A Pectic Polysaccharide from Sijunzi Decoction Promotes the Antioxidant Defenses of SW480 Cells

 , , and
, , and
Abstract
:1. Introduction
2. Results and Discussion
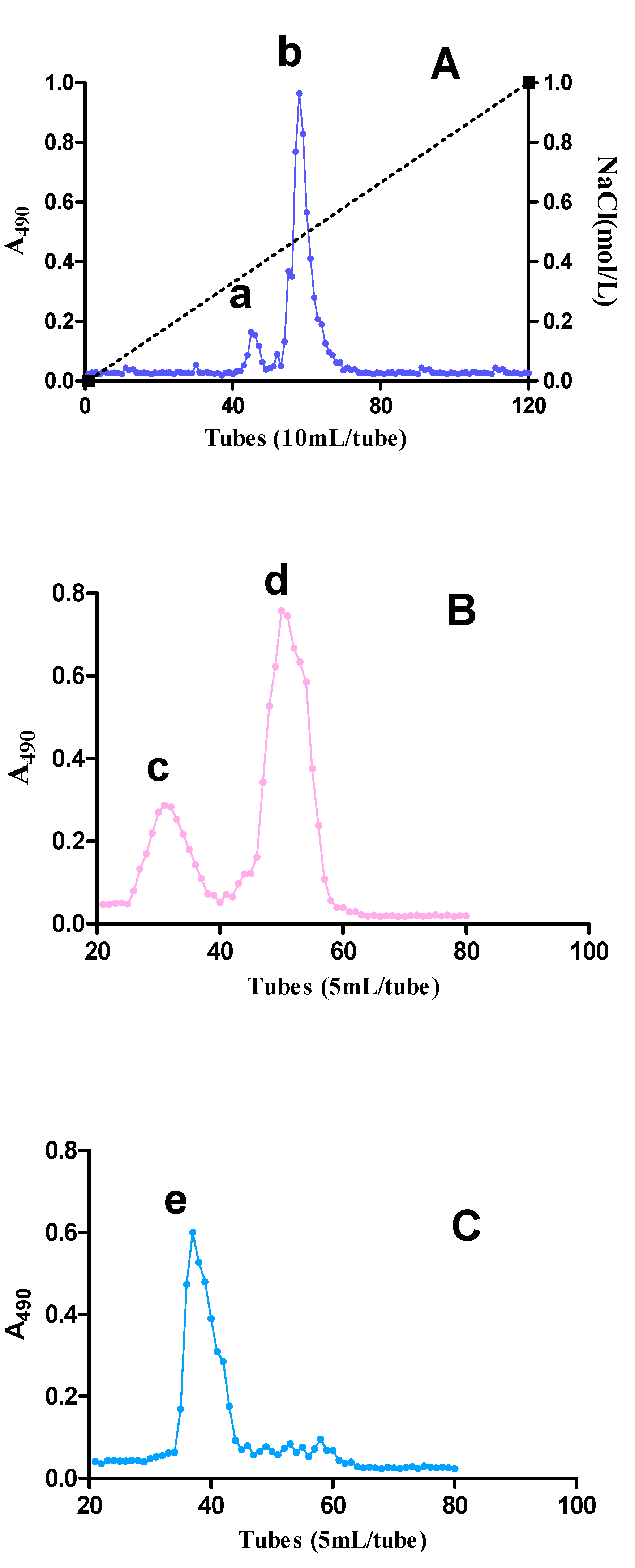
2.1. Fractionation of Pectic Polysaccharides from SJZDP
2.2. Chemical Composition of Polysaccharide
2.3. Determination of the Glycosidic Linkages
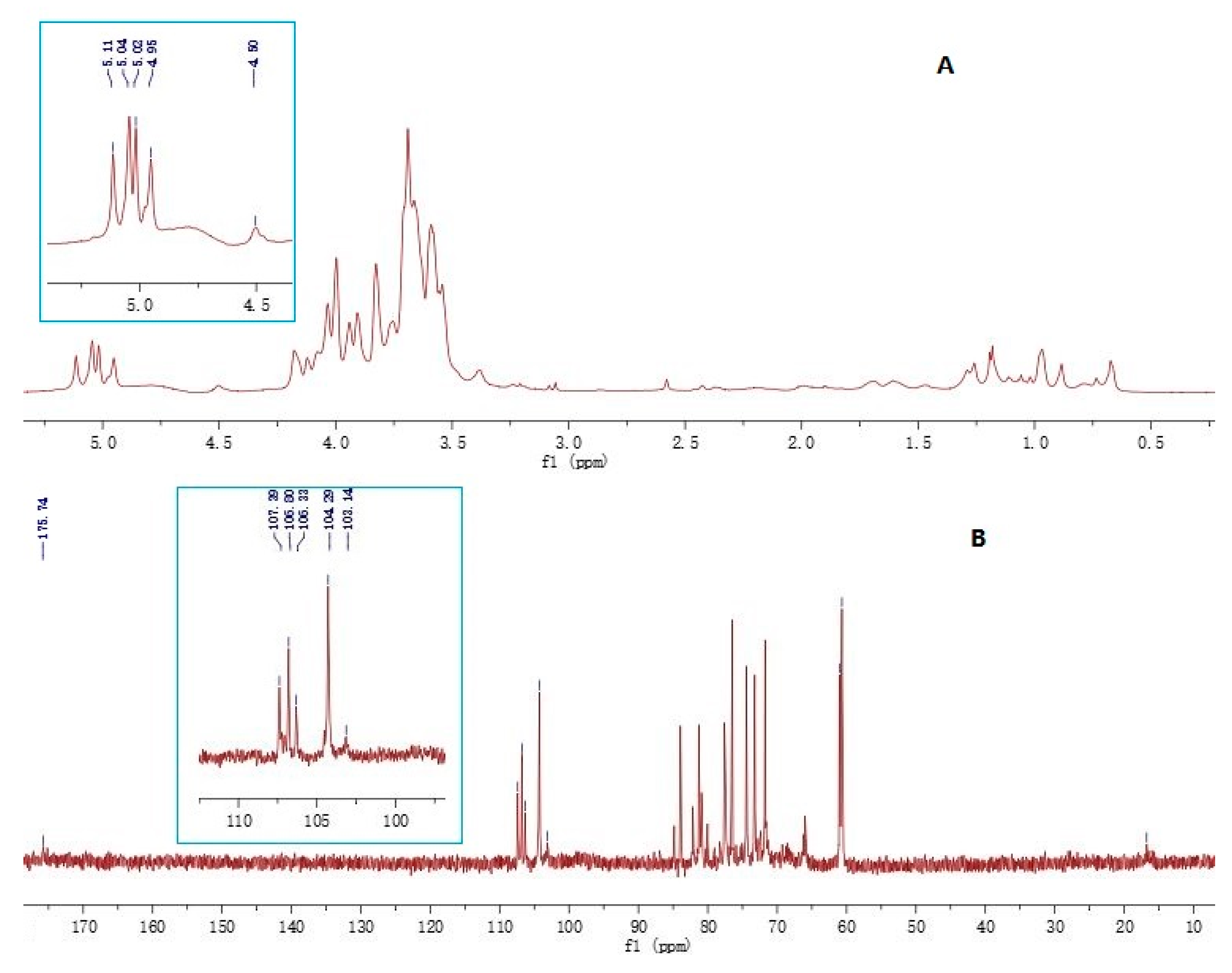
2.4. Structural Features
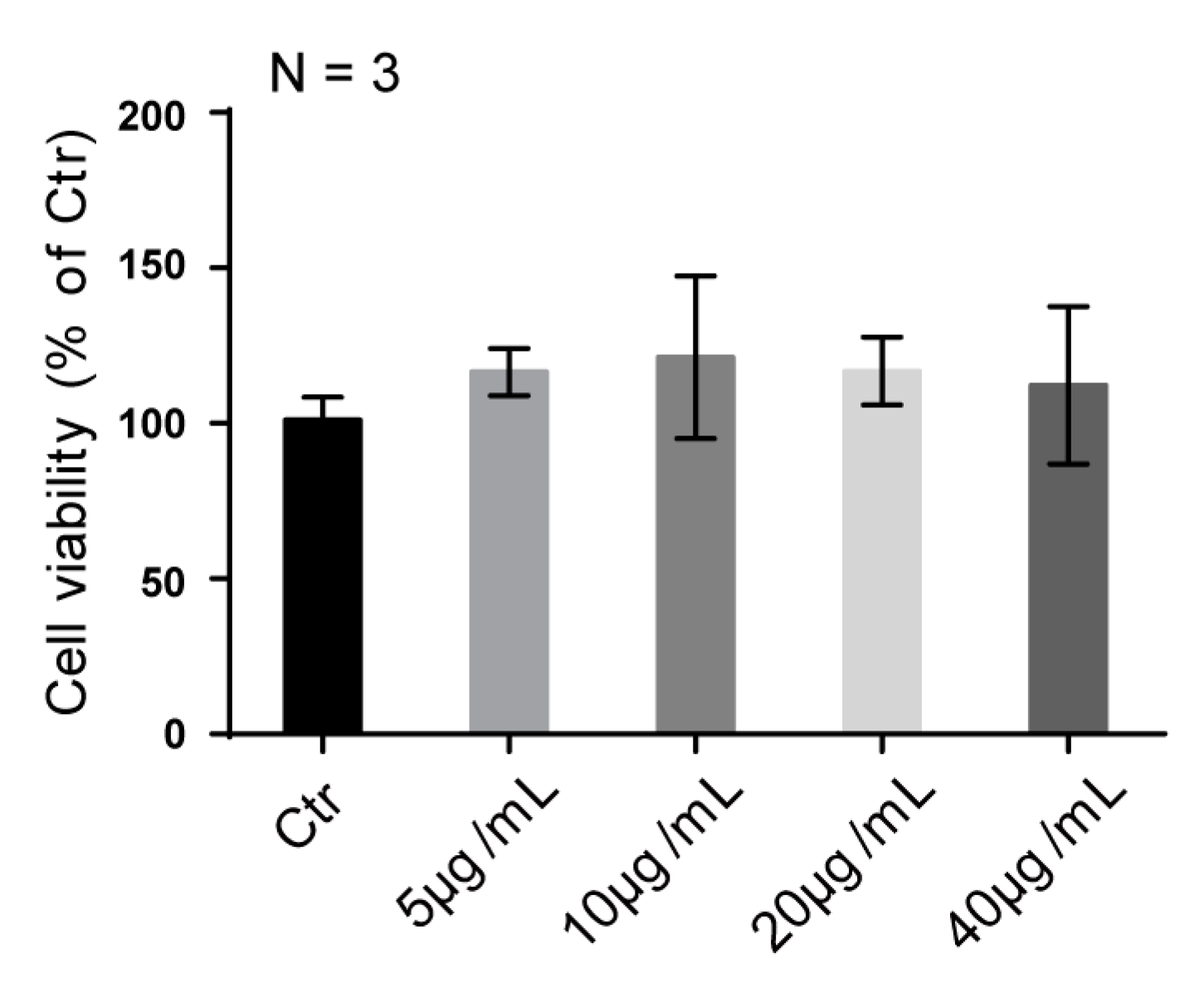
2.5. SJZDP-II-I Treatment Didn’t Affect the Proliferation of SW480 Cells
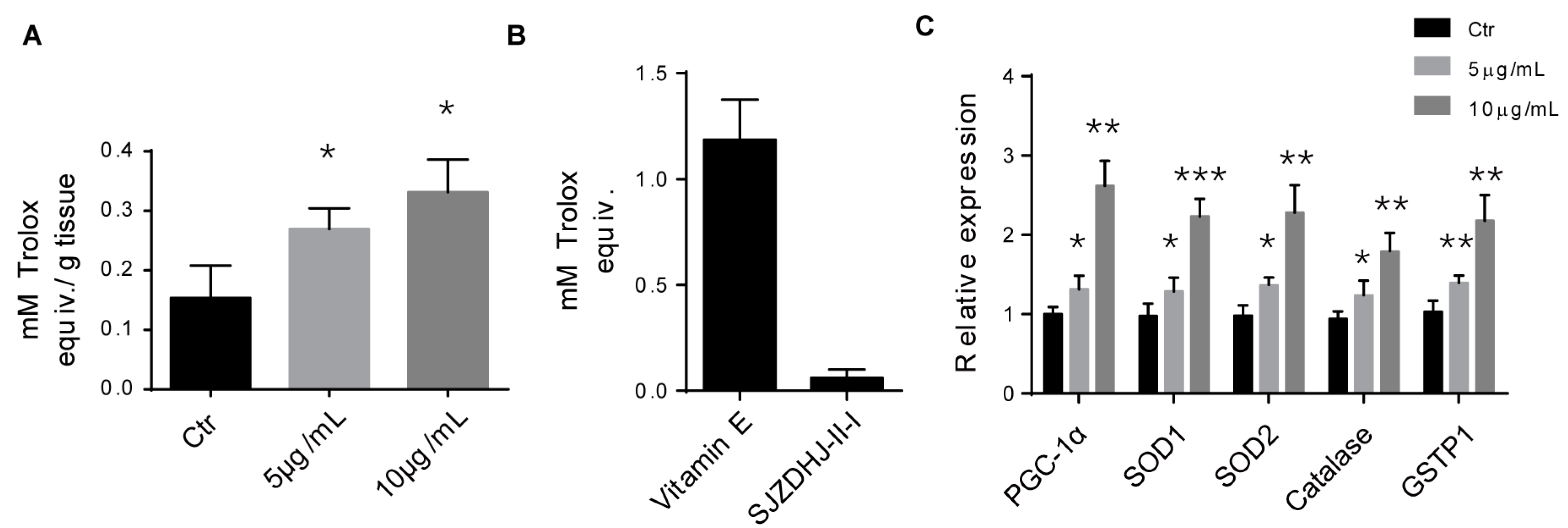
2.6. SJZDP-II-I Promoted the Antioxidant Defense of SW480 Cells
3. Materials and Methods
3.1. Preparation of Polysaccharides
3.2. Chemical Compositions and Linkage Determination
3.3. Molecular Weight Determination
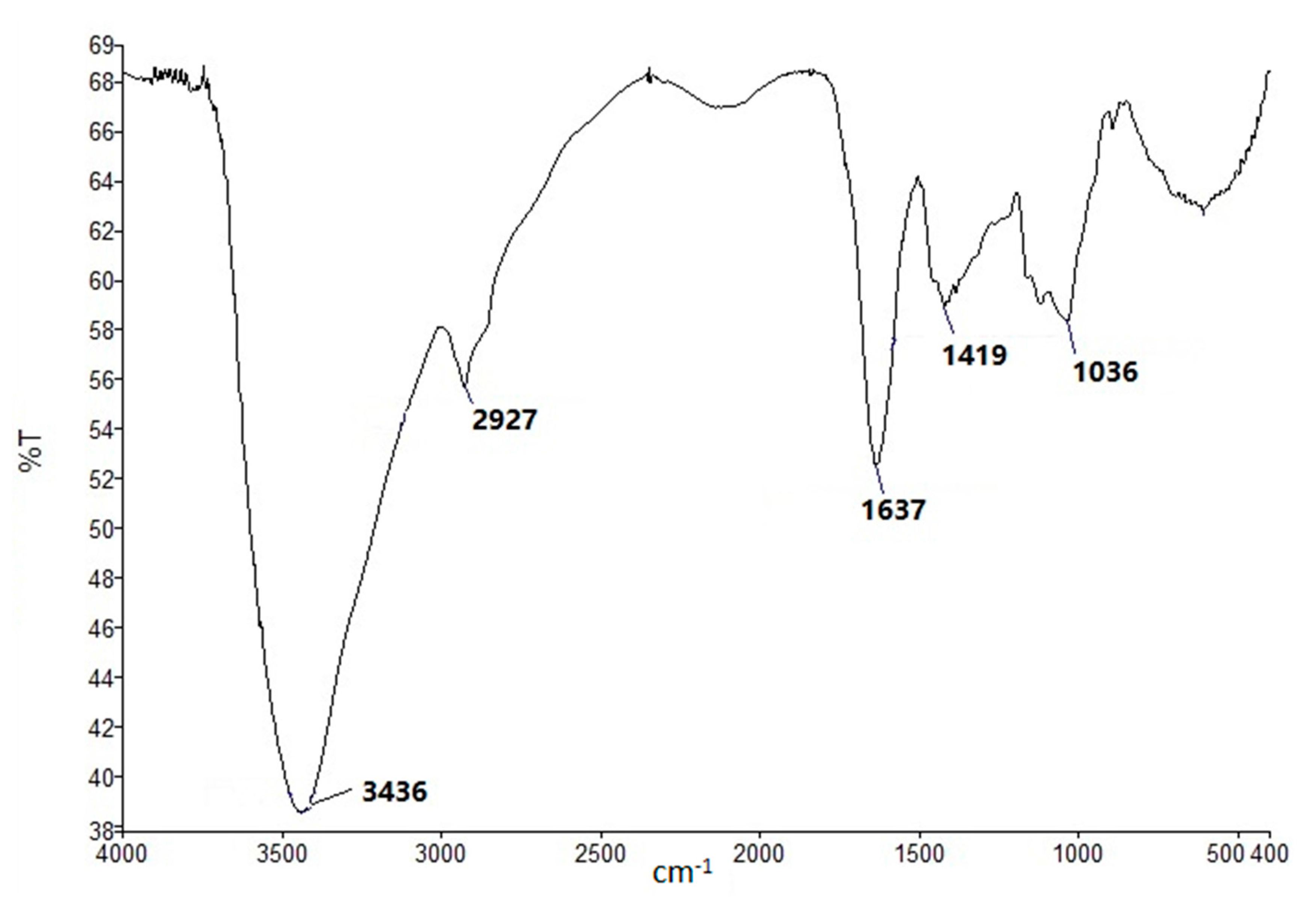
3.4. FT-IR and NMR Spectroscopy
3.5. Cell Culture
3.6. Cell Viability Assay
3.7. Quantitative Realtime PCR
3.8. ABTS Radical Scavenging Activity Assay
3.9. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Liu, Y.; Yang, J.; Cai, Z. Chemical investigation on sijunzi decoction and its two major herbs Panax ginseng and Glycyrrhiza uralensis by LC/MS/MS. J. Pharm. Biomed. Anal. 2006, 41, 1642–1647. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Cui, Z.; Zhou, Z.; Shan, T.; Li, D.; Cui, N. Si-jun-zi decoction treatment promotes the restoration of intestinal function after obstruction by regulating intestinal homeostasis. Evid. Based Complement. Altern. Med. 2014, 2014, 928579. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Xuan, Z.-R. Progress in research on applying sijunzi decoction in treating digestive malignant tumor. Chin. J. Integr. Med. 2007, 13, 156–159. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.; Zhang, S.; Cai, Z. Effects of early intestinal application of sijunzi decoction on immune function in post-operational patients of gastrointestinal tumor. Chin. J. Integr. Tradit. West. Med. 2005, 25, 1070–1073. [Google Scholar]
- Chinese Pharmacopoeia Commission. Pharmacopoeia of the People’s Republic of China; China Medical Science Press: Beijing, China, 2015; Volume 1, pp. 392–393. [Google Scholar]
- Wang, Y.-L.; Wang, X.-W.; Zeng, D.-M.; Liu, Y.-M. Microwave extraction and spectral analysis of the total polysaccharides in sijunzi decoction. Chin. J. Exp. Tradit. Med. Formulae 2012, 16, 025. [Google Scholar]
- Wang, Y.; He, S.; Cheng, X.; Lu, Y.; Zou, Y.; Zhang, Q. UPLC–Q-TOF-MS/MS fingerprinting of traditional chinese formula SiJunZiTang. J. Pharm. Biomed. Anal. 2013, 80, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Liu, J. Structural characterization of a water-soluble polysaccharide from the roots of codonopsis pilosula and its immunity activity. Int. J. Biol. Macromol. 2008, 43, 279–282. [Google Scholar]
- Zhang, X.; Zhu, C.; Li, H.; Lai, X.; Mo, J. Pharmacological action of polysaccharides from radix codonopsis on immune function and hematopoiesis in mice. Tradit. Chin. Drug Res. Clin. Pharmacol. 2003, 14, 174–176. [Google Scholar]
- Xu, C.; Liu, Y.; Yuan, G.; Guan, M. The contribution of side chains to antitumor activity of a polysaccharide from codonopsis pilosula. Int. J. Biol. Macromol. 2012, 50, 891–894. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.-F.; Chen, X.-F.; Malterud, K.E.; Rise, F.; Barsett, H.; Inngjerdingen, K.T.; Michaelsen, T.E.; Paulsen, B.S. Structural features and complement fixing activity of polysaccharides from Codonopsis pilosula Nannf. var. modesta L.T.Shen roots. Carbohydr. Polym. 2014, 113, 420–429. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Zhou, G.; Wang, M.; Peng, Y.; Li, X. The metabolism of polysaccharide from atractylodes macrocephala koidz and its effect on intestinal microflora. Evid. Based Complement. Altern. Med. 2014, 2014, 926381. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Nie, J.; Li, D.; Zhu, W.; Zhang, S.; Ma, F.; Sun, Q.; Song, J.; Zheng, Y.; Chen, P. Characterization and antioxidant activities of degraded polysaccharides from poria cocos sclerotium. Carbohydr. Polym. 2014, 105, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Wittschier, N.; Faller, G.; Hensel, A. Aqueous extracts and polysaccharides from liquorice roots (Glycyrrhiza glabra L.) inhibit adhesion of Helicobacter pylori to human gastric mucosa. J. Ethnopharmacol. 2009, 125, 218–223. [Google Scholar] [CrossRef] [PubMed]
- Han, L.; Wang, P.; Han, B. Total polysaccharide or four separate polysaccharides from sijunzi tang decoction to influence intestinal epithelial cell-6 proliferation in rat’s intestinal epithelium. Chin. Rem. Clin. 2005, 5, 496–499. [Google Scholar]
- Liu, L.; Han, L.; Wong, D.Y.; Yue, P.Y.; Ha, W.Y.; Hu, Y.H.; Wang, P.X.; Wong, R.N. Effects of Si-Jun-Zi decoction polysaccharides on cell migration and gene expression in wounded rat intestinal epithelial cells. Br. J. Nutr. 2005, 93, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Peng, Y.; Meng, H.; Li, X. Protective effect of polysaccharides fractions from sijunzi decoction in reserpine-induced spleen deficiency rats. RSC Adv. 2016, 6, 60657–60665. [Google Scholar] [CrossRef]
- McCord, J.M. The evolution of free radicals and oxidative stress. Am. J. Med. 2000, 108, 652–659. [Google Scholar] [CrossRef]
- Finkel, T.; Holbrook, N.J. Oxidants, oxidative stress and the biology of ageing. Nature 2000, 408, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Ames, B.N. Dietary carcinogens and anticarcinogens. Oxygen radicals and degenerative diseases. Science 1983, 221, 1256–1264. [Google Scholar] [CrossRef] [PubMed]
- Circu, M.L.; Aw, T.Y. Intestinal redox biology and oxidative stress. Semin. Cell Dev. Biol. 2012, 23, 729–737. [Google Scholar] [CrossRef] [PubMed]
- Piechota-Polanczyk, A.; Fichna, J. The role of oxidative stress in pathogenesis and treatment of inflammatory bowel diseases. Naunyn Schmiedebergs Arch. Pharmacol. 2014, 387, 605–620. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Gui, S.; Wang, P.; Chen, W. Purification and physi-chemical properties of polysaccharides sjzps-vb-1–2 from immunocompetence parts of sijunzi decoction. J. Chin. Med. Mater. 2003, 26, 751–753. [Google Scholar]
- Cao, G.; Zhang, X.; Cong, X. The research progress of polysaccharides from atractylodes macrocephala koidz. J. Beijing Union Univ. (Nat. Sci.) 2009, 23, 14–18. [Google Scholar]
- Chen, X.; Tang, Q.; Chen, Y.; Wang, W.; Li, S. Simultaneous extraction of polysaccharides from poria cocos by ultrasonic technique and its inhibitory activities against oxidative injury in rats with cervical cancer. Carbohydr. Polym. 2010, 79, 409–413. [Google Scholar] [CrossRef]
- Zhang, C.-H.; Yu, Y.; Liang, Y.-Z.; Chen, X.-Q. Purification, partial characterization and antioxidant activity of polysaccharides from Glycyrrhiza uralensis. Int. J. Biol. Macromol. 2015, 79, 681–686. [Google Scholar] [CrossRef] [PubMed]
- Jia, X.; Ma, L.; Li, P.; Chen, M.; He, C. Prospects of poria cocos polysaccharides: Isolation process, structural features and bioactivities. Trends Food Sci. Technol. 2016, 54, 52–62. [Google Scholar] [CrossRef]
- Waldron, K.; Faulds, C. Cell wall polysaccharides: Composition and structure. Compr. Glycosci. 2007, 1, 181–201. [Google Scholar]
- Maxwell, E.G.; Belshaw, N.J.; Waldron, K.W.; Morris, V.J. Pectin—An emerging new bioactive food polysaccharide. Trends Food Sci. Technol. 2012, 24, 64–73. [Google Scholar] [CrossRef]
- Zhang, Q.; Xu, Y.; Zou, S.; Zhang, X.; Cao, K.; Fan, Q. Novel functional polysaccharides from radix polygoni multiflori water extracted residue: Preliminary characterization and immunomodulatory activity. Carbohydr. Polym. 2016, 137, 625–631. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Nie, S.-P.; Yin, J.-Y.; Wang, Y.-X.; Xie, M.-Y. Structural characterization of a heterogalactan purified from fruiting bodies of ganoderma atrum. Food Hydrocoll. 2014, 36, 339–347. [Google Scholar] [CrossRef]
- Ho, G.T.T.; Zou, Y.-F.; Aslaksen, T.H.; Wangensteen, H.; Barsett, H. Structural characterization of bioactive pectic polysaccharides from elderflowers (sambuci flos). Carbohydr. Polym. 2016, 135, 128–137. [Google Scholar] [CrossRef] [PubMed]
- Chai, Y.; Zhao, M. Purification, characterization and anti-proliferation activities of polysaccharides extracted from Viscum coloratum (Kom.) Nakai. Carbohydr. Polym. 2016, 149, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Fan, L.; Ding, S. Isolation, purification and structure of a new water-soluble polysaccharide from Zizyphus jujuba cv. Jinsixiaozao. Carbohydr. Polym. 2011, 83, 477–482. [Google Scholar] [CrossRef]
- Li, J.; Ai, L.; Yang, Q.; Liu, Y.; Shan, L. Isolation and structural characterization of a polysaccharide from fruits of Zizyphus jujuba cv. Junzao. Int. J. Biol. Macromol. 2013, 55, 83–87. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Liu, Y.; Zhu, R.; Yu, J.; Lu, W.; Pan, C.; Yao, W.; Gao, X. Structure characterization, chemical and enzymatic degradation, and chain conformation of an acidic polysaccharide from Lycium barbarum L. Carbohydr. Polym. 2016, 147, 114–124. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Zhang, R.; Liu, Y.; Xiao, J.; Su, D.; Yi, Y.; Wang, G.; Wei, Z.; Zhang, M. Characterization and mesenteric lymph node cells-mediated immunomodulatory activity of litchi pulp polysaccharide fractions. Carbohydr. Polym. 2016, 152, 496–503. [Google Scholar] [CrossRef] [PubMed]
- Aw, T.Y. Molecular and cellular responses to oxidative stress and changes in oxidation-reduction imbalance in the intestine. Am. J. Clin. Nutr. 1999, 70, 557–565. [Google Scholar] [PubMed]
- Bhattacharyya, A.; Chattopadhyay, R.; Mitra, S.; Crowe, S.E. Oxidative stress: An essential factor in the pathogenesis of gastrointestinal mucosal diseases. Physiol. Rev. 2014, 94, 329–354. [Google Scholar] [CrossRef] [PubMed]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.T.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Austarheim, I.; Christensen, B.E.; Hegna, I.K.; Petersen, B.O.; Duus, J.O.; Bye, R.; Michaelsen, T.E.; Diallo, D.; Inngjerdingen, M.; Paulsen, B.S. Chemical and biological characterization of pectin-like polysaccharides from the bark of the malian medicinal tree cola cordifolia. Carbohydr. Polym. 2012, 89, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.B.; Carpita, N.C. Changes in esterification of the uronic acid groups of cell wall polysaccharides during elongation of maize coleoptiles. Plant Physiol. 1992, 98, 646–653. [Google Scholar] [CrossRef] [PubMed]
- Ciucanu, I.; Kerek, F. A simple and rapid method for the permethylation of carbohydrates. Carbohydr. Res. 1984, 131, 209–217. [Google Scholar] [CrossRef]
- Sweet, D.P.; Shapiro, R.H.; Albersheim, P. Quantitative analysis by various glc response-factor theories for partially methylated and partially ethylated alditol acetates. Carbohydr. Res. 1975, 40, 217–225. [Google Scholar] [CrossRef]
- Zhu, Z.-Y.; Liu, R.-Q.; Si, C.-L.; Zhou, F.; Wang, Y.-X.; Ding, L.-N.; Jing, C.; Liu, A.-J.; Zhang, Y.-M. Structural analysis and anti-tumor activity comparison of polysaccharides from astragalus. Carbohydr. Polym. 2011, 85, 895–902. [Google Scholar] [CrossRef]
- Yan, J.-K.; Li, L.; Wang, Z.-M.; Wu, J.-Y. Structural elucidation of an exopolysaccharide from mycelial fermentation of a Tolypocladium sp. fungus isolated from wild Cordyceps sinensis. Carbohydr. Polym. 2010, 79, 125–130. [Google Scholar] [CrossRef]
- Huang, C.; Chen, M.; Pang, D.; Bi, D.; Zou, Y.; Xia, X.; Yang, W.; Luo, L.; Deng, R.; Tan, H.; et al. Developmental and activity-dependent expression of lancl1 confers antioxidant activity required for neuronal survival. Dev. Cell 2014, 30, 479–487. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds SJZDP-I-I, SJZDP-I-II and SJZDP-II-I are available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SJZDP-I-I | SJZDP-I-II | SJZDP-II-I | |
|---|---|---|---|
| Ara a | 51.2 | 18.9 | 32.8 |
| Rha a | 2.3 | 1.7 | 5.0 |
| Fuc a | 2.1 | n.d. | 0.4 |
| Xyl a | n.d. | 2.4 | 0.4 |
| Man a | 3.7 | 1.4 | 0.4 |
| Gal a | 23.6 | 4.7 | 40.5 |
| Glc a | 11.2 | 66.9 | 1.5 |
| GlcA a | 0.8 | n.d. | 1.5 |
| GalA a | 5.2 | 4.0 | 17.4 |
| Mw (kDa) | 351.0 | 16.9 | 171.9 |
| Linkage Type | mol % | |
|---|---|---|
| Ara | Tf | 22.7 |
| 1→5f | 3.7 | |
| 1→3,5f | 6.4 | |
| Rha | Tp | 0.4 |
| 1→2p | 1.8 | |
| 1→2, 4p | 2.8 | |
| Gal | Tp | 2.4 |
| 1→4p | 10.7 | |
| 1→3p | 2.2 | |
| 1→6p | 5.1 | |
| 1→3, 6p | 20.1 | |
| Glc | Tp | 1.5 |
| GlcA | Tp | 1.5 |
| GalA | Tp | 0.9 |
| 1→4p | 16.5 |
| SOD1 | Fr 5′-CAAGCGGTGAACCAGTTGTG-3′ |
| Rv 5′-TGAGGTCCTGCACTGGTAC-3′ | |
| SOD2 | Fr 5′-GCCTGCACTGAAGTTCAATG-3′ |
| Rv 5′-ATCTGTAAGCGACCTTGCTC-3′ | |
| Catalase | Fr 5′-ACCCTCTTATACCAGTTGGC-3′ |
| Rv 5′-GCATGCACATGGGGCCATCA-3′ | |
| GSTP1 | Fr 5′-ATGCCACCATACACCATTGTC-3′ |
| Rv 5′-GGGAGCTGCCCATACAGAC-3′ | |
| PGC-1α | Fr 5′-CTCCCTGTGGATGAAGACGG-3′ |
| Rv 5′-GCAAATCACAATCACAGGAT-3′ |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, C.; Zhu, Z.; Cao, X.; Chen, X.; Fu, Y.; Chen, Z.; Li, L.; Song, X.; Jia, R.; Yin, Z.; et al. A Pectic Polysaccharide from Sijunzi Decoction Promotes the Antioxidant Defenses of SW480 Cells. Molecules 2017, 22, 1341. https://doi.org/10.3390/molecules22081341
Huang C, Zhu Z, Cao X, Chen X, Fu Y, Chen Z, Li L, Song X, Jia R, Yin Z, et al. A Pectic Polysaccharide from Sijunzi Decoction Promotes the Antioxidant Defenses of SW480 Cells. Molecules. 2017; 22(8):1341. https://doi.org/10.3390/molecules22081341
Chicago/Turabian StyleHuang, Chao, Zhongkai Zhu, Xiyue Cao, Xingfu Chen, Yuping Fu, Zhengli Chen, Lixia Li, Xu Song, Renyong Jia, Zhongqiong Yin, and et al. 2017. "A Pectic Polysaccharide from Sijunzi Decoction Promotes the Antioxidant Defenses of SW480 Cells" Molecules 22, no. 8: 1341. https://doi.org/10.3390/molecules22081341
APA StyleHuang, C., Zhu, Z., Cao, X., Chen, X., Fu, Y., Chen, Z., Li, L., Song, X., Jia, R., Yin, Z., Ye, G., Feng, B., & Zou, Y. (2017). A Pectic Polysaccharide from Sijunzi Decoction Promotes the Antioxidant Defenses of SW480 Cells. Molecules, 22(8), 1341. https://doi.org/10.3390/molecules22081341