
Optimization of Culture Condition for Ganoderic Acid Production in Ganoderma lucidum Liquid Static Culture and Design of a Suitable Bioreactor


Abstract
:1. Introduction
2. Results and Discussion
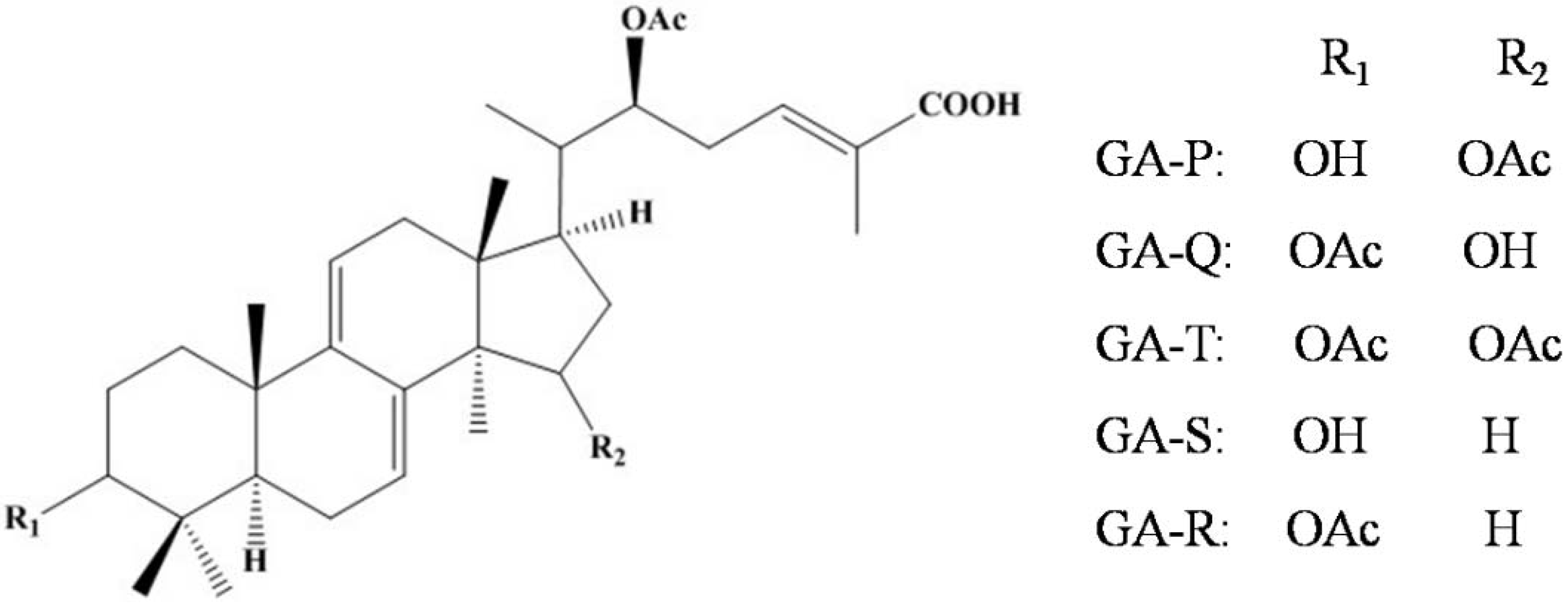
2.1. Isolation and Structural Elucidation of GAs
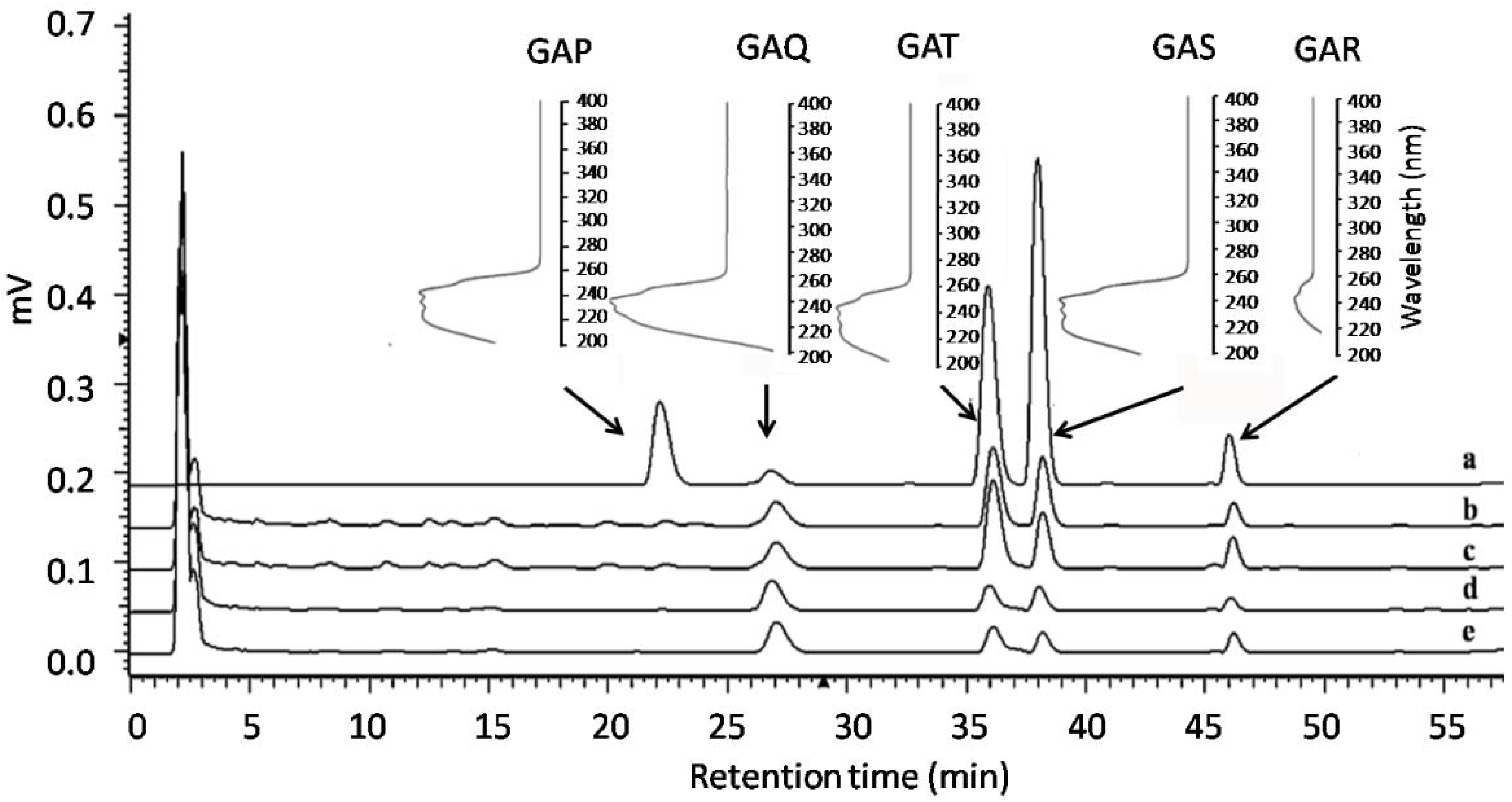
2.2. Content Determination of Five GAs in Mycelial Matsin Different Experimental Groups Using RP-HPLC
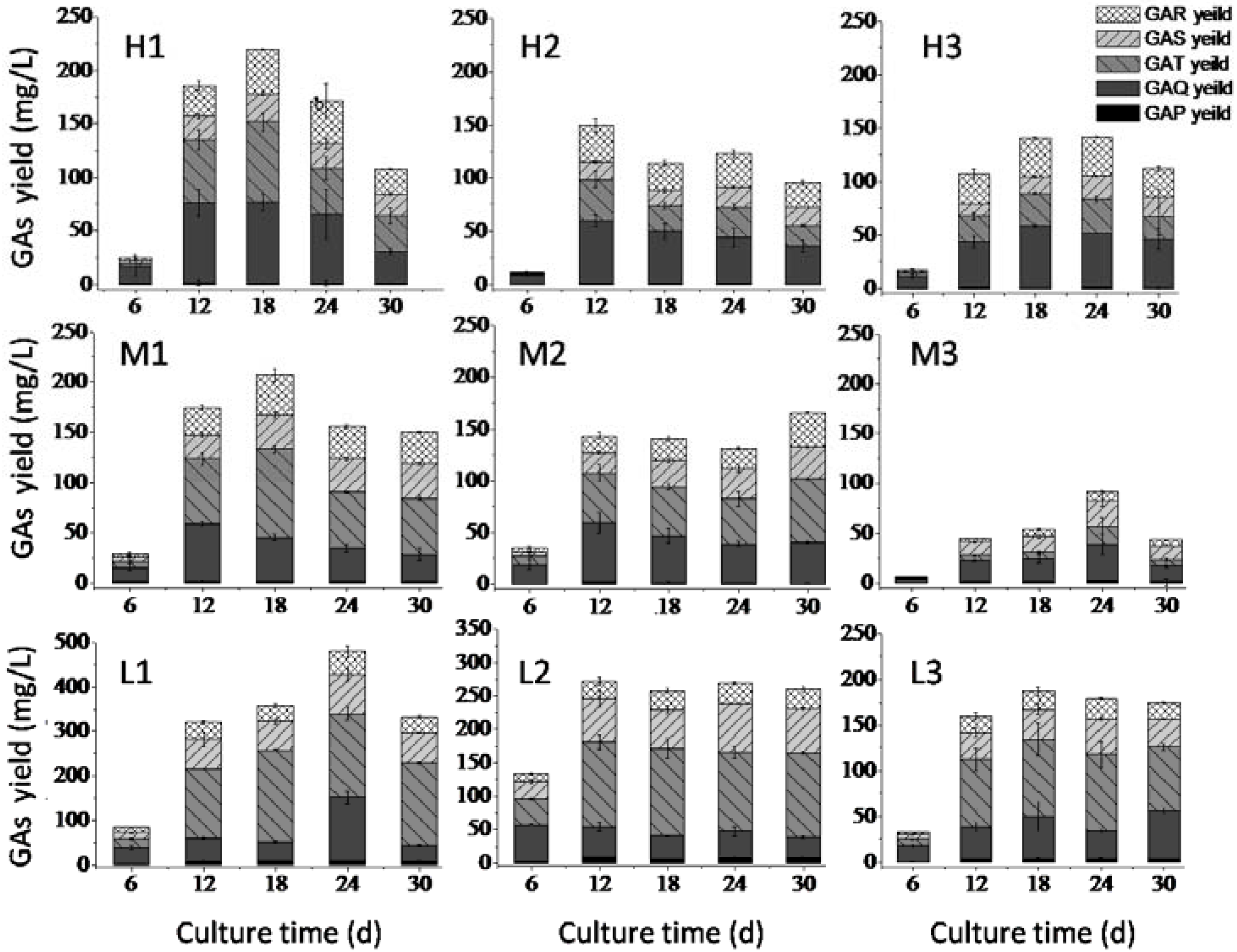
2.3. Effects of Nitrogen Limitation on GA Accumulation
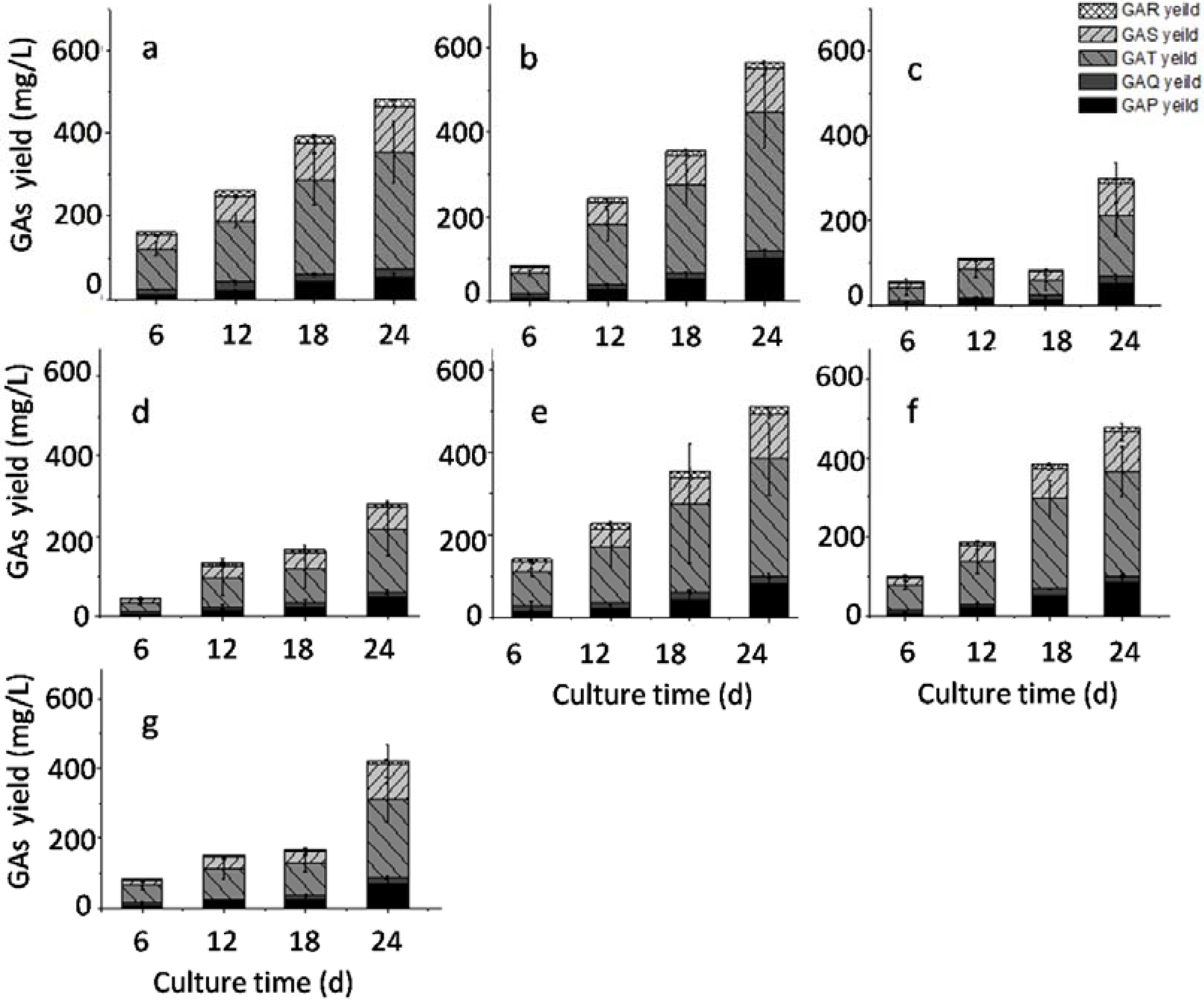
2.4. Effects of Glucose Concentration and Glucose Supply Strategy on GA Accumulation
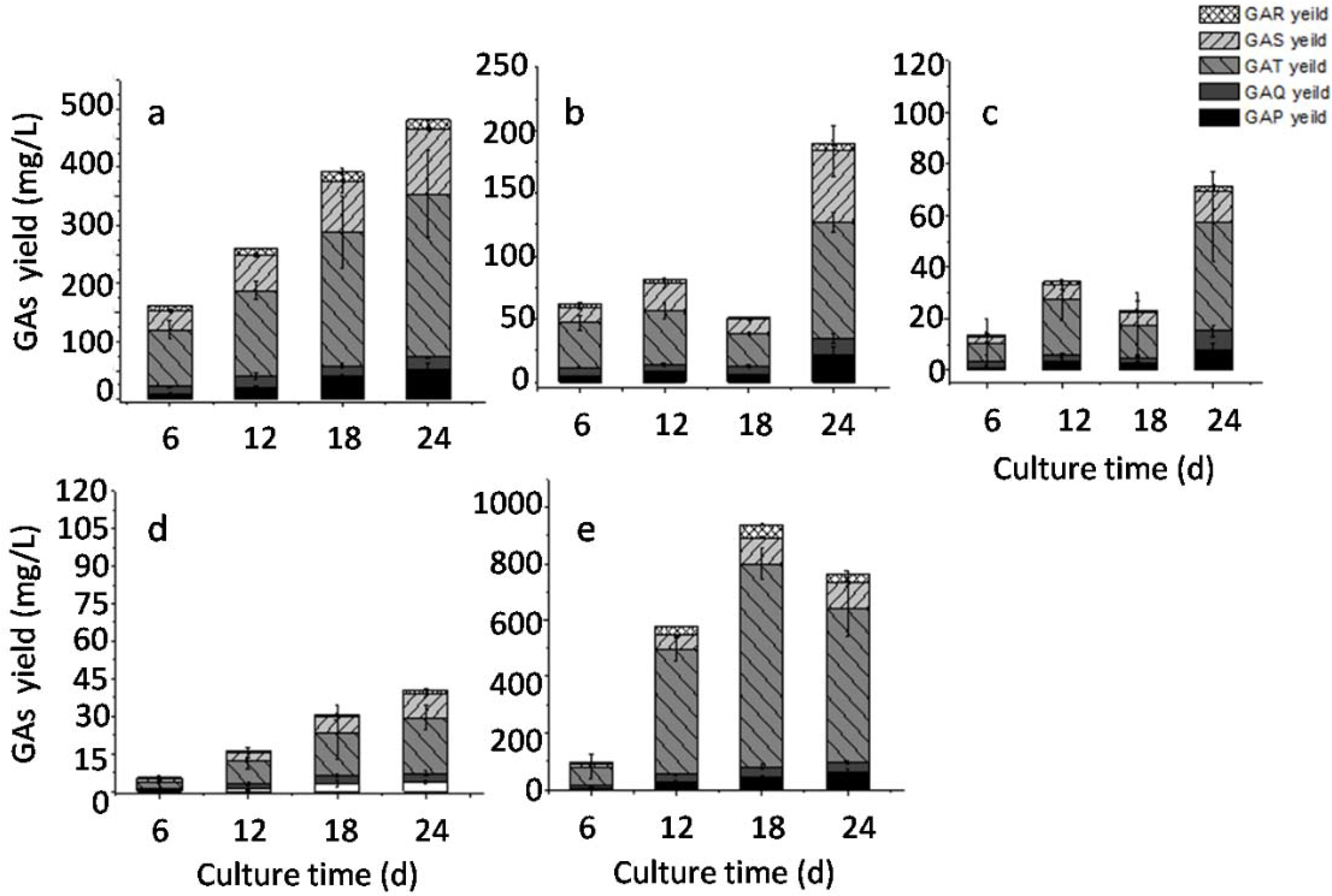
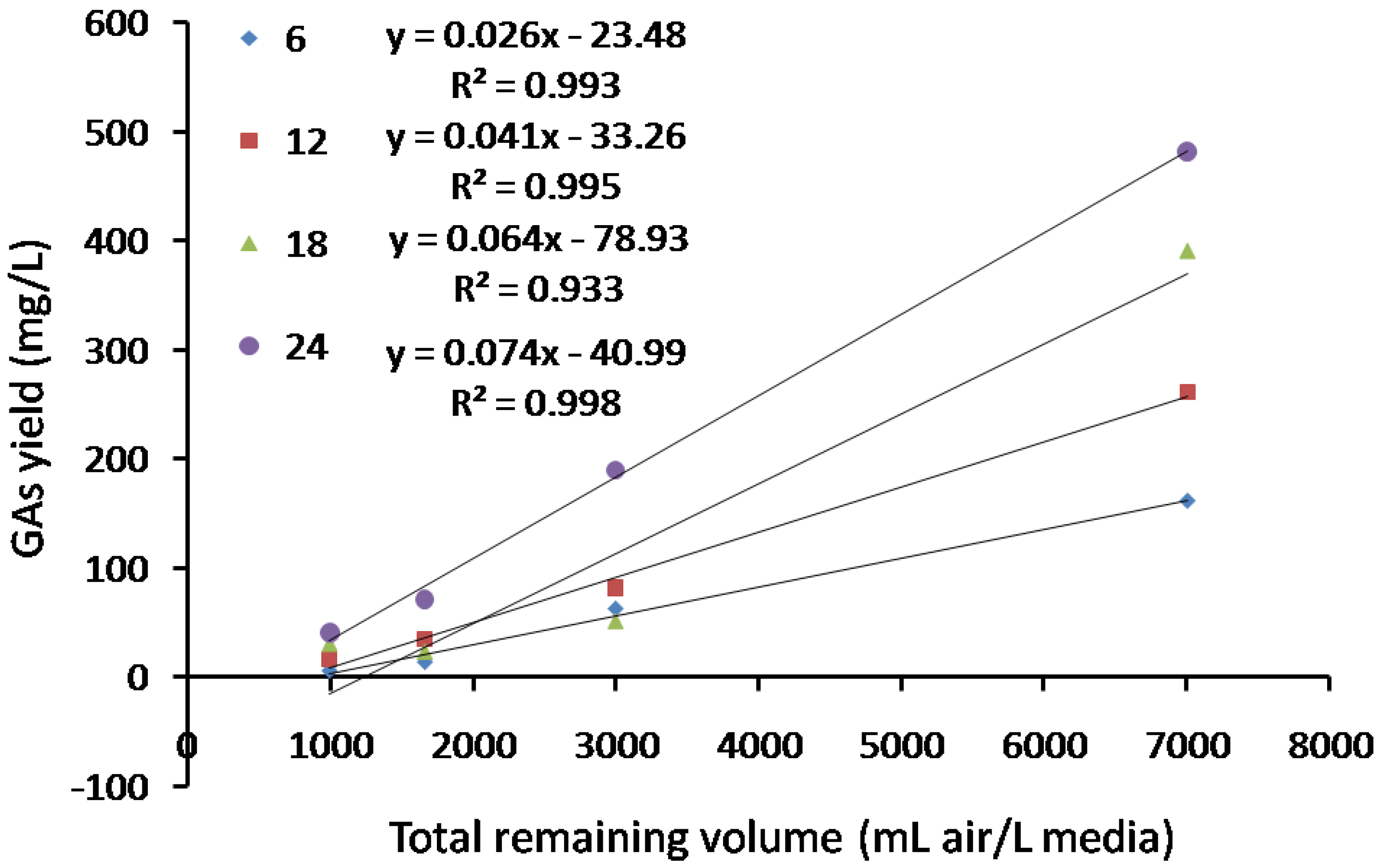
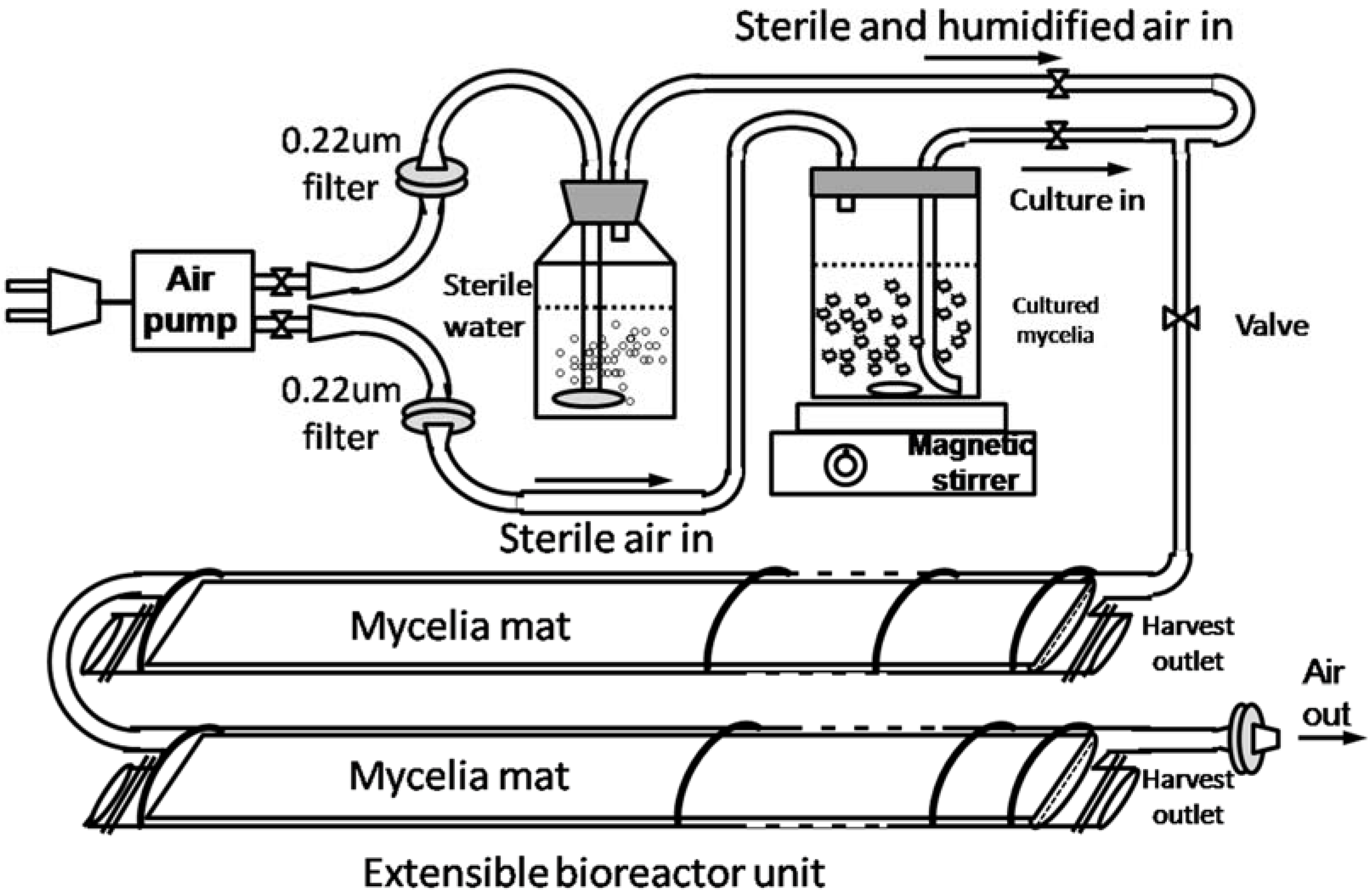
2.5. Effects of Static Culture Volume and Air Supply on GA Accumulation
3. Materials and Methods
3.1. Chemicals and Reagents
3.2. Strain, Culture Conditions and Sampling Method
3.2.1. Strain, Seed Culture and Sampling Conditions
3.2.2. Nitrogen Limitation
3.2.3. Glucose Concentration Optimization
3.2.4. Static Culture Volume Optimization
3.2.5. Air Supply in Static Liquid Culture
3.3. Isolation Procedure for GA-P, GA-Q, GA-T, GA-S, GA-R
3.4. HR-ESI-MS Analysis
3.5. Sample Preparation for Content Determination
3.6. HPLC Conditions and Standard Curve Preparation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Goyal, S.; Ramawat, K.G.; Mérillon, J.M. Fungal Metabolites; Springer International Publishing: Basel, Switzerland, 2016; pp. 1–27. ISBN 978-3-319-19456-1. [Google Scholar]
- Liu, R.M.; Li, Y.B.; Zhong, J.J. Cytotoxic and pro-apoptotic effects of novel ganoderic acid derivatives on human cervical cancer cells in vitro. Eur. J. Pharmacol. 2012, 681, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.S.; Guo, J.J.; Bao, J.L.; Li, X.W.; Chen, X.P.; Lu, J.J.; Wang, Y.T. Anti-cancer properties of triterpenoids isolated from Ganoderma lucidum—A review. Expert Opin. Investig. Drug 2013, 22, 981–992. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.H.; Liu, J.J.; Zhong, J.J. Ganoderic acid T inhibits tumor invasion in vitro and in vivo through inhibition of MMP expression. Pharmacol. Rep. 2010, 62, 150–163. [Google Scholar] [CrossRef]
- Liu, R.M.; Zhong, J.J. Ganoderic acid Mf and S induce mitochondria mediated apoptosis in human cervical carcinoma HeLa cells. Phytomedicine 2011, 18, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Hirotani, M.; Asaka, I.; Ino, C.; Furuya, T.; Shiro, M. Ganoderic acid derivatives and ergosta-4,7,22-triene-3,6-dione from Ganoderma lucidum. Phytochemistry 1987, 26, 2797–2803. [Google Scholar] [CrossRef]
- Li, Y.B.; Liu, R.M.; Zhong, J.J. A new ganoderic acid from Ganoderma lucidum mycelia and its stability. Fitoterapia 2013, 84, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Masahiko, I.; Panida, C.; Malipan, S.; Kannawat, D.; Thitiya, B.; Rattaket, C. Antitubercular lanostane triterpenes from cultures of the basidiomycete Ganoderma sp. BCC 16642. J. Nat. Prod. 2016, 79, 161–169. [Google Scholar]
- Zhang, Z.; Zhang, J.S.; Liu, Y.F.; Zhou, S.; Wang, J.Y.; Yu, H.J.; Tang, Q.J. Discussion on the determination of total triterpenoids in Ganoderma lucidum by spectrophotometry. Acta Agric. Shanghai 2016, 32, 61–65. [Google Scholar]
- Chen, Z.J.; Gu, J.X. Progress of studies on the main active substances and submerged fermentation technology of Ganoderma lucimum. Food Res. Dev. 2008, 39, 186–189. [Google Scholar]
- Huang, L.M.; Chen, C.W.; Chen, M.T.; Liu, Y.C.; Chen, K.D. Industrial Scale Process of Cultivating Ganoderma lucidum Mycelium. U.S. Patent 9820443B2, 21 November 2017. [Google Scholar]
- Fang, Q.H.; Zhong, J.J. Submerged fermentation of higher fungus Ganoderma lucidum for production of valuable bioactive metabolites-ganoderic acid and polysaccharide. Biochem. Eng. J. 2002, 10, 61–65. [Google Scholar] [CrossRef]
- Shen, W.; Hu, L.L.; Li, H.Y.; Hu, G.S. Studies on Adenosine Accumulation in Two-Stage Cultured Cordyceps sinensis Mycelium. Food Res. Dev. 2015, 36, 53–56. [Google Scholar]
- Xu, J.W.; Zhao, W.; Zhong, J.J. Biotechnological production and application of ganoderic acids. Appl. Microbiol. Biotechnol. 2010, 87, 457–466. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.Y.; Tsai, G.J.; Houng, J.Y. Optimization of the medium composition for the submerged culture of Ganoderma lucidum, by Taguchi array design and steepest ascent method. Enzym. Microb. Technol. 2006, 38, 407–414. [Google Scholar] [CrossRef]
- Zhang, W.; Tang, Y.J. A novel three-stage light irradiation strategy in the submerged fermentation of medicinal mushroom Ganoderma lucidum for the efficient production of ganoderic acid and Ganoderma polysaccharides. Biotechnol. Prog. 2008, 24, 1249–1261. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.X.; Li, Y.B.; Xu, J.W.; Wang, J.L.; Miao, X.L.; Tang, Y.J.; Gu, T.Y.; Zhong, J.J. Enhanced biosynthetic gene expressions and production of ganoderic acids in static liquid culture of Ganoderma lucidum under phenobarbital induction. Appl. Microbiol. Biotechnol. 2010, 86, 1367–1374. [Google Scholar] [CrossRef] [PubMed]
- Baabitskaia, V.G.; Shcherba, V.V.; Puchkova, T.A.; Smirnov, D.A. Polysaccharides of Ganoderma lucidum: Factors affecting their production. Appl. Biochem. Microbiol. 2005, 41, 194–199. [Google Scholar] [CrossRef]
- Shiao, M.S. Natural products of the medicinal fungus Ganoderma lucidum: Occurrence, biological activities, and pharmacological functions. Chem. Rec. 2003, 3, 172. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.N.; Chen, J.S.; Shiao, M.S.; Wang, C.T. Activation of human platelet phospholipases C and A2 by various oxygenated triterpenes. Eur. J. Pharmacol. 1994, 267, 33. [Google Scholar] [CrossRef]
- Hirotani, M.; Ino, C.; Furuya, T.; Shiro, M. Ganoderic acid T, S and R, new triterpenoids from the cultured mycelia of Ganoderma lucidum. Chem. Pharm. Bull. 1986, 34, 2282–2285. [Google Scholar] [CrossRef]
- Xu, J.W.; Zhong, J. Production of individual ganoderic acids and expression of biosynthetic genesin liquid static and shaking cultures of Ganoderma lucidum. Appl. Microbiol. Biotechnol. 2010, 85, 941–948. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Xu, J.W.; Zhong, J.J. Enhanced production of ganoderic acids in static liquid culture of Ganoderma lucidum under nitrogen-limiting conditions. Bioresour. Technol. 2011, 102, 8185–8190. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.H.; Liu, L.L.; Guo, X.F.; Li, Y.J.; Hou, B.C.; Fan, Q.L.; Wang, K.X.; Luo, Y.D.; Zhong, J.J. Sucrose fed-batch strategy enhanced biomass, polysaccharide, and ganoderic acids production in fermentation of Ganoderma lucidum 5.26. Bioprocess Biosyst. Eng. 2016, 39, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.X.; Zhong, J.J. Effect of oxygen concentration in gas phase on sporulation and individual ganodericacids accumulation in liquid static culture of Ganoderma lucidum. J. Biosci. Bioeng. 2010, 109, 37–40. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.X.; Tang, Y.J.; Zhong, J.J. Impact of oxygen level in gaseous phase on gene transcription and ganoderic acid biosynthesis in liquid static cultures of Ganoderma lucidum. Bioprocess Biosyst. Eng. 2010, 33, 683–690. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.-J.; Zhong, J.-J. Scale-up of a liquid static culture process for hyperproduction of ganoderic acid by the medicinal mushroom Ganoderma lucidum. Biotechnol. Prog. 2003, 19, 1842–1846. [Google Scholar] [CrossRef] [PubMed]
- Ha, D.T.; Loan, L.T.; Hung, T.M.; Han, L.V.; Khoi, N.M.; Dung, L.V.; Min, B.S.; Nguyen, N.P. An improved HPLC-DAD method for quantitative comparisons of triterpenes in Ganoderma lucidum and its five related species originating from Vietnam. Molecules 2015, 20, 1059–1077. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Rt (min) | Regression Equation | Linear Range (µg/mL) | R2 |
|---|---|---|---|---|
| GA-P | 22.1 | y = 6222x − 4019 | 1.224~122.4 | 0.999 |
| GA-Q | 27.2 | y =5446x − 12,150 | 2.88~288 | 0.999 |
| GA-T | 36 | y = 4056x + 3721 | 0.8~1000 | 0.999 |
| GA-S | 38.5 | y = 8163x + 5397 | 0.5536~415.2 | 0.999 |
| GA-R | 41.5 | y = 5948x − 6041 | 0.496~248 | 0.999 |
| Groups | H1 | H2 | H3 | M1 | M2 | M3 | L1 | L2 | L3 |
|---|---|---|---|---|---|---|---|---|---|
| Soybean powder (g/L) | 5 | 10 | 15 | 2.5 | 5 | 7.5 | 1.25 | 2.5 | 3.75 |
| Peptone (g/L) | 7.5 | 5 | 2.5 | 3.75 | 2.5 | 1.25 | 1.88 | 1.25 | 0.63 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, G.; Zhai, M.; Niu, R.; Xu, X.; Liu, Q.; Jia, J. Optimization of Culture Condition for Ganoderic Acid Production in Ganoderma lucidum Liquid Static Culture and Design of a Suitable Bioreactor. Molecules 2018, 23, 2563. https://doi.org/10.3390/molecules23102563
Hu G, Zhai M, Niu R, Xu X, Liu Q, Jia J. Optimization of Culture Condition for Ganoderic Acid Production in Ganoderma lucidum Liquid Static Culture and Design of a Suitable Bioreactor. Molecules. 2018; 23(10):2563. https://doi.org/10.3390/molecules23102563
Chicago/Turabian StyleHu, Gaosheng, Manhuayun Zhai, Rong Niu, Xiaoqiang Xu, Qian Liu, and Jingming Jia. 2018. "Optimization of Culture Condition for Ganoderic Acid Production in Ganoderma lucidum Liquid Static Culture and Design of a Suitable Bioreactor" Molecules 23, no. 10: 2563. https://doi.org/10.3390/molecules23102563
APA StyleHu, G., Zhai, M., Niu, R., Xu, X., Liu, Q., & Jia, J. (2018). Optimization of Culture Condition for Ganoderic Acid Production in Ganoderma lucidum Liquid Static Culture and Design of a Suitable Bioreactor. Molecules, 23(10), 2563. https://doi.org/10.3390/molecules23102563