
Two-Dimensional Fluorescence Lifetime Correlation Spectroscopy: Concepts and Applications


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Instrumentation of 2D FLCS
3. Analytical Method of 2D FLCS
3.1. Construction of a 2D Emission-Delay Correlation Map
3.2. Subtraction of Uncorrelated Photon Pairs
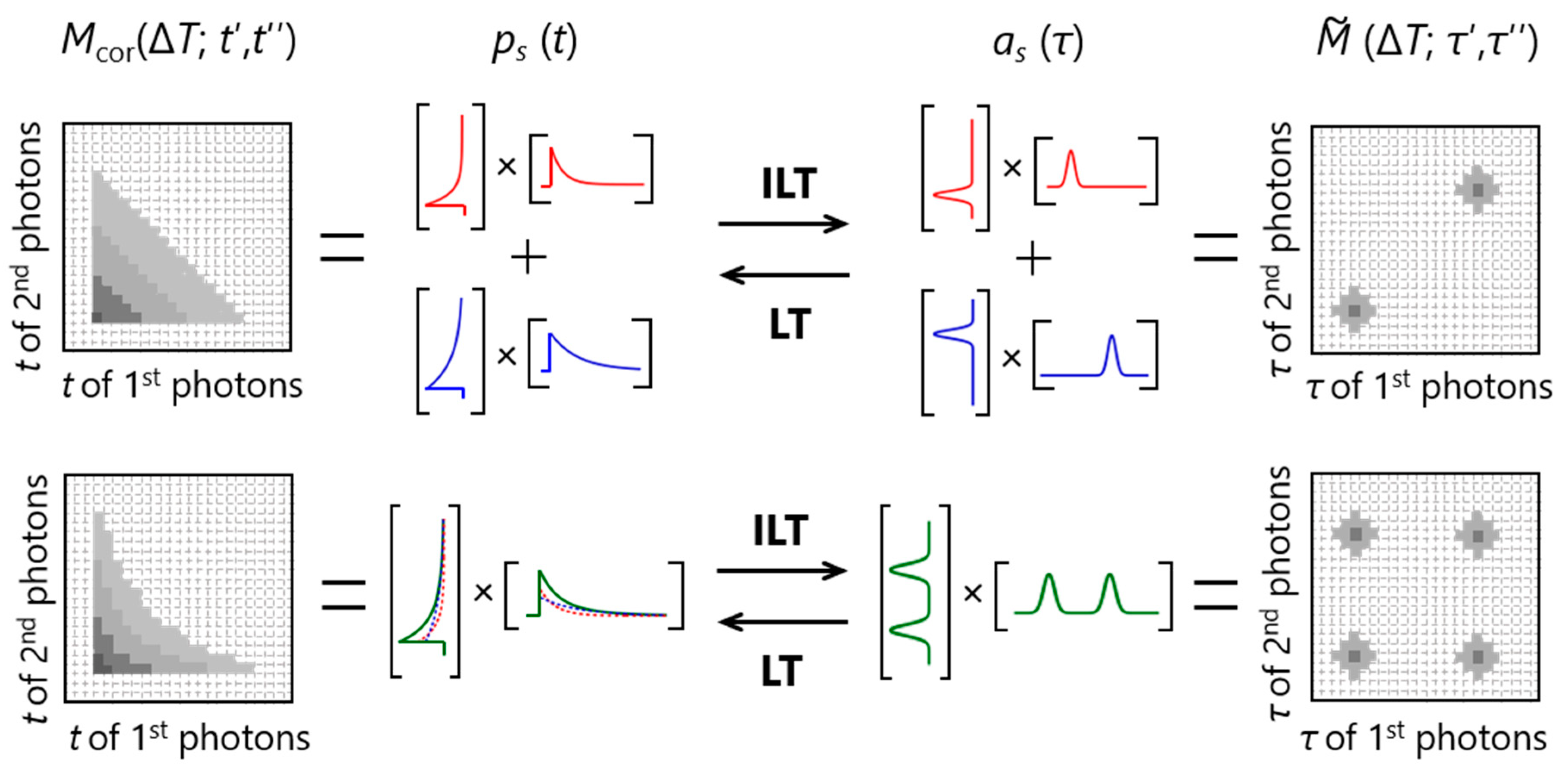
3.3. Inverse Laplace Transform with the Help of 2D Maximum Entropy Method
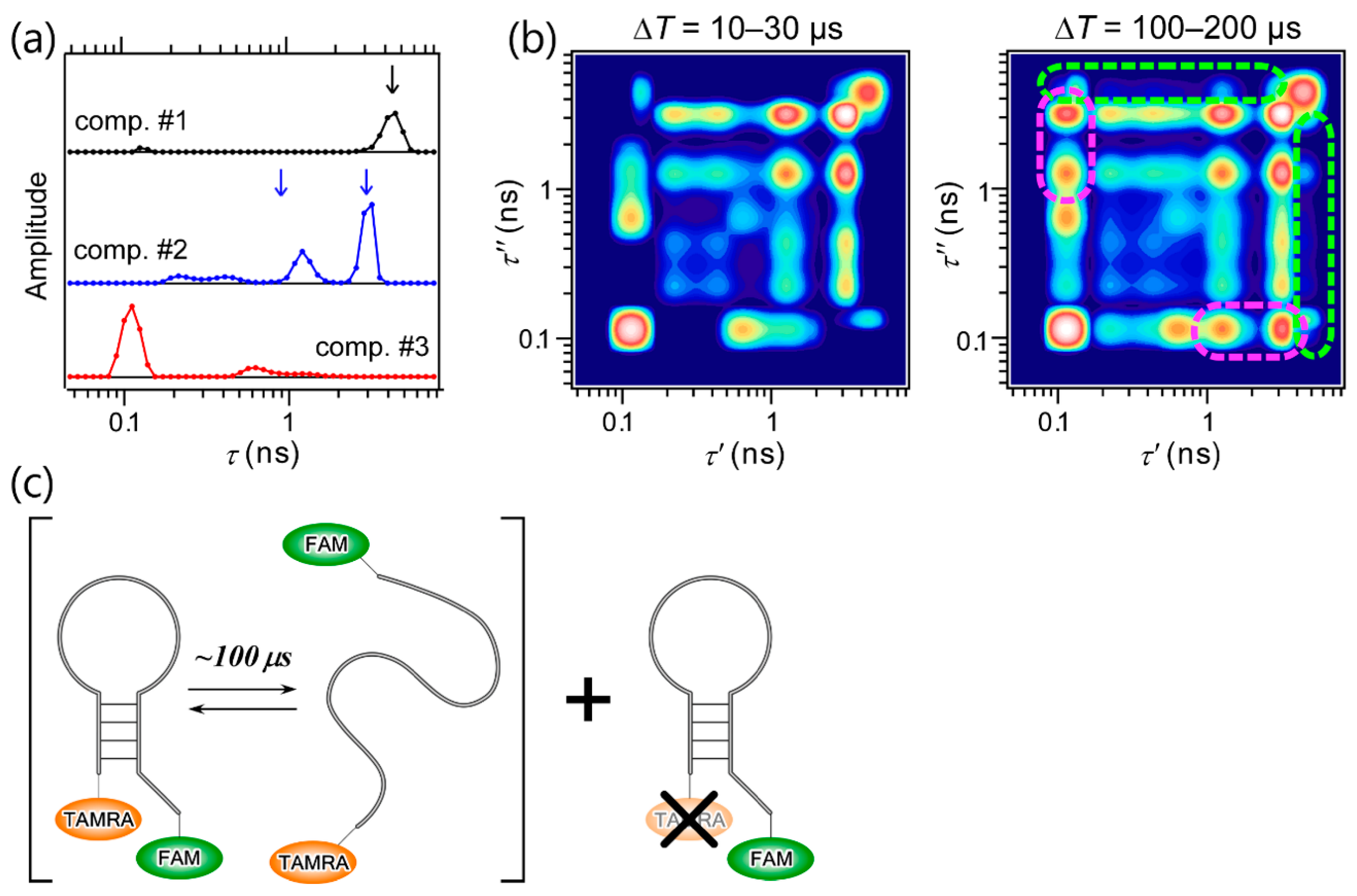
4. Application of 2D FLCS
5. Future Perspectives
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Elson, E.L.; Magde, D. Fluorescence correlation spectroscopy. I. Conceptual basis and theory. Biopolymers 1974, 13, 1–27. [Google Scholar] [CrossRef]
- Dertinger, T.; Pacheco, V.; von der Hocht, I.; Hartmann, R.; Gregor, I.; Enderlein, J. Two-focus fluorescence correlation spectroscopy: A new tool for accurate and absolute diffusion measurements. Chemphyschem 2007, 8, 433–443. [Google Scholar] [CrossRef] [PubMed]
- Nettels, D.; Hoffmann, A.; Schuler, B. Unfolded protein and peptide dynamics investigated with single-molecule fret and correlation spectroscopy from picoseconds to seconds. J. Phys. Chem. B 2008, 112, 6137–6146. [Google Scholar] [CrossRef] [PubMed]
- Elson, E.L. Fluorescence correlation spectroscopy: Past, present, future. Biophys. J. 2011, 101, 2855–2870. [Google Scholar] [CrossRef] [PubMed]
- Widengren, J.; Mets, U.; Rigler, R. Fluorescence correlation spectroscopy of triplet-states in solution: A theoretical and experimental study. J. Phys. Chem. 1995, 99, 13368–13379. [Google Scholar] [CrossRef]
- Soranno, A.; Buchli, B.; Nettels, D.; Cheng, R.R.; Muller-Spath, S.; Pfeil, S.H.; Hoffmann, A.; Lipman, E.A.; Makarov, D.E.; Schuler, B. Quantifying internal friction in unfolded and intrinsically disordered proteins with single-molecule spectroscopy. Pros. Natl. Acad. Sci. USA 2012, 109, 17800–17806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krieger, J.W.; Singh, A.P.; Bag, N.; Garbe, C.S.; Saunders, T.E.; Langowski, J.; Wohland, T. Imaging fluorescence (cross-) correlation spectroscopy in live cells and organisms. Nat. Protoc. 2015, 10, 1948–1974. [Google Scholar] [CrossRef] [PubMed]
- Mikuni, S.; Yamamoto, J.; Horio, T.; Kinjo, M. Negative correlation between the diffusion coefficient and transcriptional activity of the glucocorticoid receptor. Int. J. Mol. Sci. 2017, 18, 1855. [Google Scholar] [CrossRef] [PubMed]
- Bacia, K.; Kim, S.A.; Schwille, P. Fluorescence cross-correlation spectroscopy in living cells. Nat. Methods 2006, 3, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Hanson, K.M.; Davis, S.K.; Bardeen, C.J. Two-photon standing-wave fluorescence correlation spectroscopy. Opt. Lett. 2007, 32, 2121–2123. [Google Scholar] [CrossRef] [PubMed]
- Mueller, V.; Honigmann, A.; Ringemann, C.; Medda, R.; Schwarzmann, G.; Eggeling, C. FCS in STED microscopy: Studying the nanoscale of lipid membrane dynamics. Method Enzymol. 2013, 519, 1–38. [Google Scholar]
- Benda, A.; Kapusta, P.; Hof, M.; Gaus, K. Fluorescence spectral correlation spectroscopy (FSCS) for probes with highly overlapping emission spectra. Opt. Express 2014, 22, 2973–2988. [Google Scholar] [CrossRef] [PubMed]
- Vicidomini, G.; Ta, H.; Honigmann, A.; Mueller, V.; Clausen, M.P.; Waithe, D.; Galiani, S.; Sezgin, E.; Diaspro, A.; Hell, S.W.; et al. STED-FLCS: An advanced tool to reveal spatiotemporal heterogeneity of molecular membrane dynamics. Nano Lett. 2015, 15, 5912–5918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bohmer, M.; Wahl, M.; Rahn, H.J.; Erdmann, R.; Enderlein, J. Time-resolved fluorescence correlation spectroscopy. Chem. Phys. Lett. 2002, 353, 439–445. [Google Scholar] [CrossRef]
- Enderlein, J.; Gregor, I. Using fluorescence lifetime for discriminating detector afterpulsing in fluorescence correlation spectroscopy. Rev. Sci. Instrum. 2005, 76, 033102. [Google Scholar] [CrossRef] [Green Version]
- Kapusta, P.; Wahl, M.; Benda, A.; Hof, M.; Enderlein, J. Fluorescence lifetime correlation spectroscopy. J. Fluoresc. 2007, 17, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Humpolickova, J.; Beranova, L.; Stepanek, M.; Benda, A.; Prochazka, K.; Hof, M. Fluorescence lifetime correlation spectroscopy reveals compaction mechanism of 10 and 49 kbp DNA and differences between polycation and cationic surfactant. J. Phys. Chem. B 2008, 112, 16823–16829. [Google Scholar] [CrossRef] [PubMed]
- Felekyan, S.; Kalinin, S.; Sanabria, H.; Valeri, A.; Seidel, C.A.M. Filtered FCS: Species auto- and cross-correlation functions highlight binding and dynamics in biomolecules. Chemphyschem 2012, 13, 1036–1053. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.; Isbaner, S.; Veiga-Gutierrez, M.; Gregor, I.; Enderlein, J.; Karedla, N. Quantifying microsecond transition times using fluorescence lifetime correlation spectroscopy. J. Phys. Chem. Lett. 2017, 8, 6022–6028. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Xie, X.S. Probing single-molecule dynamics photon by photon. J. Chem. Phys. 2002, 117, 10965–10979. [Google Scholar] [CrossRef]
- Yang, H.; Xie, X.S. Statistical approaches for probing single-molecule dynamics photon-by-photon. Chem. Phys. 2002, 284, 423–437. [Google Scholar] [CrossRef]
- Ishii, K.; Tahara, T. Resolving inhomogeneity using lifetime-weighted fluorescence correlation spectroscopy. J. Phys. Chem. B 2010, 114, 12383–12391. [Google Scholar] [CrossRef] [PubMed]
- Ishii, K.; Tahara, T. Extracting decay curves of the correlated fluorescence photons measured in fluorescence correlation spectroscopy. Chem. Phys. Lett. 2012, 519–520, 130–133. [Google Scholar] [CrossRef]
- Ishii, K.; Tahara, T. Two-dimensional fluorescence lifetime correlation spectroscopy. 1. Principle. J. Phys. Chem. B 2013, 117, 11414–11422. [Google Scholar] [CrossRef] [PubMed]
- Ishii, K.; Tahara, T. Two-dimensional fluorescence lifetime correlation spectroscopy. 2. Application. J. Phys. Chem. B 2013, 117, 11423–11432. [Google Scholar] [CrossRef] [PubMed]
- Kapusta, P.; Machan, R.; Benda, A.; Hof, M. Fluorescence lifetime correlation spectroscopy (FLCS): Concepts, applications and outlook. Int. J. Mol. Sci. 2012, 13, 12890–12910. [Google Scholar] [CrossRef] [PubMed]
- Machan, R.; Kapusta, P.; Hof, M. Statistical filtering in fluorescence microscopy and fluorescence correlation spectroscopy. Anal. Bioanal. Chem. 2014, 406, 4797–4813. [Google Scholar] [CrossRef] [PubMed]
- Basit, H.; Lopez, S.G.; Keyes, T.E. Fluorescence correlation and lifetime correlation spectroscopy applied to the study of supported lipid bilayer models of the cell membrane. Methods 2014, 68, 286–299. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.; Karedla, N.; Thiele, J.C.; Gregor, I.; Enderlein, J. Fluorescence lifetime correlation spectroscopy: Basics and applications. Methods 2018, 140, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Kapusta, P.W.M.; Erdmann, R. Advanced Photon Counting: Applications, Methods, Instrumentation; Springer: New York, NY, USA, 2015; Volume 15. [Google Scholar]
- Otosu, T.; Yamaguchi, S. Total internal reflection two-dimensional fluorescence lifetime correlation spectroscopy. J. Phys. Chem. B 2018, 122, 5758–5764. [Google Scholar] [CrossRef] [PubMed]
- Burstyn, H.C. Afterpulsing effects in photon-correlation experiments. Rev. Sci. Instrum. 1980, 51, 1431–1433. [Google Scholar] [CrossRef]
- Ishii, K.; Tahara, T. Correction of the afterpulsing effect in fluorescence correlation spectroscopy using time symmetry analysis. Opt. Express 2015, 23, 32387–32400. [Google Scholar] [CrossRef] [PubMed]
- Brochon, J.C. Maximum-entropy method of data-analysis in time-resolved spectroscopy. Methods Enzymol. 1994, 240, 262–311. [Google Scholar] [PubMed]
- Lindorff-Larsen, K.; Piana, S.; Dror, R.O.; Shaw, D.E. How fast-folding proteins fold. Science 2011, 334, 517–520. [Google Scholar] [CrossRef] [PubMed]
- Piana, S.; Lindorff-Larsen, K.; Shaw, D.E. Atomic-level description of ubiquitin folding. Proc. Natl. Acad. Sci. USA 2013, 110, 5915–5920. [Google Scholar] [CrossRef] [PubMed]
- Otosu, T.; Ishii, K.; Tahara, T. Microsecond protein dynamics observed at the single-molecule level. Nat. Commun. 2015, 6, 7685. [Google Scholar] [CrossRef] [PubMed]
- Goto, Y.; Hagihara, Y.; Hamada, D.; Hoshino, M.; Nishii, I. Acid-induced unfolding and refolding transitions of cytochrome c: A three-state mechanism in H2O and D2O. Biochemistry 1993, 32, 11878–11885. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.W.; Sosnick, T.R.; Mayne, L.; Englander, S.W. Protein-folding intermediates: Native-state hydrogen exchange. Science 1995, 269, 192–197. [Google Scholar] [CrossRef] [PubMed]
- Akiyama, S.; Takahashi, S.; Kimura, T.; Ishimori, K.; Morishima, I.; Nishikawa, Y.; Fujisawa, T. Conformational landscape of cytochrome c folding studied by microsecond-resolved small-angle X-Ray scattering. Proc. Natl. Acad. Sci. USA 2002, 99, 1329–1334. [Google Scholar] [CrossRef] [PubMed]
- Perroud, T.D.; Bokoch, M.P.; Zare, R.N. Cytochrome c conformations resolved by the photon counting histogram: Watching the alkaline transition with single-molecule sensitivity. Proc. Natl. Acad. Sci. USA 2005, 102, 17570–17575. [Google Scholar] [CrossRef] [PubMed]
- Werner, J.H.; Joggerst, R.; Dyer, R.B.; Goodwin, P.M. A two-dimensional view of the folding energy landscape of cytochrome c. Proc. Natl. Acad. Sci. USA 2006, 103, 11130–11135. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, S.; Yane, A.; Nakashima, S.; Hashida, M.; Fujita, M.; Goto, Y.; Takahashi, S. A rapid flow mixer with 11 µs mixing time microfabricated by a pulsed-laser ablation technique: Observation of a barrier-limited collapse in cytochrome c folding. J. Am. Chem. Soc. 2007, 129, 3840–3841. [Google Scholar] [CrossRef] [PubMed]
- Otosu, T.; Ishii, K.; Oikawa, H.; Arai, M.; Takahashi, S.; Tahara, T. Highly heterogeneous nature of the native and unfolded states of the B domain of protein a revealed by two-dimensional fluorescence lifetime correlation spectroscopy. J. Phys. Chem. B 2017, 121, 5463–5473. [Google Scholar] [CrossRef] [PubMed]
- Gouda, H.; Torigoe, H.; Saito, A.; Sato, M.; Arata, Y.; Shimada, I. Three-dimensional solution structure of the B domain of staphylococcal protein a: Comparisons of the solution and crystal structures. Biochemistry 1992, 31, 9665–9672. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.W.; Karimi, A.; Dyson, H.J.; Wright, P.E. Absence of a stable intermediate on the folding pathway of protein A. Protein Sci. 1997, 6, 1449–1457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, S.; Religa, T.L.; Daggett, V.; Fersht, A.R. Testing protein-folding simulations by experiment: B domain of protein A. Proc. Natl. Acad. Sci. USA 2004, 101, 6952–6956. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Lerner, E.; Sato, S.; Amir, D.; Haas, E.; Fersht, A.R. Time-resolved fluorescence resonance energy transfer study shows a compact denatured state of the B domain of protein A. Biochemistry 2009, 48, 3468–3476. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Sato, S.; Sharpe, T.D.; Ying, L.; Fersht, A.R. Distinguishing between cooperative and unimodal downhill protein folding. Proc. Natl. Acad. Sci. USA 2007, 104, 123–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oikawa, H.; Suzuki, Y.; Saito, M.; Kamagata, K.; Arai, M.; Takahashi, S. Microsecond dynamics of an unfolded protein by a line confocal tracking of single molecule fluorescence. Sci. Rep. 2013, 3, 2151. [Google Scholar] [CrossRef] [PubMed]
- Oikawa, H.; Kamagata, K.; Arai, M.; Takahashi, S. Complexity of the folding transition of the B domain of protein A revealed by the high-speed tracking of single-molecule fluorescence time series. J. Phys. Chem. B 2015, 119, 6081–6091. [Google Scholar] [CrossRef] [PubMed]
- Rossow, M.J.; Sasaki, J.M.; Digman, M.A.; Gratton, E. Raster image correlation spectroscopy in live cells. Nat. Protoc. 2010, 5, 1761–1774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Digman, M.A.; Caiolfa, V.R.; Zamai, M.; Gratton, E. The phasor approach to fluorescence lifetime imaging analysis. Biophys. J. 2008, 94, 14–16. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.-H.; Ishii, K.; Tahara, T. RNA and DNA hairpin dynamics studied by temperature-controlled 2D fluorescence lifetime correlation spectroscopy. Proc. Asian Spectrosc. Conf. 2017, 92. [Google Scholar]
- Otosu, T.; Yamaguchi, S. Quantifying the diffusion of lipids in the proximal/distal leaflets of a supported lipid bilayer by two-dimensional fluorescence lifetime correlation spectroscopy. J. Phys. Chem. B 2018. [Google Scholar] [CrossRef] [PubMed]
- Castellana, E.T.; Cremer, P.S. Solid supported lipid bilayers: From biophysical studies to sensor design. Surf. Sci. Rep. 2006, 61, 429–444. [Google Scholar] [CrossRef]
- Allhusen, J.S.; Conboy, J.C. The ins and outs of lipid flip-flop. Accounts Chem. Res. 2017, 50, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.F.; Granick, S. Lipid diffusion compared in outer and inner leaflets of planar supported bilayers. J. Chem. Phys. 2005, 123, 211104. [Google Scholar] [CrossRef] [PubMed]
- Hetzer, M.; Heinz, S.; Grage, S.; Bayerl, T.M. Asymmetric molecular friction in supported phospholipid bilayers revealed by NMR measurements of lipid diffusion. Langmuir 1998, 14, 982–984. [Google Scholar] [CrossRef]
- Schoch, R.L.; Barel, I.; Brown, F.L.H.; Haran, G. Lipid diffusion in the distal and proximal leaflets of supported lipid bilayer membranes studied by single particle tracking. J. Chem. Phys. 2018, 148, 123333. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |






© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Otosu, T.; Yamaguchi, S. Two-Dimensional Fluorescence Lifetime Correlation Spectroscopy: Concepts and Applications. Molecules 2018, 23, 2972. https://doi.org/10.3390/molecules23112972
Otosu T, Yamaguchi S. Two-Dimensional Fluorescence Lifetime Correlation Spectroscopy: Concepts and Applications. Molecules. 2018; 23(11):2972. https://doi.org/10.3390/molecules23112972
Chicago/Turabian StyleOtosu, Takuhiro, and Shoichi Yamaguchi. 2018. "Two-Dimensional Fluorescence Lifetime Correlation Spectroscopy: Concepts and Applications" Molecules 23, no. 11: 2972. https://doi.org/10.3390/molecules23112972
APA StyleOtosu, T., & Yamaguchi, S. (2018). Two-Dimensional Fluorescence Lifetime Correlation Spectroscopy: Concepts and Applications. Molecules, 23(11), 2972. https://doi.org/10.3390/molecules23112972