732. [Google Scholar] [CrossRef]
- Pei, J.J.; Braak, E.; Braak, H.; Grundke-Iqbal, I.; Iqbal, K.; Winblad, B.; Cowburn, R.F. Localization of active forms of c-jun kinase (JNK) and p38 kinase in Alzheimer’s disease brains at different stages of neurofibrillary degeneration. J. Alzheimers Dis. 2001, 3, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.W.; Rottkamp, C.A.; Hartzler, A.; Sun, Z.; Takeda, A.; Boux, H.; Shimohama, S.; Perry, G.; Smith, M.A. Activation of MKK6, an upstream activator of p38, in Alzheimer’s disease. J. Neurochem. 2001, 79, 311–318. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhang, C.; Sheng, X.N.; Zhang, X.W.; Wang, B.B.; Zhang, G.H. Peripheral expression of mapk pathways in Alzheimer’s and parkinson’s diseases. J. Clin. Neurosci. 2014, 21, 810–814. [Google Scholar] [CrossRef] [PubMed]
- Caceres, A.; Kosik, K.S. Inhibition of neurite polarity by tau antisense oligonucleotides in primary cerebellar neurons. Nature 1990, 343, 461–463. [Google Scholar] [CrossRef] [PubMed]
- Lindwall, G.; Cole, R.D. Phosphorylation affects the ability of tau protein to promote microtubule assembly. J. Biol. Chem. 1984, 259, 5301–5305. [Google Scholar] [PubMed]
- Grundke-Iqbal, I.; Iqbal, K.; Tung, Y.C.; Quinlan, M.; Wisniewski, H.M.; Binder, L.I. Abnormal phosphorylation of the microtubule-associated protein tau (TAU) in Alzheimer cytoskeletal pathology. Proc. Natl. Acad. Sci. USA 1986, 83, 4913–4917. [Google Scholar] [CrossRef] [PubMed]
- Tenreiro, S.; Eckermann, K.; Outeiro, T.F. Protein phosphorylation in neurodegeneration: Friend or foe? Front. Mol. Neurosci. 2014, 7, 42. [Google Scholar] [CrossRef] [PubMed]
- Churcher, I. Tau therapeutic strategies for the treatment of Alzheimer’s disease. Curr. Top. Med. Chem. 2006, 6, 579–595. [Google Scholar] [CrossRef] [PubMed]
- Cavallini, A.; Brewerton, S.; Bell, A.; Sargent, S.; Glover, S.; Hardy, C.; Moore, R.; Calley, J.; Ramachandran, D.; Poidinger, M.; et al. An unbiased approach to identifying tau kinases that phosphorylate tau at sites associated with Alzheimer disease. J. Biol. Chem. 2013, 288, 23331–23347. [Google Scholar] [CrossRef] [PubMed]
- Hanger, D.P.; Seereeram, A.; Noble, W. Mediators of tau phosphorylation in the pathogenesis of Alzheimer’s disease. Expert Rev. Neurother. 2009, 9, 1647–1666. [Google Scholar] [CrossRef] [PubMed]
- Sheng, J.G.; Jones, R.A.; Zhou, X.Q.; McGinness, J.M.; Van Eldik, L.J.; Mrak, R.E.; Griffin, W.S.T. Interleukin-1 promotion of MAPK-p38 overexpression in experimental animals and in Alzheimer’s disease: Potential significance for tau protein phosphorylation. Neurochem. Int. 2001, 39, 341–348. [Google Scholar] [CrossRef]
- Feijoo, C.; Campbell, D.G.; Jakes, R.; Goedert, M.; Cuenda, A. Evidence that phosphorylation of the microtubule-associated protein tau by SAPK4/p38 delta at THR50 promotes microtubule assembly. J. Cell Sci. 2005, 118, 397–408. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, H.; Goedert, M. Sequential phosphorylation of tau protein by cAMP-dependent protein kinase and SAPK4/p38 delta or JNK2 in the presence of heparin generates the AT100 epitope. J. Neurochem. 2006, 99, 154–164. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.Z.; Gao, X.; Wang, Z.H. The physiology and pathology of microtubule-associated protein tau. Essays Biochem. 2014, 56, 111–123. [Google Scholar] [CrossRef] [PubMed]
- Martin, L.; Latypova, X.; Terro, F. Post-translational modifications of tau protein: Implications for Alzheimer’s disease. Neurochem. Int. 2011, 58, 458–471. [Google Scholar] [CrossRef] [PubMed]
- Kelleher, I.; Garwood, C.; Hanger, D.P.; Anderton, B.H.; Noble, W. Kinase activities increase during the development of tauopathy in htau mice. J. Neurochem. 2007, 103, 2256–2267. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.W.; Rottkamp, C.A.; Boux, H.; Takeda, A.; Perry, G.; Smith, M.A. Activation of p38 kinase links tau phosphorylation, oxidative stress, and cell cycle-related events in Alzheimer disease. J. Neuropathol. Exp. Neurol. 2000, 59, 880–888. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.K.; Liu, L.; Barger, S.W.; Griffin, W.S.T. Interleukin-1 mediates pathological effects of microglia on tau phosphorylation and on synaptophysin synthesis in cortical neurons through a P38-MAPK pathway. J. Neurosci. 2003, 23, 1605–1611. [Google Scholar] [PubMed]
- Griffin, W.S.T.; Liu, L.; Li, Y.K.; Mrak, R.E.; Barger, S.W. Interleukin-1 mediates Alzheimer and lewy body pathologies. J. Neuroinflamm. 2006, 3, 5. [Google Scholar] [CrossRef] [PubMed]
- Tanji, K.; Mori, F.; Imaizumi, T.; Yoshida, H.; Satoh, K.; Wakabayashi, K. Interleukin-1 induces tau phosphorylation and morphological changes in cultured human astrocytes. Neuroreport 2003, 14, 413–417. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Chu, Y.Q.; Zhang, L.; Yin, L.L.; Li, L. Ginsenoside Rg1 attenuates tau phosphorylation in SK-N-SH induced by Aβ-stimulated THP-1 supernatant and the involvement of p38 pathway activation. Life Sci. 2012, 91, 809–815. [Google Scholar] [CrossRef] [PubMed]
- Giraldo, E.; Lloret, A.; Fuchsberger, T.; Vina, J. Aβ and tau toxicities in Alzheimer’s are linked via oxidative stress-induced p38 activation: Protective role of vitamine. Redox Biol. 2014, 2, 873–877. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.M.; Ma, J.Q.; Liu, S.S.; Zheng, G.H.; Feng, Z.J.; Sun, J.M. Proanthocyanidins improves lead-induced cognitive impairments by blocking endoplasmic reticulum stress and nuclear factor-kappaB-mediated inflammatory pathways in rats. Food Chem. Toxicol. 2014, 72, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Watterson, D.M.; Grum-Tokars, V.L.; Roy, S.M.; Schavocky, J.P.; Bradaric, B.D.; Bachstetter, A.D.; Xing, B.; Dimayuga, E.; Saeed, F.; Zhang, H.; et al. Development of novel in vivo chemical probes to address CNS protein kinase involvement in synaptic dysfunction. PLoS ONE 2013, 8, e66226. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, S.M.; Santana, I.; Swerdlow, R.H.; Oliveira, C.R. Mitochondria dysfunction of Alzheimer’s disease cybrids enhances Aβ toxicity. J. Neurochem. 2004, 89, 1417–1426. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, R.H.; Nagao, T.; Gouras, G.K. Plaque formation and the intraneuronal accumulation of β-amyloid in Alzheimer’s disease. Pathol. Int. 2017, 67, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Ji, L.; Zhao, X.; Lu, W.; Zhang, Q.; Hua, Z. Intracellular Aβ and its pathological role in Alzheimer’s disease: Lessons from cellular to animal models. Curr. Alzheimer Res. 2016, 13, 621–630. [Google Scholar] [CrossRef] [PubMed]
- Mattson, M.P. Calcium and neuronal injury in Alzheimer’s disease. Contributions of β-amyloid precursor protein mismetabolism, free radicals, and metabolic compromise. Ann. N. Y. Acad. Sci. 1994, 747, 50–76. [Google Scholar] [CrossRef] [PubMed]
- Calkins, M.J.; Reddy, P.H. Amyloid β impairs mitochondrial anterograde transport and degenerates synapses in Alzheimer’s disease neurons. Biochim. Biophys. Acta 2011, 1812, 507–513. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.H.; de Pablo, Y.; Lee, H.P.; Lee, H.G.; Smith, M.A.; Shah, K. Cdk5 is a major regulator of p38 cascade: Relevance to neurotoxicity in Alzheimer’s disease. J. Neurochem. 2010, 113, 1221–1229. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Teng, Y.; Zhang, X.; Lv, X.; Yin, Y. Metformin alleviated Aβ-induced apoptosis via the suppression of JNK mapk signaling pathway in cultured hippocampal neurons. Biomed. Res. Int. 2016, 2016, 1421430. [Google Scholar] [CrossRef] [PubMed]
- Suwanna, N.; Thangnipon, W.; Soi-Ampornkul, R. Neuroprotective effects of diarylpropionitrile against β-amyloid peptide-induced neurotoxicity in rat cultured cortical neurons. Neurosci. Lett. 2014, 578, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Zeng, K.W.; Wang, X.M.; Fu, H. Protective effect of cerebrospinal fluid containing jiawei wuzi yanzong formula on β-amyloid protein-induced injury of hippocampal neurons. Zhongguo Zhong Xi Yi Jie He Za Zhi 2010, 30, 851–856. [Google Scholar] [PubMed]
- Pierucci, F.; Garcia-Gil, M.; Frati, A.; Bini, F.; Martinesi, M.; Vannini, E.; Mainardi, M.; Luzzati, F.; Peretto, P.; Caleo, M.; et al. Vitamin D3 protects against Aβ peptide cytotoxicity in differentiated human neuroblastoma SH-SY5Y cells: A role for S1P1/p38MAPK/ATF4 axis. Neuropharmacology 2017, 116, 328–342. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Tan, Y.; Zheng, Y.; Du, X.; Liu, Q. Ebselen ameliorates β-amyloid pathology, tau pathology, and cognitive impairment in triple-transgenic Alzheimer’s disease mice. J. Biol. Inorg. Chem. 2017, 22, 851–865. [Google Scholar] [CrossRef] [PubMed]
- Fang, W.L.; Zhao, D.Q.; Wang, F.; Li, M.; Fan, S.N.; Liao, W.; Zheng, Y.Q.; Liao, S.W.; Xiao, S.H.; Luan, P.; et al. Neurotropin(r) alleviates hippocampal neuron damage through a HIF-1α/MAPK pathway. CNS Neurosci. Ther. 2017, 23, 428–437. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.Q.; Wang, Q.; Zhang, D.M.; Wang, J.Y.; Xiao, B.; Zheng, Y.; Wang, X.M. Triptolide rescues spatial memory deficits and amyloid-β aggregation accompanied by inhibition of inflammatory responses and mapks activity in APP/PS1 transgenic mice. Curr. Alzheimer Res. 2016, 13, 288–296. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Liu, M.; Yan, X. Effects of total glucosides of peony on expression of inflammatory cytokines and phosphorylated MAPK signal molecules in hippocampus induced by fibrillar Aβ42. Zhongguo Zhong Yao Za Zhi 2011, 36, 795–800. [Google Scholar] [PubMed]
- Ghasemi, R.; Zarifkar, A.; Rastegar, K.; Maghsoudi, N.; Moosavi, M. Repeated intra-hippocampal injection of β-amyloid 25–35 induces a reproducible impairment of learning and memory: Considering caspase-3 and mapks activity. Eur. J. Pharmacol. 2014, 726, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Du, H.; Yan, S.; Wu, X.; McKhann, G.M.; Chen, J.X.; Yan, S.S. Cyclophilin d deficiency rescues axonal mitochondrial transport in Alzheimer’s neurons. PLoS ONE 2013, 8, e54914. [Google Scholar] [CrossRef] [PubMed]
- Vina, J.; Lloret, A.; Valles, S.L.; Borras, C.; Badia, M.C.; Pallardo, F.V.; Sastre, J.; Alonso, M.D. Effect of gender on mitochondrial toxicity of Alzheimer’s Aβ peptide. Antioxid. Redox. Signal. 2007, 9, 1677–1690. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Wu, M.D.; Shaftel, S.S.; Kyrkanides, S.; LaFerla, F.M.; Olschowka, J.A.; O’Banion, M.K. Sustained interleukin-1β overexpression exacerbates tau pathology despite reduced amyloid burden in an Alzheimer’s mouse model. J. Neurosci. 2013, 33, 5053–5064. [Google Scholar] [CrossRef] [PubMed]
- Sha, D.; Li, L.; Ye, L.; Liu, R.; Xu, Y. Icariin inhibits neurotoxicity of β-amyloid by upregulating cocaine-regulated and amphetamine-regulated transcripts. Neuroreport 2009, 20, 1564–1567. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.Q.; Sun, X.B.; Xu, Y.X.; Zhao, H.; Zhu, Q.Y.; Zhu, C.Q. Astaxanthin upregulates heme oxygenase-1 expression through ERK1/2 pathway and its protective effect against β-amyloid-induced cytotoxicity in SH-SY5Y cells. Brain Res. 2010, 1360, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Wang, J.L.; Wang, Y.R.; Fa, X.Z. Apigenin attenuates copper-mediated β-amyloid neurotoxicity through antioxidation, mitochondrion protection and mapk signal inactivation in an ad cell model. Brain Res. 2013, 1492, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Wang, G.; Li, P.; Wang, Y.; Si, C.L.; He, J.; Long, W.; Bai, Y.; Feng, Z.; Wang, X. Neuroprotective effects of macranthoin g from eucommia ulmoides against hydrogen peroxide-induced apoptosis in PC12 cells via inhibiting NF-kappaB activation. Chem. Biol. Interact. 2014, 224, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Thangnipon, W.; Puangmalai, N.; Chinchalongporn, V.; Jantrachotechatchawan, C.; Kitiyanant, N.; Soi-Ampornkul, R.; Tuchinda, P.; Nobsathian, S. N-benzylcinnamide protects rat cultured cortical neurons from β-amyloid peptide-induced neurotoxicity. Neurosci. Lett. 2013, 556, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Lei, H.; Zhao, C.Y.; Liu, D.M.; Zhang, Y.; Li, L.; Wang, X.L.; Peng, Y. L-3-n-butylphthalide attenuates β-amyloid-induced toxicity in neuroblastoma SH-SY5Y cells through regulating mitochondrion-mediated apoptosis and MAPK signaling. J. Asian Nat. Prod. Res. 2014, 16, 854–864. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.N.; Xu, L.X.; Shan, L.; Zhang, W.D.; Li, H.L.; Wang, R. Neuroprotection of (+)-2-(1-hydroxyl-4-oxocyclohexyl) ethyl caffeate against hydrogen peroxide and lipopolysaccharide induced injury via modulating arachidonic acid network and p38-MAPK signaling. Curr. Alzheimer Res. 2015, 12, 892–902. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Yang, L.; Li, J. Protection against β-amyloid-induced neurotoxicity by naturally occurring z-ligustilide through the concurrent regulation of p38 and PI3-K/Akt pathways. Neurochem. Int. 2016, 100, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.K.; Choi, I.S.; Ban, J.O.; Lee, H.J.; Lee, U.S.; Han, S.B.; Jung, J.K.; Kim, Y.H.; Kim, K.H.; Oh, K.W.; et al. 4-O-methylhonokiol attenuated β-amyloid-induced memory impairment through reduction of oxidative damages via inactivation of p38 MAP kinase. J. Nutr. Biochem. 2011, 22, 476–486. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.I.; Lee, Y.K.; Park, S.G.; Choi, I.S.; Ban, J.O.; Park, H.K.; Nam, S.Y.; Yun, Y.W.; Han, S.B.; Oh, K.W.; et al. L-theanine, an amino acid in green tea, attenuates β-amyloid-induced cognitive dysfunction and neurotoxicity: Reduction in oxidative damage and inactivation of ERK/p38 kinase and NF-kappaB pathways. Free Radic. Biol. Med. 2009, 47, 1601–1610. [Google Scholar] [CrossRef] [PubMed]
- Arunsundar, M.; Shanmugarajan, T.S.; Ravichandran, V. 3,4-dihydroxyphenylethanol attenuates spatio-cognitive deficits in an Alzheimer’s disease mouse model: Modulation of the molecular signals in neuronal survival-apoptotic programs. Neurotox. Res. 2015, 27, 143–155. [Google Scholar] [CrossRef] [PubMed]
- Sabogal-Guaqueta, A.M.; Osorio, E.; Cardona-Gomez, G.P. Linalool reverses neuropathological and behavioral impairments in old triple transgenic Alzheimer’s mice. Neuropharmacology 2016, 102, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Wu, C.X.; Zhou, D.; Yang, F.; Tian, S.; Zhang, L.; Zhang, T.T.; Du, G.H. Pinocembrin protects against β-amyloid-induced toxicity in neurons through inhibiting receptor for advanced glycation end products (rage)-independent signaling pathways and regulating mitochondrion-mediated apoptosis. BMC Med. 2012, 10, 105. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Liu, Y.; Lao, M.; Ma, Z.; Yi, X. Puerarin protects Alzheimer’s disease neuronal cybrids from oxidant-stress induced apoptosis by inhibiting pro-death signaling pathways. Exp. Gerontol. 2011, 46, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Wu, Y.; Zha, S.; Liu, M.; Wang, Y.; Yang, G.; Ma, K.; Fei, Y.; Zhang, Y.; Hu, X.; et al. Treatment effects of tanshinone iia against intracerebroventricular streptozotocin induced memory deficits in mice. Brain Res. 2016, 1631, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Zou, L.; Qin, H.; He, Y.; Huang, H.; Lu, Y.; Chu, X. Inhibiting p38 mitogen-activated protein kinase attenuates cerebral ischemic injury in swedish mutant amyloid precursor protein transgenic mice. Neural Regen. Res. 2012, 7, 1088–1094. [Google Scholar] [PubMed]
- Lai, A.Y.; McLaurin, J. Clearance of amyloid-β peptides by microglia and macrophages: The issue of what, when and where. Future Neurol. 2012, 7, 165–176. [Google Scholar] [CrossRef] [PubMed]
- Cai, Z.; Hussain, M.D.; Yan, L.J. Microglia, neuroinflammation, and β-amyloid protein in Alzheimer’s disease. Int. J. Neurosci. 2014, 124, 307–321. [Google Scholar] [CrossRef] [PubMed]
- Heppner, F.L.; Ransohoff, R.M.; Becher, B. Immune attack: The role of inflammation in Alzheimer disease. Nat. Rev. Neurosci. 2015, 16, 358–372. [Google Scholar] [CrossRef] [PubMed]
- Varnum, M.M.; Kiyota, T.; Ingraham, K.L.; Ikezu, S.; Ikezu, T. The anti-inflammatory glycoprotein, CD200, restores neurogenesis and enhances amyloid phagocytosis in a mouse model of Alzheimer’s disease. Neurobiol. Aging 2015, 36, 2995–3007. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Rauvala, H.; Gahmberg, C.G. Neuronal regulation of immune responses in the central nervous system. Trends Immunol. 2009, 30, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Perry, V.H.; Holmes, C. Microglial priming in neurodegenerative disease. Nat. Rev. Neurol. 2014, 10, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Heneka, M.T.; Kummer, M.P.; Stutz, A.; Delekate, A.; Schwartz, S.; Vieira-Saecker, A.; Griep, A.; Axt, D.; Remus, A.; Tzeng, T.C.; et al. Nlrp3 is activated in Alzheimer’s disease and contributes to pathology in APP/PS1 mice. Nature 2013, 493, 674–678. [Google Scholar] [CrossRef] [PubMed]
- Paradisi, S.; Sacchetti, B.; Balduzzi, M.; Gaudi, S.; Malchiodi-Albedi, F. Astrocyte modulation of in vitro β-amyloid neurotoxicity. Glia 2004, 46, 252–260. [Google Scholar] [CrossRef] [PubMed]
- Thal, D.R. The role of astrocytes in amyloid β-protein toxicity and clearance. Exp. Neurol. 2012, 236, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Bhat, N.R.; Zhang, P.S.; Lee, J.C.; Hogan, E.L. Extracellular signal-regulated kinase and p38 subgroups of mitogen-activated protein kinases regulate inducible nitric oxide synthase and tumor necrosis factor-α gene expression in endotoxin-stimulated primary glial cultures. J. Neurosci. 1998, 18, 1633–1641. [Google Scholar] [PubMed]
- Lee, Y.B.; Schrader, J.W.; Kim, S.U. P38 map kinase regulates tnf-α production in human astrocytes and microglia by multiple mechanisms. Cytokine 2000, 12, 874–880. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Smith, C.J.; Van Eldik, L.J. Importance of mapk pathways for microglial pro-inflammatory cytokine IL-1β production. Neurobiol. Aging 2004, 25, 431–439. [Google Scholar] [CrossRef]
- McDonald, D.R.; Bamberger, M.E.; Combs, C.K.; Landreth, G.E. Β-amyloid fibrils activate parallel mitogen-activated protein kinase pathways in microglia and thp1 monocytes. J. Neurosci. 1998, 18, 4451–4460. [Google Scholar] [PubMed]
- Giovannini, M.G.; Scali, C.; Prosperi, C.; Bellucci, A.; Vannucchi, M.G.; Rosi, S.; Pepeu, G.; Casamenti, F. Β-amyloid-induced inflammation and cholinergic hypofunction in the rat brain in vivo: Involvement of the p38-MAPK pathway. Neurobiol. Dis. 2002, 11, 257–274. [Google Scholar] [CrossRef] [PubMed]
- Moynagh, P.N. The interleukin-1 signalling pathway in astrocytes: A key contributor to inflammation in the brain. J. Anat. 2005, 207, 265–269. [Google Scholar] [CrossRef] [PubMed]
- DaSilva, J.; Pierrat, B.; Mary, J.L.; Lesslauer, W. Blockade of p38 mitogen-activated protein kinase pathway inhibits inducible nitric-oxide synthase expression in mouse astrocytes. J. Biol. Chem. 1997, 272, 28373–28380. [Google Scholar] [CrossRef]
- Hua, L.W.L.; Zhao, M.L.; Cosenza, M.; Kim, M.O.; Huang, H.; Tanowitz, H.B.; Brosnan, C.F.; Lee, S.C. Role of mitogen-activated protein kinases in inducible nitric oxide synthase and tnf α expression in human fetal astrocytes. J. Neuroimmunol. 2002, 126, 180–189. [Google Scholar] [CrossRef]
- Bhat, N.R.; Feinstein, D.L.; Shen, Q.; Bhat, A.N. P38 MAPK-mediated transcriptional activation of inducible nitric-oxide synthase in glial cells—Roles of nuclear factors, nuclear factor kappaB, cAMP response element-binding protein, CCAAT/enhancer-binding protein-β, and activating transcription factor-2. J. Biol. Chem. 2002, 277, 29584–29592. [Google Scholar] [CrossRef] [PubMed]
- Saha, R.N.; Jana, M.; Pahan, K. Mapk p38 regulates transcriptional activity of NF-kappaB in primary human astrocytes via acetylation of p65. J. Immunol. 2007, 179, 7101–7109. [Google Scholar] [CrossRef] [PubMed]
- Munoz, L.; Ranaivo, H.R.; Roy, S.M.; Hu, W.; Craft, J.M.; McNamara, L.K.; Chico, L.W.; Van Eldik, L.J.; Watterson, D.M. Novel p38α MAPK inhibitor suppresses brain proinflammatory cytokine up-regulation and attenuates synaptic dysfunction and behavioral deficits in an Alzheimer’s disease mouse model. J. Neuroinflamm. 2007, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, M.; Kwon, B.M.; Suk, K.; McGeer, E.; McGeer, P.L. Effects of obovatol on GSH depleted glia-mediated neurotoxicity and oxidative damage. J. Neuroimmune Pharmacol. 2012, 7, 173–186. [Google Scholar] [CrossRef] [PubMed]
- Gan, P.; Zhang, L.; Chen, Y.; Zhang, Y.; Zhang, F.; Zhou, X.; Zhang, X.; Gao, B.; Zhen, X.; Zhang, J.; et al. Anti-inflammatory effects of glaucocalyxin b in microglia cells. J. Pharmacol. Sci. 2015, 128, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Park, S.J.; Park, N.J.; Joo, W.H.; Lee, S.J.; Choi, Y.W. Alpha-iso-cubebene exerts neuroprotective effects in amyloid β stimulated microglia activation. Neurosci. Lett. 2013, 555, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Li, Y.X.; Dewapriya, P.; Ryu, B.; Kim, S.K. Floridoside suppresses pro-inflammatory responses by blocking MAPK signaling in activated microglia. BMB Rep. 2013, 46, 398–403. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; McGeer, E.; Kodela, R.; Kashfi, K.; McGeer, P.L. Nosh-aspirin (NBS-1120), a novel nitric oxide and hydrogen sulfide releasing hybrid, attenuates neuroinflammation induced by microglial and astrocytic activation: A new candidate for treatment of neurodegenerative disorders. Glia 2013, 61, 1724–1734. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.G.; Moon, M.; Choi, J.G.; Park, G.; Kim, A.J.; Hur, J.; Lee, K.T.; Oh, M.S. Donepezil inhibits the amyloid-β oligomer-induced microglial activation in vitro and in vivo. Neurotoxicology 2014, 40, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Wang, S.; Yu, L.; Zhu, X.; Xu, Y. Esculentoside a suppresses β(1-42)-induced neuroinflammation by down-regulating mapks pathways in vivo. Neurol. Res. 2015, 37, 859–866. [Google Scholar] [CrossRef] [PubMed]
- Walsh, D.M.; Klyubin, I.; Fadeeva, J.V.; Cullen, W.K.; Anwyl, R.; Wolfe, M.S.; Rowan, M.J.; Selkoe, D.J. Naturally secreted oligomers of amyloid β protein potently inhibit hippocampal long-term potentiation in vivo. Nature 2002, 416, 535–539. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.W.; Rowan, M.J.; Anwyl, R. Inhibition of LTP by β-amyloid is prevented by activation of β2 adrenoceptors and stimulation of the cAMP/Pka signalling pathway. Neurobiol. Aging 2009, 30, 1608–1613. [Google Scholar] [CrossRef] [PubMed]
- Bolshakov, V.Y.; Carboni, L.; Cobb, M.H.; Siegelbaum, S.A.; Belardetti, F. Dual MAP kinase pathways mediate opposing forms of long-term plasticity at CA3-CA1 synapses. Nat. Neurosci. 2000, 3, 1107–1112. [Google Scholar] [PubMed]
- Rush, A.M.; Wu, J.; Rowan, M.J.; Anwyl, R. Group I metabotropic glutamate receptor (mGluR)-dependent long-term depression mediated via p38 mitogen-activated protein kinase is inhibited by previous high-frequency stimulation and activation of mGluRs and protein kinase C in the rat dentate gyrus in vitro. J. Neurosci. 2002, 22, 6121–6128. [Google Scholar] [PubMed]
- Izumi, Y.; Tokuda, K.; Zorumski, C.F. Long-term potentiation inhibition by low-level N-methyl-d-aspartate receptor activation involves calcineurin, nitric oxide, and p38 mitogen-activated protein kinase. Hippocampus 2008, 18, 258–265. [Google Scholar] [CrossRef] [PubMed]
- Um, J.W.; Kaufman, A.C.; Kostylev, M.; Heiss, J.K.; Stagi, M.; Takahashi, H.; Kerrisk, M.E.; Vortmeyer, A.; Wisniewski, T.; Koleske, A.J.; et al. Metabotropic glutamate receptor 5 is a coreceptor for Alzheimer Aβ oligomer bound to cellular prion protein. Neuron 2013, 79, 887–902. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, A.; Esseltine, J.L.; DeVries, R.A.; Cregan, S.P.; Ferguson, S.S. Metabotropic glutamate receptor 5 knockout reduces cognitive impairment and pathogenesis in a mouse model of Alzheimer’s disease. Mol. Brain 2014, 7, 40. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Lin, R.; Chang, L.; Xu, S.; Wei, X.; Zhang, J.; Wang, C.; Anwyl, R.; Wang, Q. Enhancement of long-term depression by soluble amyloid β protein in rat hippocampus is mediated by metabotropic glutamate receptor and involves activation of p38 MAPK, step and caspase-3. Neuroscience 2013, 253, 435–443. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, H.; Boehm, J.; Sato, C.; Iwatsubo, T.; Tomita, T.; Sisodia, S.; Malinow, R. Ampar removal underlies Aβ-induced synaptic depression and dendritic spine loss. Neuron 2006, 52, 831–843. [Google Scholar] [CrossRef] [PubMed]
- Shankar, G.M.; Walsh, D.M. Alzheimer’s disease: Synaptic dysfunction and Aβ. Mol. Neurodegener. 2009, 4, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nomura, I.; Takechi, H.; Kato, N. Intraneuronally injected amyloid β inhibits long-term potentiation in rat hippocampal slices. J. Neurophysiol. 2012, 107, 2526–2531. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Jin, M.; Koeglsperger, T.; Shepardson, N.E.; Shankar, G.M.; Selkoe, D.J. Soluble Aβ oligomers inhibit long-term potentiation through a mechanism involving excessive activation of extrasynaptic NR2b-containing nmda receptors. J. Neurosci. 2011, 31, 6627–6638. [Google Scholar] [CrossRef] [PubMed]
- Mondragon-Rodriguez, S.; Trillaud-Doppia, E.; Dudilot, A.; Bourgeois, C.; Lauzon, M.; Leclerc, N.; Boehm, J. Interaction of endogenous tau protein with synaptic proteins is regulated by N-methyl-d-aspartate receptor-dependent tau phosphorylation. J. Biol.Chem. 2012, 287, 32040–32053. [Google Scholar] [CrossRef] [PubMed]
- Regan, P.; Whitcomb, D.J.; Cho, K. Physiological and pathophysiological implications of synaptic tau. Neuroscientist 2017, 23, 137–151. [Google Scholar] [CrossRef] [PubMed]
- Kimura, T.; Whitcomb, D.J.; Jo, J.; Regan, P.; Piers, T.; Heo, S.; Brown, C.; Hashikawa, T.; Murayama, M.; Seok, H.; et al. Microtubule-associated protein tau is essential for long-term depression in the hippocampus. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2014, 369, 20130144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Regan, P.; Piers, T.; Yi, J.H.; Kim, D.H.; Huh, S.; Park, S.J.; Ryu, J.H.; Whitcomb, D.J.; Cho, K. Tau phosphorylation at serine 396 residue is required for hippocampal ltd. J. Neurosci. 2015, 35, 4804–4812. [Google Scholar] [CrossRef] [PubMed]
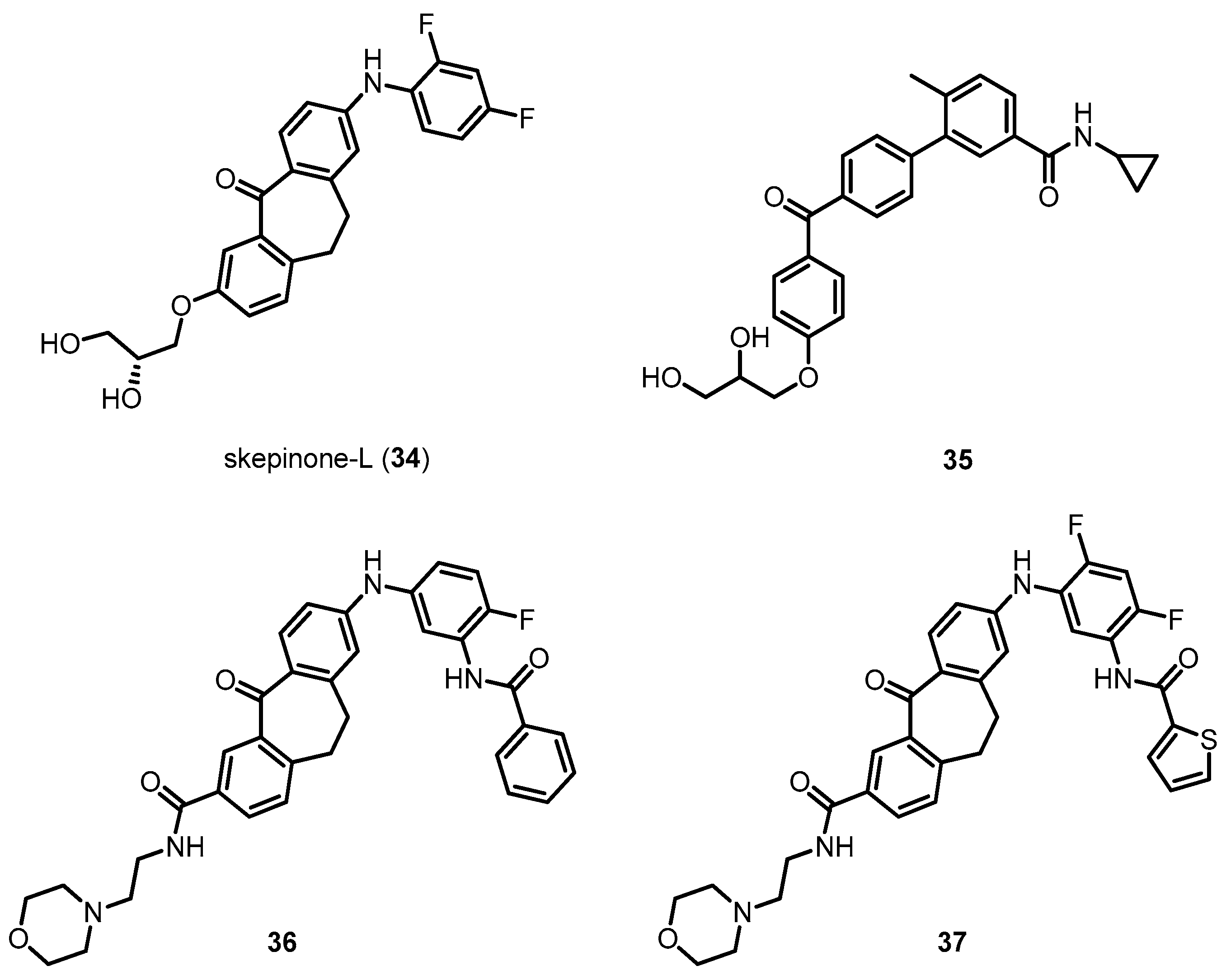
- Koeberle, S.C.; Romir, J.; Fischer, S.; Koeberle, A.; Schattel, V.; Albrecht, W.; Grutter, C.; Werz, O.; Rauh, D.; Stehle, T.; et al. Skepinone-l is a selective p38 mitogen-activated protein kinase inhibitor. Nat. Chem. Biol. 2012, 8, 141–143. [Google Scholar] [CrossRef] [PubMed]
- Heo, J.; Shin, H.; Lee, J.; Kim, T.; Inn, K.S.; Kim, N.J. Synthesis and biological evaluation of N-cyclopropylbenzamide-benzophenone hybrids as novel and selective p38 mitogen activated protein kinase (MAPK) inhibitors. Bioorg. Med. Chem. Lett. 2015, 25, 3694–3698. [Google Scholar] [CrossRef] [PubMed]
- Koeberle, S.C.; Fischer, S.; Schollmeyer, D.; Schattel, V.; Grutter, C.; Rauh, D.; Laufer, S.A. Design, synthesis, and biological evaluation of novel disubstituted dibenzosuberones as highly potent and selective inhibitors of p38 mitogen activated protein kinase. J. Med. Chem. 2012, 55, 5868–5877. [Google Scholar] [CrossRef] [PubMed]
- Martz, K.E.; Dorn, A.; Baur, B.; Schattel, V.; Goettert, M.I.; Mayer-Wrangowski, S.C.; Rauh, D.; Laufer, S.A. Targeting the hinge glycine flip and the activation loop: Novel approach to potent p38 α inhibitors. J. Med. Chem. 2012, 55, 7862–7874. [Google Scholar] [CrossRef] [PubMed]
- Baur, B.; Storch, K.; Martz, K.E.; Goettert, M.I.; Richters, A.; Rauh, D.; Laufer, S.A. Metabolically stable dibenzo[b,e]oxepin-11(6H)-ones as highly selective p38 map kinase inhibitors: Optimizing anti-cytokine activity in human whole blood. J. Med. Chem. 2013, 56, 8561–8578. [Google Scholar] [CrossRef] [PubMed]
- Fischer, S.; Wentsch, H.K.; Mayer-Wrangowski, S.C.; Zimmermann, M.; Bauer, S.M.; Storch, K.; Niess, R.; Koeberle, S.C.; Grutter, C.; Boeckler, F.M.; et al. Dibenzosuberones as p38 mitogen-activated protein kinase inhibitors with low ATP competitiveness and outstanding whole blood activity. J. Med. Chem. 2013, 56, 241–253. [Google Scholar] [CrossRef] [PubMed]
- Wentsch, H.K.; Walter, N.M.; Buhrmann, M.; Mayer-Wrangowski, S.; Rauh, D.; Zaman, G.J.R.; Willemsen-Seegers, N.; Buijsman, R.C.; Henning, M.; Dauch, D.; et al. Optimized target residence time: Type I1/2 inhibitors for p38 MAP kinase with improved binding kinetics through direct interaction with the R-spine. Angew. Chem. Int. Ed. 2017, 56, 5363–5367. [Google Scholar] [CrossRef] [PubMed]
- Maphis, N.; Jiang, S.Y.; Xu, G.X.; Kokiko-Cochran, O.N.; Roy, S.M.; Van Eldik, L.J.; Watterson, D.M.; Lamb, B.T.; Bhaskar, K. Selective suppression of the α isoform of p38 MAPK rescues late-stage tau pathology. Alzheimers Res. Ther. 2016, 8. [Google Scholar] [CrossRef] [PubMed]
- Ittner, A.; Chua, S.W.; Bertz, J.; Volkerling, A.; van der Hoven, J.; Gladbach, A.; Przybyla, M.; Bi, M.; van Hummel, A.; Stevens, C.H.; et al. Site-specific phosphorylation of tau inhibits amyloid-β toxicity in Alzheimer’s mice. Science 2016, 354, 904–908. [Google Scholar] [CrossRef] [PubMed]
- Joosen, M.J.A.; Vester, S.M.; Hamelink, J.; Klaassen, S.D.; van den Berg, R.M. Increasing nerve agent treatment efficacy by P-glycoprotein inhibition. Chem. Biol. Interact. 2016, 259, 115–121. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | M.W. 1 | Mode of Action | Activities | Reference |
|---|---|---|---|---|
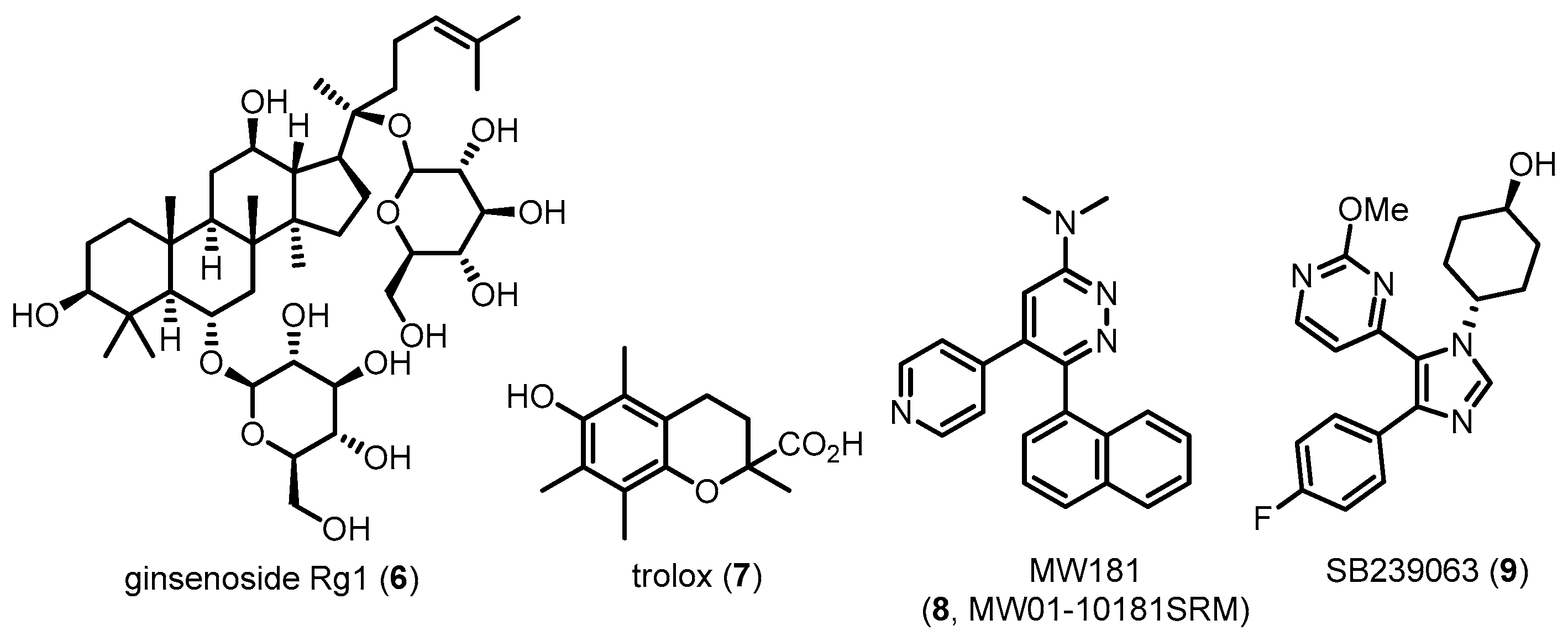
| Ginsenoside Rg1 (6) | 801.01 | Inhibiting p38 MAPK activation | Attenuating tau hyperphosphorylation | [75] |
| Trolox (7) | 250.29 | Inhibiting p38 MAPK activation | Decreasing tau toxicities | [76] |
| MW181 (8) | 326.39 | Directly inhibiting p38 MAPK | Attenuating tau hyperphosphorylation/Preventing cognitive impairments | [78,163] |
| SB239063 (9) | 368.40 | Directly inhibiting p38 MAPK | Attenuating tau hyperphosphorylation/Preventing cognitive impairments | [78] |
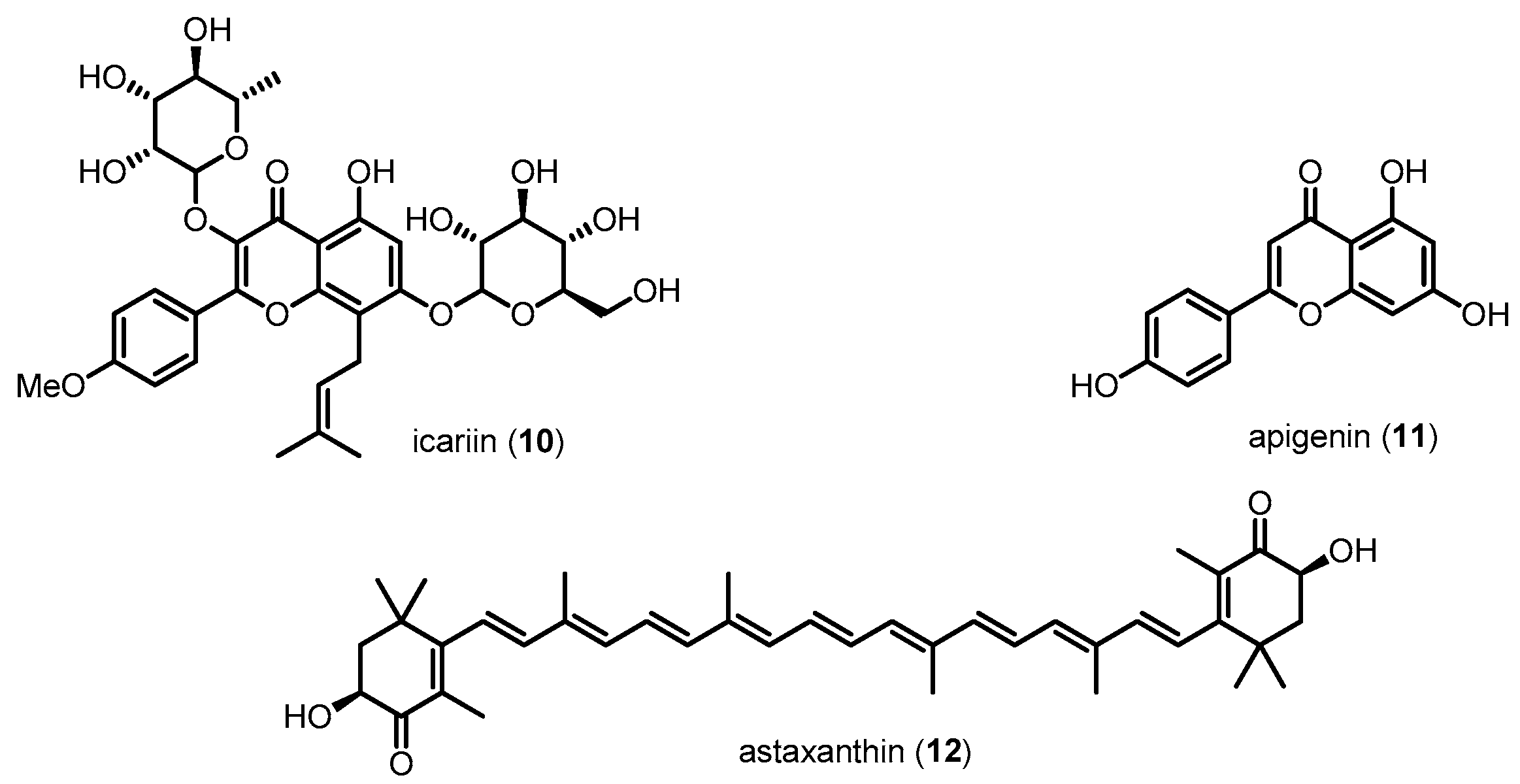
| Icarin (10) | 676.66 | Inhibiting p38 MAPK activation | Reducing Aβ-induced neurotoxicity | [97] |
| Apigenin (11) | 270.24 | Inhibiting p38 MAPK activation | Reducing Aβ-induced neurotoxicity | [99] |
| Astaxanthin (12) | 596.84 | Inhibiting p38 MAPK activation | Reducing Aβ-induced neurotoxicity | [98] |
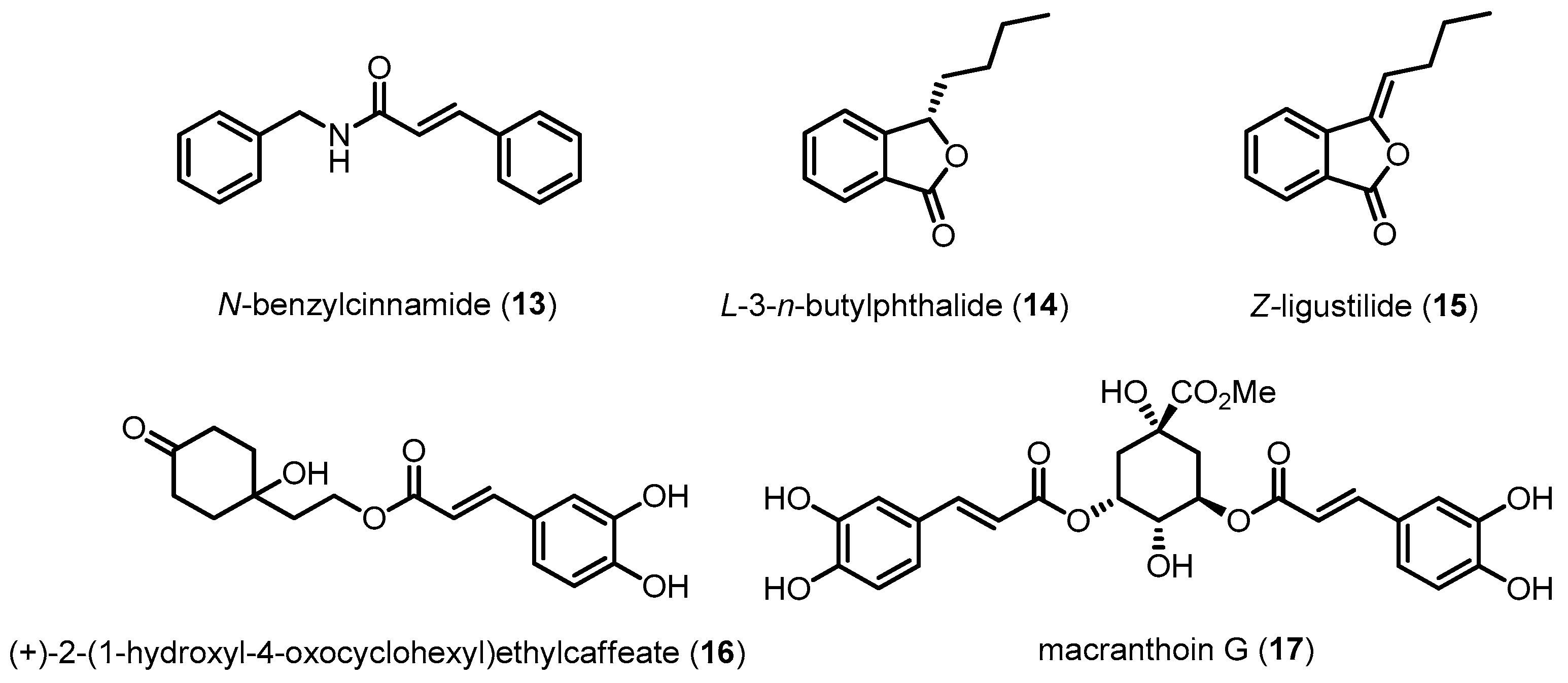
| N-Benzylcinnamide (13) | 237.30 | Inhibiting p38 MAPK activation | Reducing Aβ-induced neurotoxicity | [101] |
| l-3-n-Butylphthalide (14) | 190.24 | Inhibiting p38 MAPK activation | Reducing Aβ-induced neurotoxicity | [102] |
| z-Ligustilide (15) | 188.22 | Inhibiting p38 MAPK activation | Reducing Aβ-induced neurotoxicity | [104] |
| (+)-2-(1-Hydroxyl-4-oxocyclohexyl) ethylcaffeate (16) | 320.34 | Inhibiting p38 MAPK activation | Reducing H2O2-induced neurotoxicity | [103] |
| Macranthoin G (17) | 530.48 | Inhibiting p38 MAPK activation | Reducing H2O2-induced neurotoxicity | [100] |
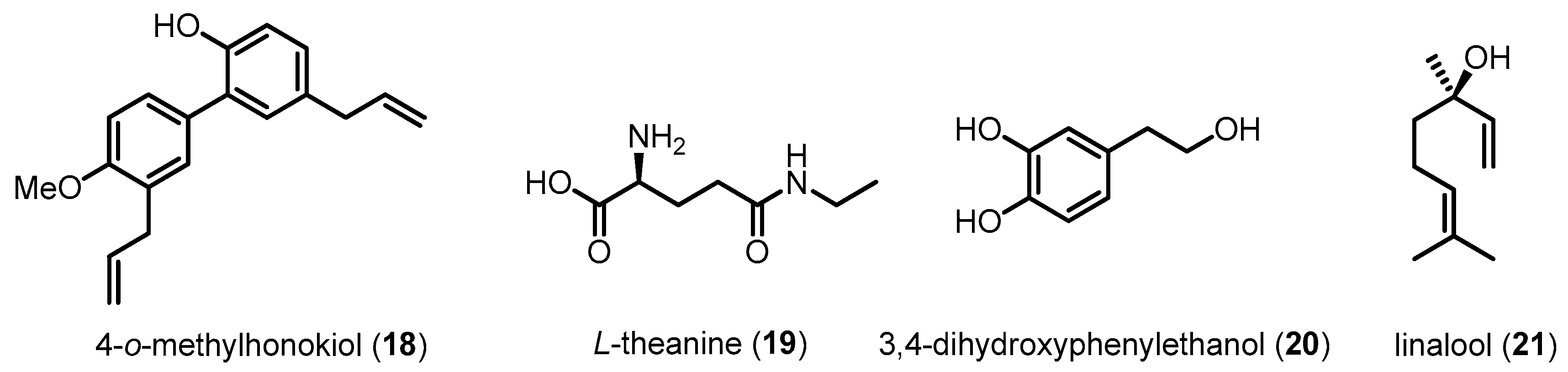
| 4-O-methylhonokiol (18) | 280.36 | Inhibiting p38 MAPK activation | Reducing Aβ-induced neurotoxicity and neuroinflammation/Preventing memory impairment | [105] |
| l-Theanine (19) | 160.17 | Inhibiting p38 MAPK activation | Reducing Aβ-induced neurotoxicity/Preventing memory impairment | [106] |
| 3,4-Dihydroxyphenyl ethanol (20) | 154.16 | Inhibiting p38 MAPK activation | Reducing Aβ-induced neurotoxicity/Preventing memory impairment | [107] |
| Linalool (21) | 154.25 | Inhibiting p38 MAPK activation | Attenuating tau hyperphosphorylation/Improving learning and spatial memory/Reducing neuroinflammation | [108] |
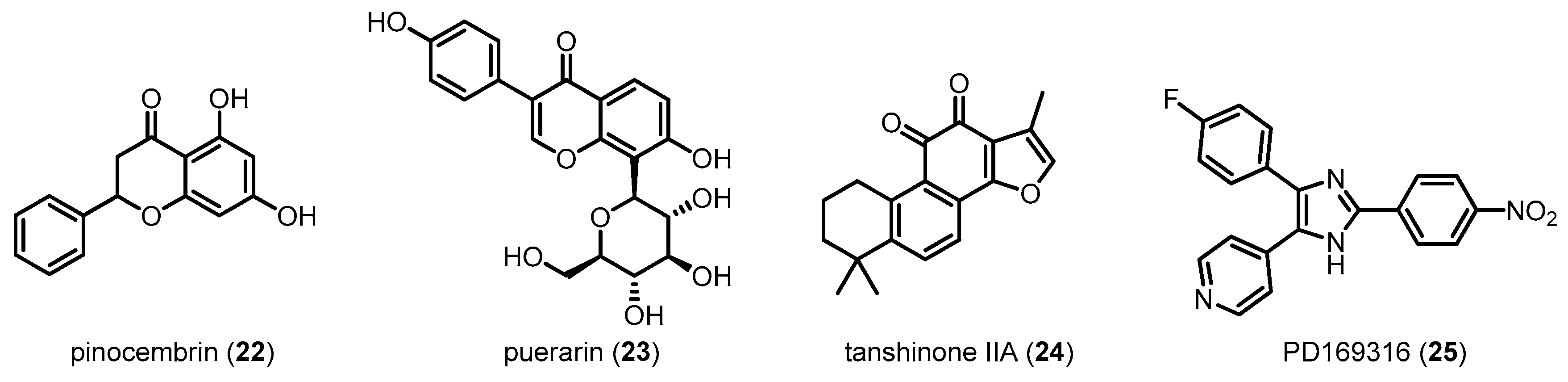
| Pinocembrin (22) | 256.25 | Inhibiting p38 MAPK | Reducing Aβ-induced neurotoxicity/Improving behavioral performance | [109] |
| Puerarin (23) | 416.38 | Inhibiting p38 MAPK | Alleviating mitochondrial dysfunction | [110] |
| Tanshinone IIA (24) | 294.34 | Inhibiting p38 MAPK | Preventing memory impairment | [111] |
| PD169316 (25) | 360.34 | Directly inhibiting p38 MAPK | Reducing Aβ-induced neurotoxicity | [28] |
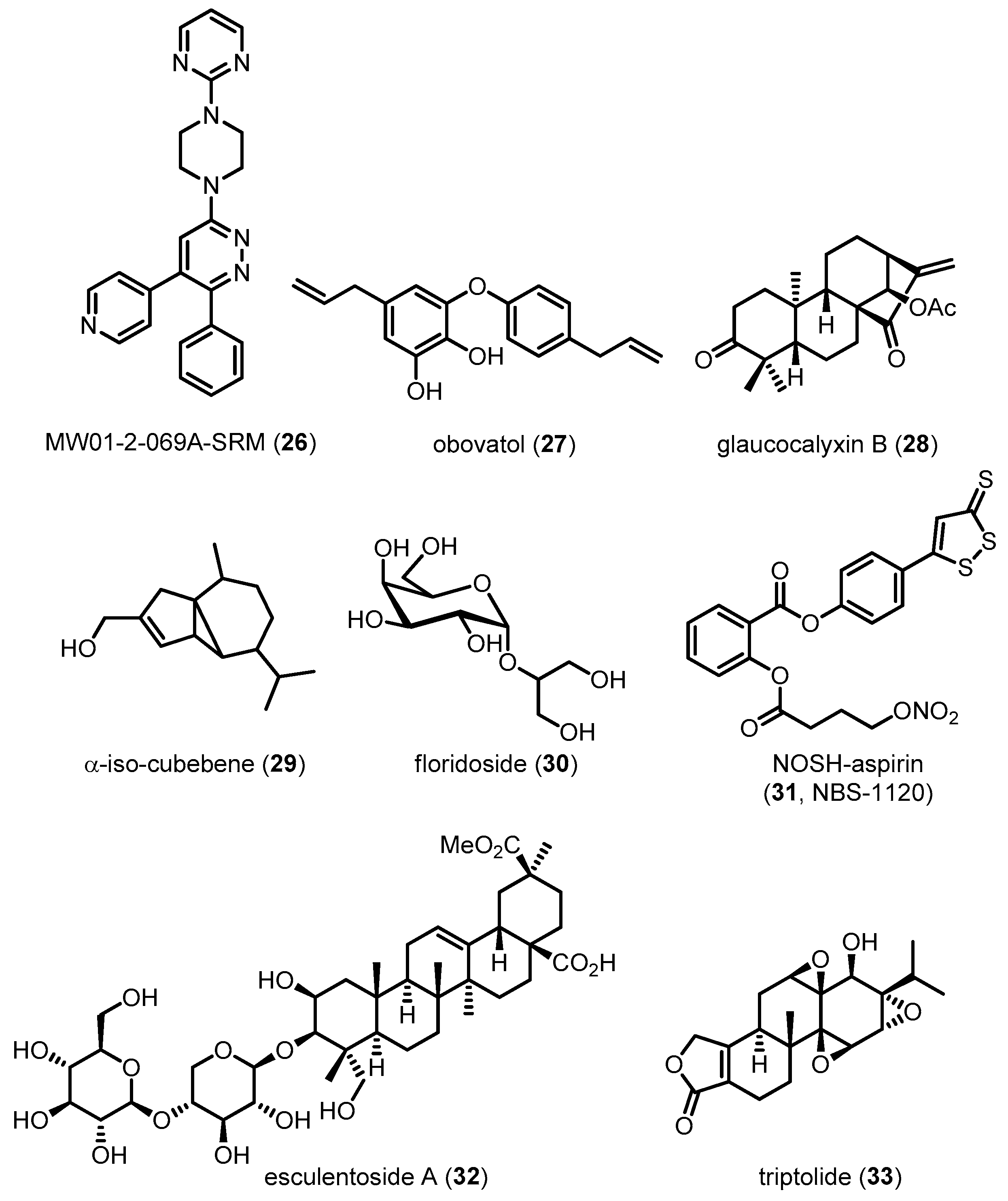
| MW01-2-069A-SRM (26) | 395.46 | Directly inhibiting p38 MAPK | Reducing neuroinflammation/Improving behavioral performance | [132] |
| Obovatol (27) | 282.33 | Inhibiting p38 MAPK activation | Reducing neuroinflammation | [133] |
| Glaucocalyxin B (28) | 358.47 | Inhibiting p38 MAPK activation | Reducing neuroinflammation | [134] |
| α-iso-cubebene (29) | 220.35 | Inhibiting p38 MAPK activation | Reducing neuroinflammation | [135] |
| Floridoside (30) | 254.23 | Inhibiting p38 MAPK activation | Reducing neuroinflammation | [136] |
| NOSH-aspirin (31, NBS-1120) | 461.47 | Inhibiting p38 MAPK activation | Reducing neuroinflammation | [137] |
| Esculentoside A (32) | 826.96 | Inhibiting p38 MAPK activation | Reducing neuroinflammation/Improving learning and spatial memory | [139] |
| Triptolide (33) | 360.40 | Inhibiting p38 MAPK activation | Reducing neuroinflammation/Improving learning and spatial memory | [91] |
| Skepinone-l (34) | 425.42 | Directly inhibiting p38 MAPK | ND 2 | [156] |
| Compound 35 | 445.51 | Directly inhibiting p38 MAPK | ND 2 | [157] |
| Compound 36 | 592.66 | Directly inhibiting p38 MAPK | ND 2 | [162] |
| Compound 37 | 616.68 | Directly inhibiting p38 MAPK | ND 2 | [162] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.K.; Kim, N.-J. Recent Advances in the Inhibition of p38 MAPK as a Potential Strategy for the Treatment of Alzheimer’s Disease. Molecules 2017, 22, 1287. https://doi.org/10.3390/molecules22081287
Lee JK, Kim N-J. Recent Advances in the Inhibition of p38 MAPK as a Potential Strategy for the Treatment of Alzheimer’s Disease. Molecules. 2017; 22(8):1287. https://doi.org/10.3390/molecules22081287
Chicago/Turabian StyleLee, Jong Kil, and Nam-Jung Kim. 2017. "Recent Advances in the Inhibition of p38 MAPK as a Potential Strategy for the Treatment of Alzheimer’s Disease" Molecules 22, no. 8: 1287. https://doi.org/10.3390/molecules22081287
APA StyleLee, J. K., & Kim, N. -J. (2017). Recent Advances in the Inhibition of p38 MAPK as a Potential Strategy for the Treatment of Alzheimer’s Disease. Molecules, 22(8), 1287. https://doi.org/10.3390/molecules22081287