Dietary Copper Reduces the Hepatotoxicity of (−)-Epigallocatechin-3-Gallate in Mice

 , and
, and
Abstract
:
1. Introduction
2. Results
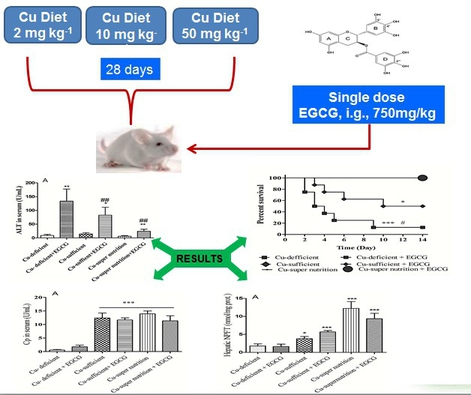
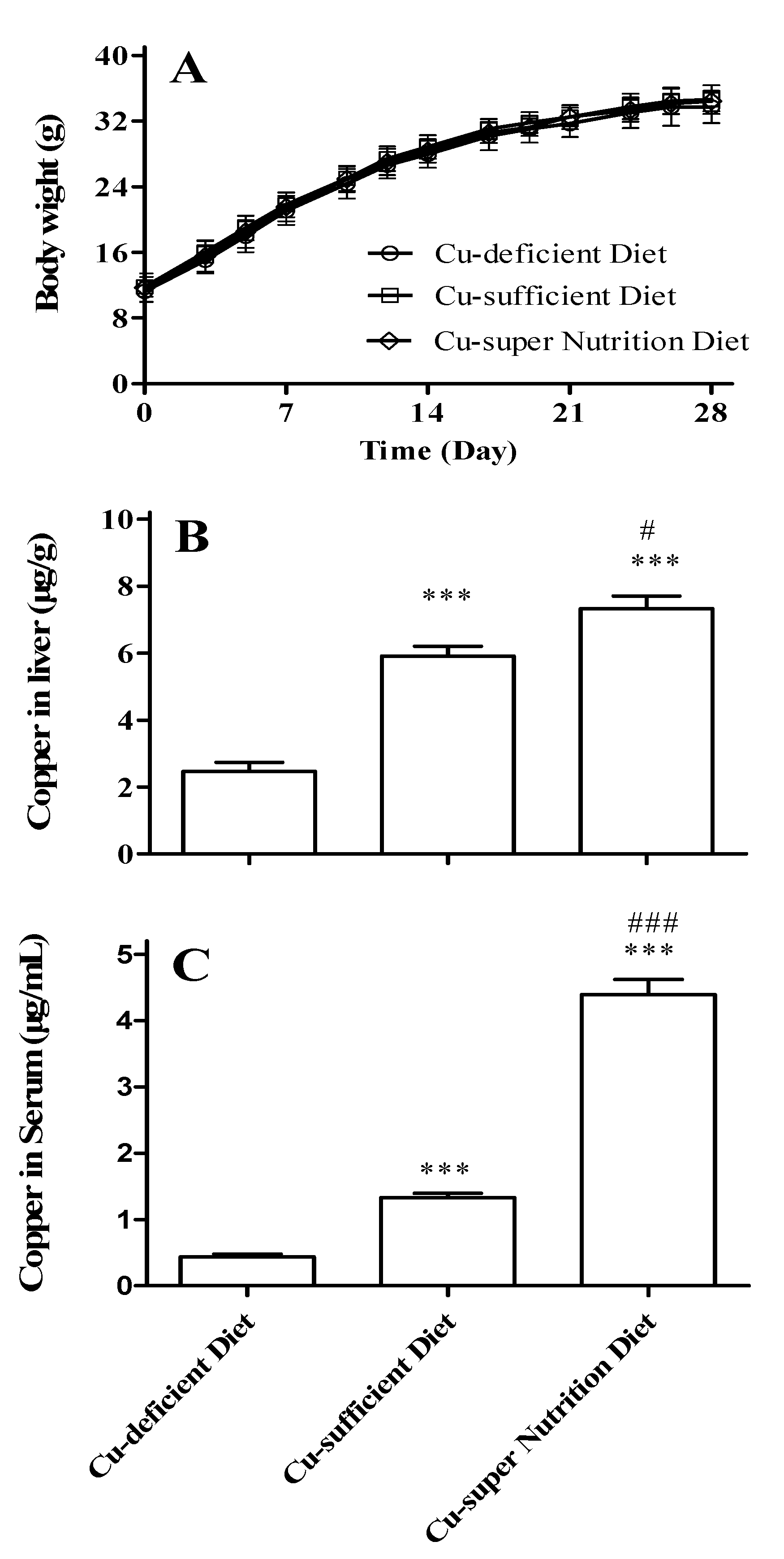
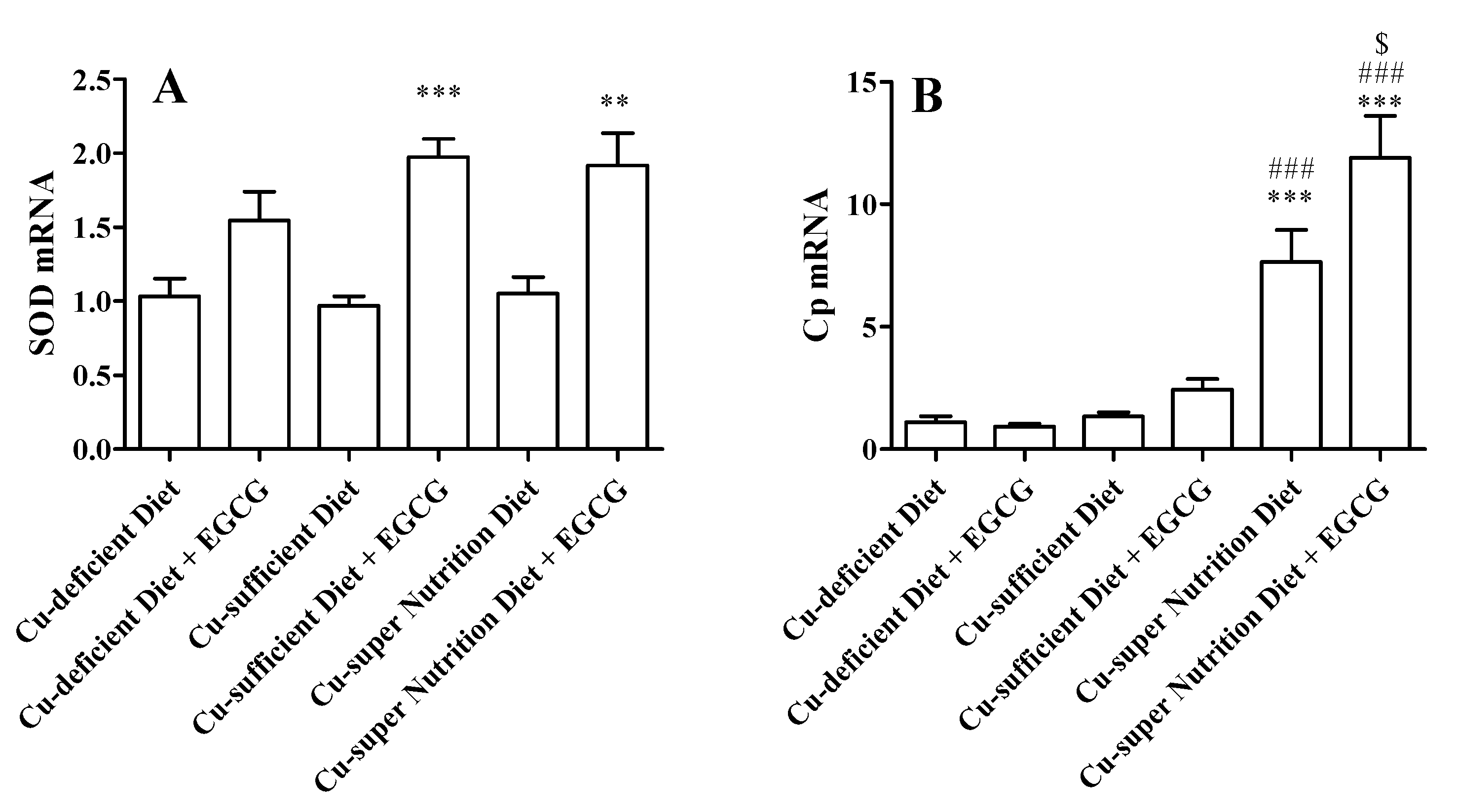
2.1. Generation of Mice Models with Different Copper Status
2.2. Copper Decreased the Lethal Effects of EGCG
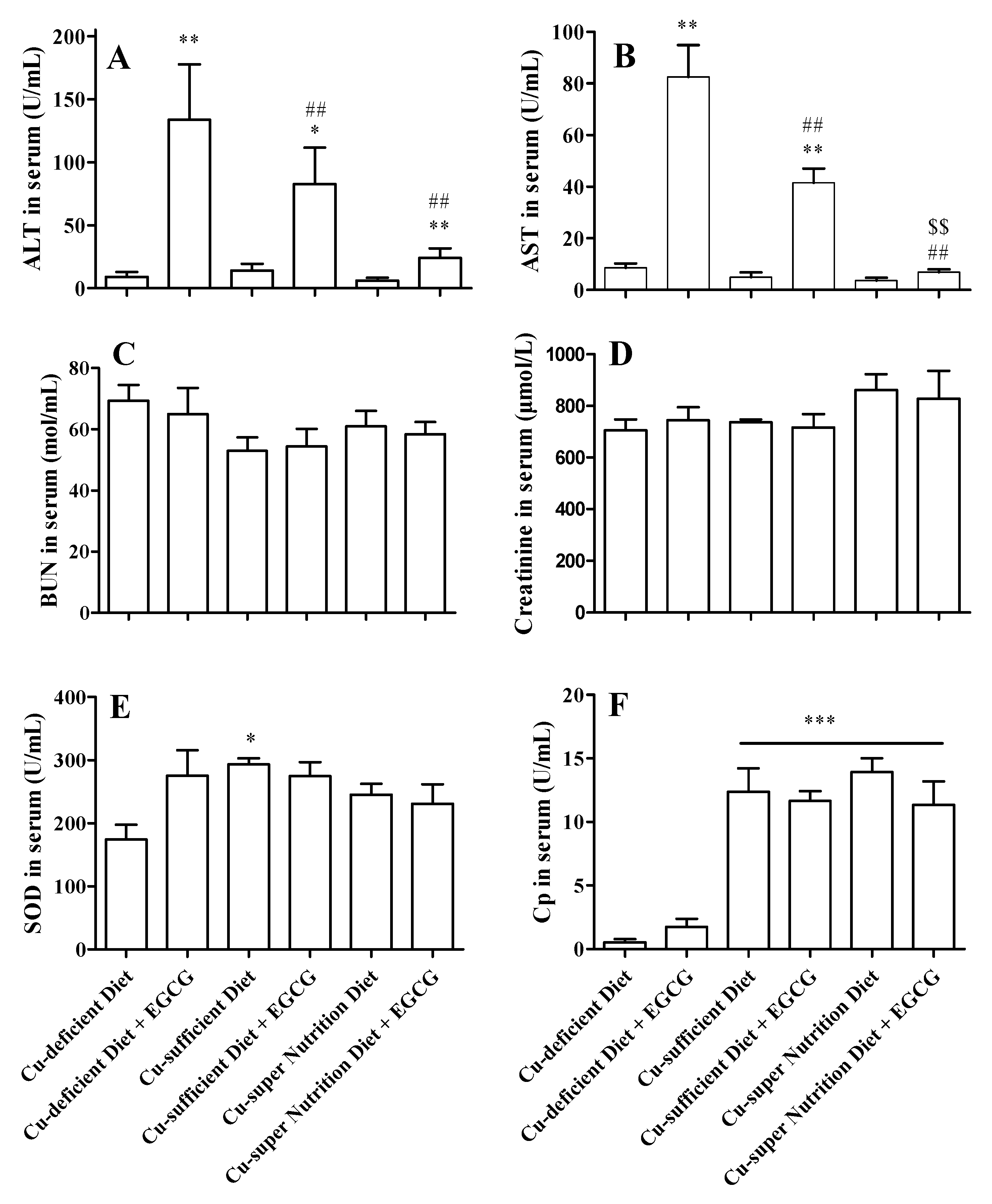
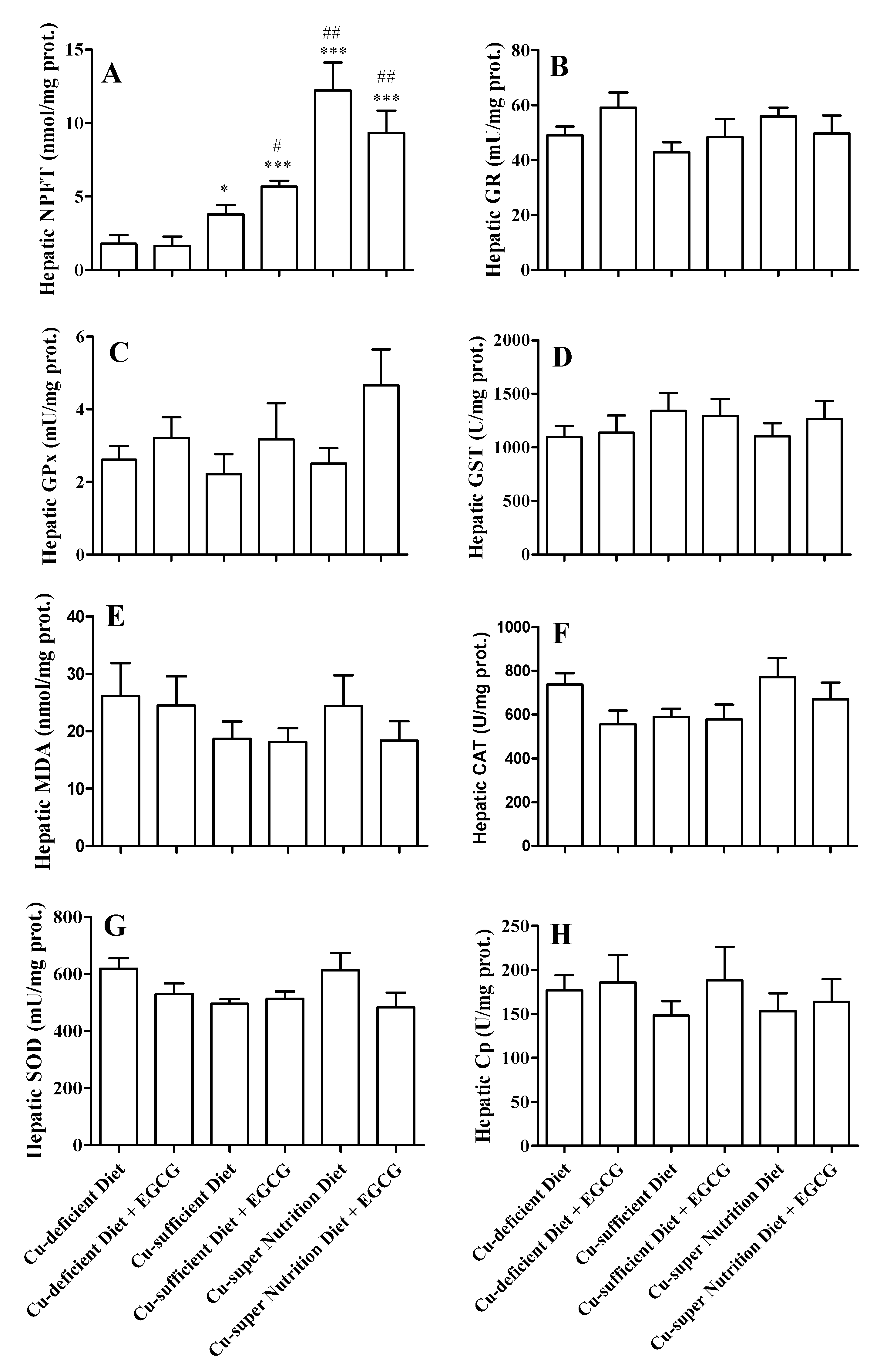
2.3. Copper Reduced the Acute Hepatotoxicity of EGCG
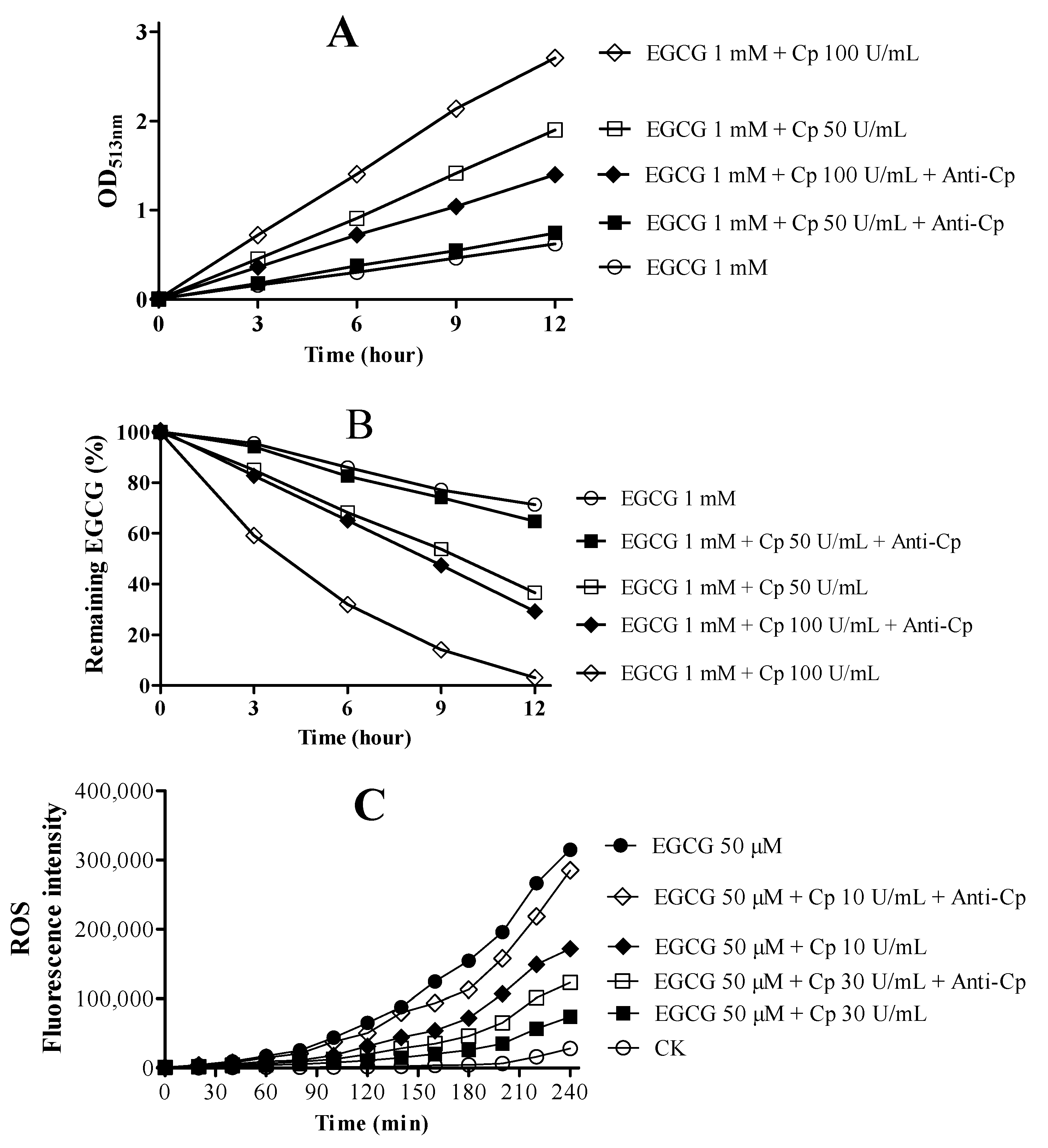
2.4. Cp Promotes EGCG Oxidation with Low Generation of ROS In Vitro
3. Discussion
4. Materials and Methods
4.1. Ethical Guidelines
4.2. Chemicals and Drugs
4.3. Animals Experiments
4.4. Tissue Preparation and Biomarkers Assessment
4.5. RNA Isolation, Reverse Transcriptase Polymerase Chain Reaction and Real-Time PCR
4.6. EGCG Oxidation Assessment
4.7. ROS Detection
4.8. Statistical Analyses
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Singh, B.N.; Shankar, S.; Srivastava, R.K. Green tea catechin, epigallocatechin-3-gallate (EGCG): Mechanisms, perspectives and clinical applications. Biochem. Pharmacol. 2011, 82, 1807–1821. [Google Scholar] [CrossRef] [PubMed]
- Afzal, M.; Safer, A.M.; Menon, M. Green tea polyphenols and their potential role in health and disease. Inflammopharmacology 2015, 23, 151–161. [Google Scholar] [CrossRef] [PubMed]
- Sabu, M.C.; Priya, T.T.; Ramadasan, K.; Ikuo, N. Beneficial effects of green tea: A literature review. Chin. Med. 2010, 5, 13. [Google Scholar]
- Chowdhury, A.; Sarkar, J.; Chakraborti, T.; Pramanik, P.K.; Chakraborti, S. Protective role of epigallocatechin-3-gallate in health and disease: A perspective. Biomed. Pharmacother. Biomed. Pharmacother. 2016, 78, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Cabrera, C.; Artacho, R.; Giménez, R. Beneficial Effects of Green Tea—A Review. J. Am. Coll. Nutr. 2006, 25, 79–99. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.S.; Wang, H. Mechanistic issues concerning cancer prevention by tea catechins. Mol. Nutr. Food Res. 2011, 55, 819–831. [Google Scholar] [CrossRef] [PubMed]
- Shen, G.; Xu, C.; Hu, R.; Jain, M.R.; Nair, S.; Lin, W.; Yang, C.S.; Chan, J.Y.; Kong, A.N. Comparison of (−)-epigallocatechin-3-gallate elicited liver and small intestine gene expression profiles between C57BL/6J mice and C57BL/6J/Nrf2 (−/−) mice. Pharm. Res. 2005, 22, 1805–1820. [Google Scholar] [CrossRef] [PubMed]
- Lambert, J.D.; Kennett, M.J.; Sang, S.M.; Reuhl, K.R.; Ju, J.Y.; Yang, C.S. Hepatotoxicity of high oral dose (−)-epigallocatechin-3-gallate in mice. Food Chem. Toxicol. 2010, 48, 409–416. [Google Scholar] [CrossRef] [PubMed]
- Isbrucker, R.A.; Edwards, J.A.; Wolz, E.; Davidovich, A.; Bausch, J. Safety studies on epigallocatechin gallate (EGCG) preparations. Part 2: Dermal, acute and short-term toxicity studies. Food Chem. Toxicol. 2006, 44, 636–650. [Google Scholar] [CrossRef] [PubMed]
- Galati, G.; Lin, A.; Sultan, A.M.; O’Brien, P.J. Cellular and in vivo hepatotoxicity caused by green tea phenolic acids and catechins. Free Radic. Boil. Med. 2006, 40, 570–580. [Google Scholar] [CrossRef] [PubMed]
- Mazzanti, G.; Menniti-Ippolito, F.; Moro, P.A.; Cassetti, F.; Raschetti, R.; Santuccio, C.; Mastrangelo, S. Hepatotoxicity from green tea: A review of the literature and two unpublished cases. Eur. J. Clin. Pharmacol. 2009, 65, 331–341. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Wei, Y.; Wang, T.; Wan, X.; Yang, C.S.; Reiter, R.J.; Zhang, J. Melatonin attenuates (−)-epigallocatehin-3-gallate-triggered hepatotoxicity without compromising its downregulation of hepatic gluconeogenic and lipogenic genes in mice. J. Pineal Res. 2015, 59, 497–507. [Google Scholar] [CrossRef] [PubMed]
- Mazzanti, G.; Di, S.A.; Vitalone, A. Hepatotoxicity of green tea: An update. Arch. Toxicol. 2015, 89, 1175–1191. [Google Scholar] [CrossRef] [PubMed]
- Prohaska, J.R. Comparison between dietary and cenetic copper deficiency in mice: Copper-dependent anemia. Nutr. Res. 1981, 1, 159–167. [Google Scholar] [CrossRef]
- Harris, Z.L.; Gitlin, J.D. Genetic and molecular basis for copper toxicity. Am. J. Clin. Nutr. 1996, 63, 836S. [Google Scholar] [PubMed]
- Lisa, M.G.; Hannah, S.C.-J.; Ching, K.C. Copper: Toxicological relevance and mechanisms. Arch. Toxicol. 2014, 88, 1929–1938. [Google Scholar]
- Ishino, A.; Mita, S.; Watanabe, S.; Sakagami, H. Effect of anticancer drugs, metals and antioxidants on cytotoxic activity of epigallocatechin gallate. Anticancer Res. 1999, 19, 4343–4348. [Google Scholar] [PubMed]
- Lambert, J.D.; Elias, R.J. The antioxidant and pro-oxidant activities of green tea polyphenols: A role in cancer prevention. Arch. Biochem. Biophys. 2010, 501, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Broderius, M.; Mostad, E.; Wendroth, K.; Prohaska, J.R. Levels of plasma ceruloplasmin protein are markedly lower following dietary copper deficiency in rodents. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2010, 151, 473–479. [Google Scholar] [CrossRef] [PubMed]
- Shim, H.; Harris, Z.L. Genetic defects in copper metabolism. J. Nutr. 2003, 133, 1527S. [Google Scholar] [PubMed]
- Farhan, M.; Oves, M.; Chibber, S.; Hadi, S.M.; Ahmad, A. Mobilization of Nuclear Copper by Green Tea Polyphenol Epicatechin-3-Gallate and Subsequent Prooxidant Breakage of Cellular DNA: Implications for Cancer Chemotherapy. Int. J. Mol. Sci. 2016, 18, 34. [Google Scholar] [CrossRef] [PubMed]
- Azam, S.; Hadi, N.; Khan, N.U.; Hadi, S.M. Prooxidant property of green tea polyphenols epicatechin and epigallocatechin-3-gallate: Implications for anticancer properties. Toxicol. In Vitro 2004, 18, 555–561. [Google Scholar] [CrossRef] [PubMed]
- Malik, A.; Azam, S.; Hadi, N.; Hadi, S.M. DNA degradation by water extract of green tea in the presence of copper ions: Implications for anticancer properties. Phytother. Res. 2010, 17, 358–363. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; Wang, B.; Zhu, L. DNA binding and oxidative DNA damage induced by a quercetin copper(II) complex: Potential mechanism of its antitumor properties. J. Biol. Inorg. Chem. 2009, 14, 727–739. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, N.; Tanemura, H.; Kawanishi, S. Mechanism of oxidative DNA damage induced by quercetin in the presence of Cu(II). Mutat. Res. 1999, 425, 107–115. [Google Scholar] [CrossRef]
- Ahsan, H.; Hadi, S.M. Strand scission in DNA induced by curcumin in the presence of Cu(II). Cancer Lett. 1998, 124, 23–30. [Google Scholar] [CrossRef]
- Wang, X.; Wang, H.; Li, J.; Yang, Z.; Zhang, J.; Qin, Z.; Wang, L.; Kong, X. Evaluation of Bioaccumulation and Toxic Effects of Copper on Hepatocellular Structure in Mice. Biol. Trace Elem. Res. 2014, 159, 312–319. [Google Scholar] [CrossRef] [PubMed]
- Sokol, R.J.; Devereaux, M.W.; O’Brien, K.; Khandwala, R.A.; Loehr, J.P. Abnormal hepatic mitochondrial respiration and cytochrome C oxidase activity in rats with long-term copper overload. Gastroenterology 1993, 105, 178–187. [Google Scholar] [CrossRef]
- Lin, W.H.; Chen, M.D.; Wang, C.C.; Lin, P.Y. Dietary copper supplementation increases the catecholamine levels in genetically obese (ob/ob) mice. Biol. Trace Elem. Res. 1995, 50, 243–247. [Google Scholar] [CrossRef] [PubMed]
- Hurley, L.S.; Bell, L.T. Amelioration by copper supplementation of mutant gene effects in the crinkled mouse. Proc. Soc. Exp. Biol. Med. 1975, 149, 830–834. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhu, S.; Hodgkinson, V.; Prohaska, J.R.; Weisman, G.A.; Gitlin, J.D.; Petris, M.J. Maternofetal and neonatal copper requirements revealed by enterocyte-specific deletion of the Menkes disease protein. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 303, 1236–1244. [Google Scholar] [CrossRef] [PubMed]
- Hebert, C. NTP technical report on the toxicity studies of Cupric Sulfate (CAS No. 7758-99-8) Administered in Drinking Water and Feed to F344/N Rats and B6C3F1 Mice. Toxic. Rep. 1993, 29, 1–D3. [Google Scholar]
- Massie, H.R.; Aiello, V.R. Excessive intake of copper: Influence on longevity and cadmium accumulation in mice. Mechan. Ageing Dev. 1984, 26, 195–203. [Google Scholar] [CrossRef]
- Pribyl, T. Serum polyphenol oxidase activity (ceruloplasmin) in conventional laboratory animals and man. Folia Biol. 1978, 24, 136–141. [Google Scholar]
- Hou, Z.; Sang, S.; You, H.; Lee, M.J.; Hong, J.; Chin, K.V.; Yang, C.S. Mechanism of action of (−)-epigallocatechin-3-gallate: Auto-oxidation-dependent inactivation of epidermal growth factor receptor and direct effects on growth inhibition in human esophageal cancer KYSE 150 cells. Cancer Res. 2005, 65, 8049–8056. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Dong, R.; Sun, K.; Wang, X.; Wang, J.; Yang, C.S.; Zhang, J. Synergistic toxicity of epigallocatechin-3-gallate and diethyldithiocarbamate, a lethal encounter involving redox-active copper. Free Radic. Boil. Med. 2017, 113, 143–156. [Google Scholar] [CrossRef] [PubMed]
- Khan, H.Y.; Zubair, H.; Ullah, M.F.; Ahmad, A.; Hadi, S.M. Oral administration of copper to rats leads to increased lymphocyte cellular DNA degradation by dietary polyphenols: Implications for a cancer preventive mechanism. Biometals 2011, 24, 1169–1178. [Google Scholar] [CrossRef] [PubMed]
- Khan, H.Y.; Zubair, H.; Faisal, M.; Ullah, M.F.; Farhan, M.; Sarkar, F.H.; Ahmad, A.; Hadi, S.M. Plant polyphenol induced cell death in human cancer cells involves mobilization of intracellular copper ions and reactive oxygen species generation: A mechanism for cancer chemopreventive action. Mol. Nutr. Food Res. 2014, 58, 437–446. [Google Scholar] [CrossRef] [PubMed]
- Heikkila, R.E.; Cabbat, F.S.; Cohen, G. In vivo inhibition of superoxide dismutase in mice by diethyldithiocarbamate. J. Biol. Chem. 1976, 251, 2182–2185. [Google Scholar] [PubMed]
- Valentine, H.L.; Viquez Omvalentine, W.M. Peripheral nerve and brain differ in their capacity to resolve N,N-diethyldithiocarbamate-mediated elevations in copper and oxidative injury. Toxicology 2010, 274, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Viquez, O.M.; Valentine, H.L.; Amarnath, K.; Milatovic, D.; Valentine, W.M. Copper accumulation and lipid oxidation precede inflammation and myelin lesions in N,N-diethyldithiocarbamate peripheral myelinopathy. Toxicol. Appl. Pharmacol. 2008, 229, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Howell, J.M. Effect of sodium Diethyldithiocarbamate on blood copper-levels and pregnancy in the rabbit. Nature 1964, 201, 83–84. [Google Scholar] [CrossRef] [PubMed]
- Sunderman, F.W. Nickel and copper mobilization by sodium Diethyldithiocarbamate. J. New Drugs 1964, 4, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Letelier, M.E.; Faúndez, M.; Jara-Sandoval, J.; Molina-Berríos, A.; Cortés-Troncoso, J.; Aracena-Parks, P.; Marín-Catalán, R. Mechanisms underlying the inhibition of the cytochrome P450 system by copper ions. J. Appl. Toxicol. 2010, 29, 695–702. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Ahn, T.; Yim, S.K.; Yun, C.H. Differential effect of copper(II) on the cytochrome P450 enzymes and NADPH-cytochrome P450 reductase: Inhibition of cytochrome P450-catalyzed reactions by copper(II) ion. Biochemistry 2002, 41, 9438–9447. [Google Scholar] [CrossRef] [PubMed]
- Korobkova, E.A. Effect of Natural Polyphenols on CYP Metabolism: Implications for Diseases. Chem. Res. Toxicol. 2015, 28, 1359–1390. [Google Scholar] [CrossRef] [PubMed]
- Teschke, R.; Zhang, L.; Melzer, L.; Schulze, J.; Eickhoff, A. Green tea extract and the risk of drug-induced liver injury. Expert Opin. Drug Metab. Toxicol. 2014, 10, 1663–1676. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, J.; Xu, T. Cyclophosphamide-evoked heart failure involves pronounced co-suppression of cytoplasmic thioredoxin reductase activity and non-protein free thiol level. Eur. J. Heart Fail. 2009, 11, 154–162. [Google Scholar] [CrossRef] [PubMed]
- Carlberg, I.; Mannervik, B. Glutathione reductase. Methods Enzymol. 1985, 113, 484–490. [Google Scholar] [PubMed]
- Smith, A.D.; Levander, O.A. High-throughput 96-well microplate assays for determining specific activities of glutathione peroxidase and thioredoxin reductase. Methods Enzymol. 2002, 347, 113–120. [Google Scholar] [PubMed]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione S-transferases. The first enzymatic step in mercapturic acid formation. J. Boil. Chem. 1974, 249, 7130–7139. [Google Scholar]
- Sun, Y.; Oberley, L.W.; Li, Y. A simple method for clinical assay of superoxide dismutase. Clin. Chem. 1988, 34, 497–500. [Google Scholar] [PubMed]
- Claiborne, A. Catalase activity. In CRC Handbook of Methods for Oxygen Radical Research; Greenwald, R.A., Ed.; CRC Press: Boca Raton, FL, USA, 1985; pp. 283–284. [Google Scholar]
Sample Availability: Samples of the compounds are not available from the authors. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Copper-Deficient Diet | Copper-Sufficient Diet | Copper-Super Nutrition Diet | |
|---|---|---|---|
| Casein (g) | 200.0 | 200.0 | 200.0 |
| l-cystine (g) | 30.0 | 30.0 | 30.0 |
| Corn Starch (g) | 397.5 | 397.5 | 397.5 |
| Maltodextrin (g) | 132.0 | 132.0 | 132.0 |
| Sucrose (g) | 100.0 | 100.0 | 100.0 |
| Cellulose (g) | 50.0 | 50.0 | 50.0 |
| Soybean Oil (g) | 70.0 | 70.0 | 70.0 |
| Choline Bitartrate (g) | 2.5 | 2.5 | 2.5 |
| Vitamin Complex (g) | 10.0 | 10.0 | 10.0 |
| Mine Complex (g) | 35.0 | 35.0 | 35.0 |
| Ca (g) | 5.0 | 5.0 | 5.0 |
| P (g) | 3.0 | 3.0 | 3.0 |
| K (g) | 3.6 | 3.6 | 3.6 |
| S (g) | 0.3 | 0.3 | 0.3 |
| Mg (g) | 0.5 | 0.5 | 0.5 |
| Na (g) | 1.0 | 1.0 | 1.0 |
| Cl (g) | 1.6 | 1.6 | 1.6 |
| Cu (mg) * | 0.0 | 10.0 | 50.0 |
| Fe (mg) | 45.0 | 45.0 | 45.0 |
| Mn (mg) | 10.2 | 10.2 | 10.2 |
| Zn (mg) | 30.0 | 30.0 | 30.0 |
| F (mg) | 1.0 | 1.0 | 1.0 |
| Se (mg) | 0.2 | 0.2 | 0.2 |
| I (mg) | 0.2 | 0.2 | 0.2 |
| Genes | Primers | Sequences |
|---|---|---|
| SOD1 | Sense | TGGAGACCTGGGCAATGTGACT |
| Antisense | TCCACCTTTGCCCAAGTCATCT | |
| Cp1 | Sense | TTCAGTGCCAGAAGCATAGTCCCA |
| Antisense | GGCCACAGGGAACTGTGTTTGTTT | |
| β-Actin | Sense | GCTGAGAGGGAAATCGTGCGT |
| Antisense | ACCGCTCGTTGCCAATAGTGA |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaleri, N.A.; Sun, K.; Wang, L.; Li, J.; Zhang, W.; Chen, X.; Li, X. Dietary Copper Reduces the Hepatotoxicity of (−)-Epigallocatechin-3-Gallate in Mice. Molecules 2018, 23, 38. https://doi.org/10.3390/molecules23010038
Kaleri NA, Sun K, Wang L, Li J, Zhang W, Chen X, Li X. Dietary Copper Reduces the Hepatotoxicity of (−)-Epigallocatechin-3-Gallate in Mice. Molecules. 2018; 23(1):38. https://doi.org/10.3390/molecules23010038
Chicago/Turabian StyleKaleri, Najeeb Ahmed, Kang Sun, Le Wang, Jin Li, Wenzheng Zhang, Xuan Chen, and Xinghui Li. 2018. "Dietary Copper Reduces the Hepatotoxicity of (−)-Epigallocatechin-3-Gallate in Mice" Molecules 23, no. 1: 38. https://doi.org/10.3390/molecules23010038
APA StyleKaleri, N. A., Sun, K., Wang, L., Li, J., Zhang, W., Chen, X., & Li, X. (2018). Dietary Copper Reduces the Hepatotoxicity of (−)-Epigallocatechin-3-Gallate in Mice. Molecules, 23(1), 38. https://doi.org/10.3390/molecules23010038