Coptisine Suppresses Mast Cell Degranulation and Ovalbumin-Induced Allergic Rhinitis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
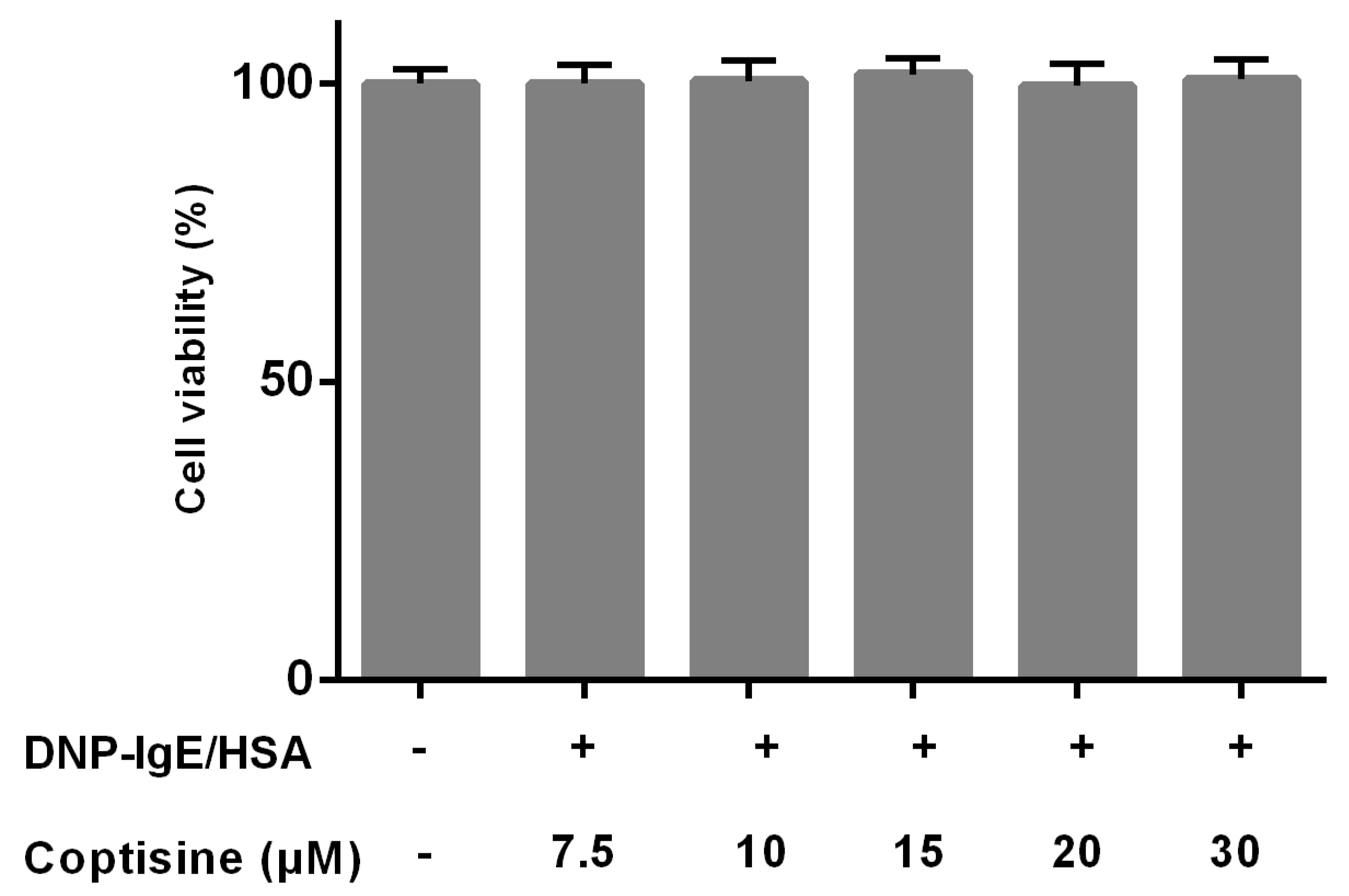
2.1. Effect of Coptisine on Cell Viability
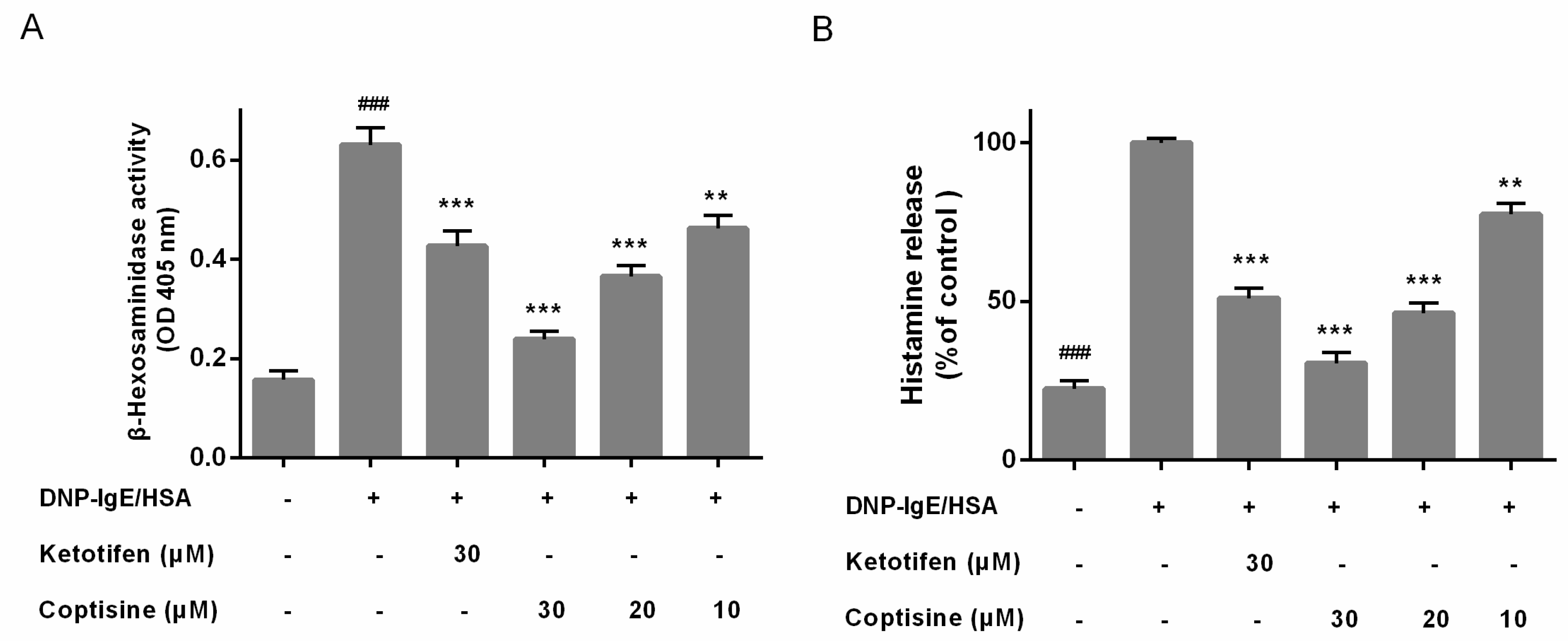
2.2. Effect of Coptisine on the Levels of β-Hexosaminidase and Histamine from Mast Cells
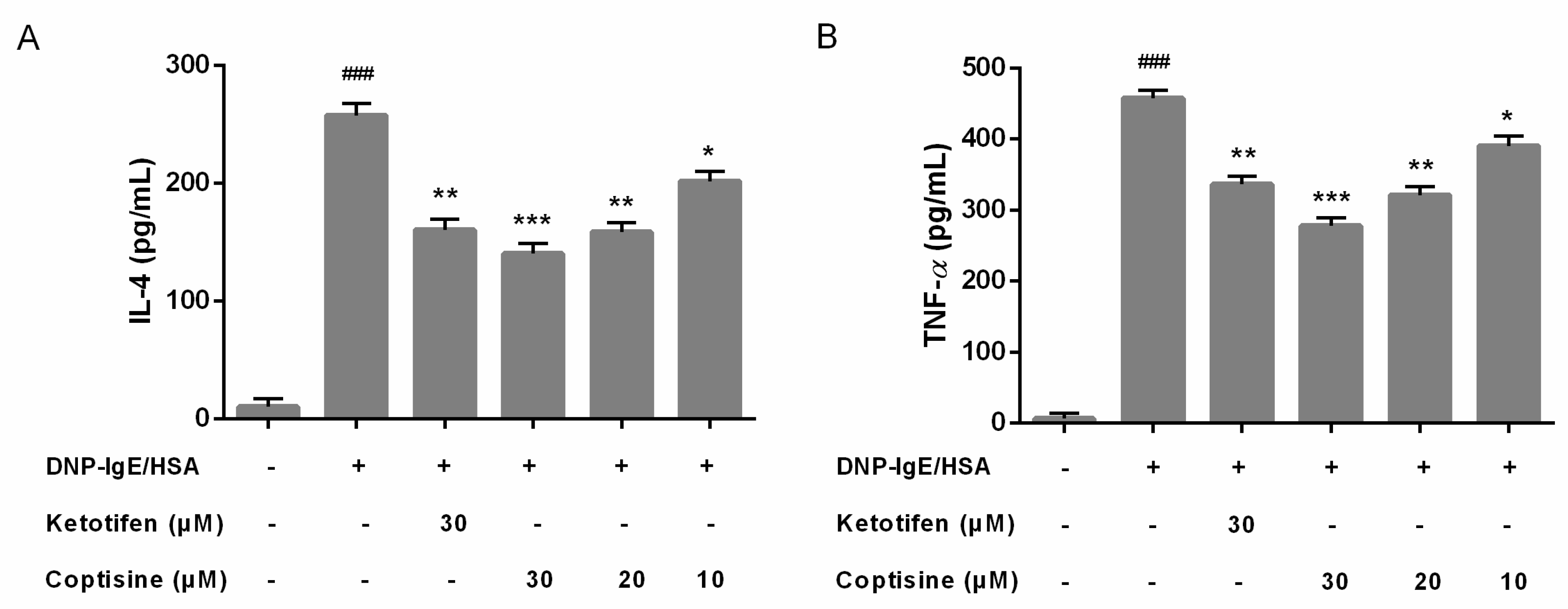
2.3. Effect of Coptisine on IL-4, TNF-α Levels in DNP-IgE/HSA-Stimulated RBL-2H3 Cells
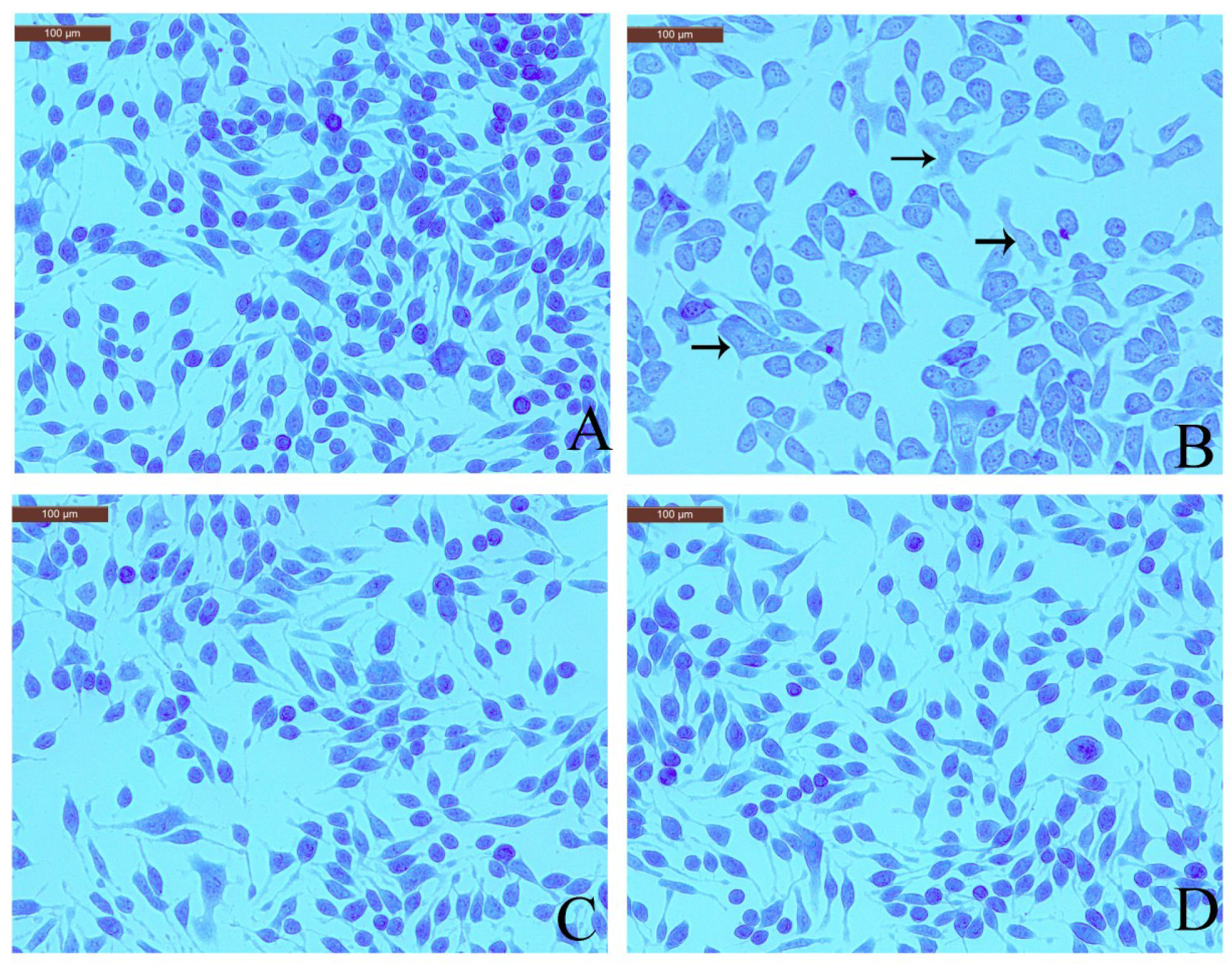
2.4. Effect of Coptisine Granule Release by DNP-IgE/HSA-Stimulated RBL-2H3 Cells
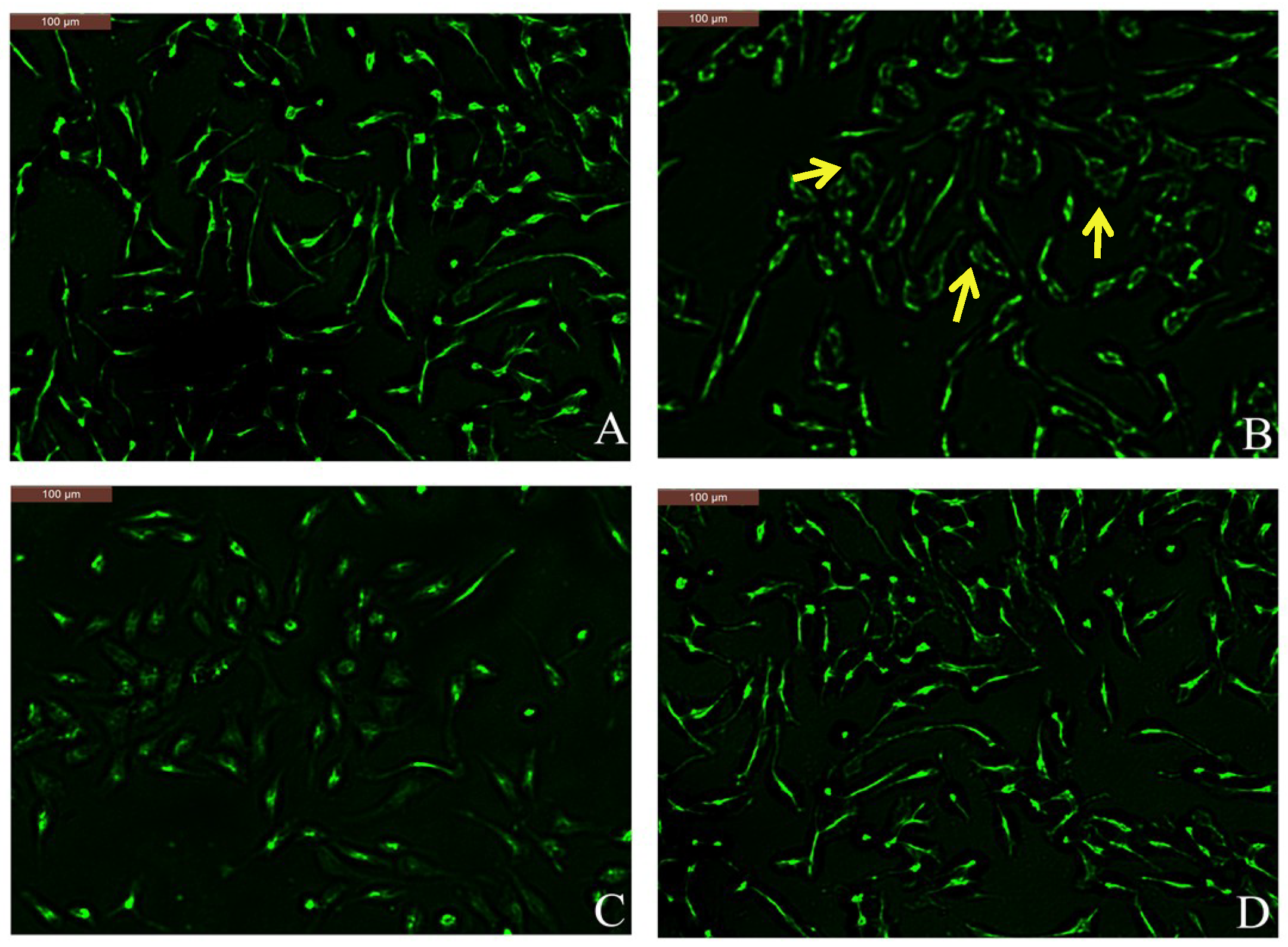
2.5. Effect of Coptisine on F-Actin Rearrangement in RBL-2H3 Cells
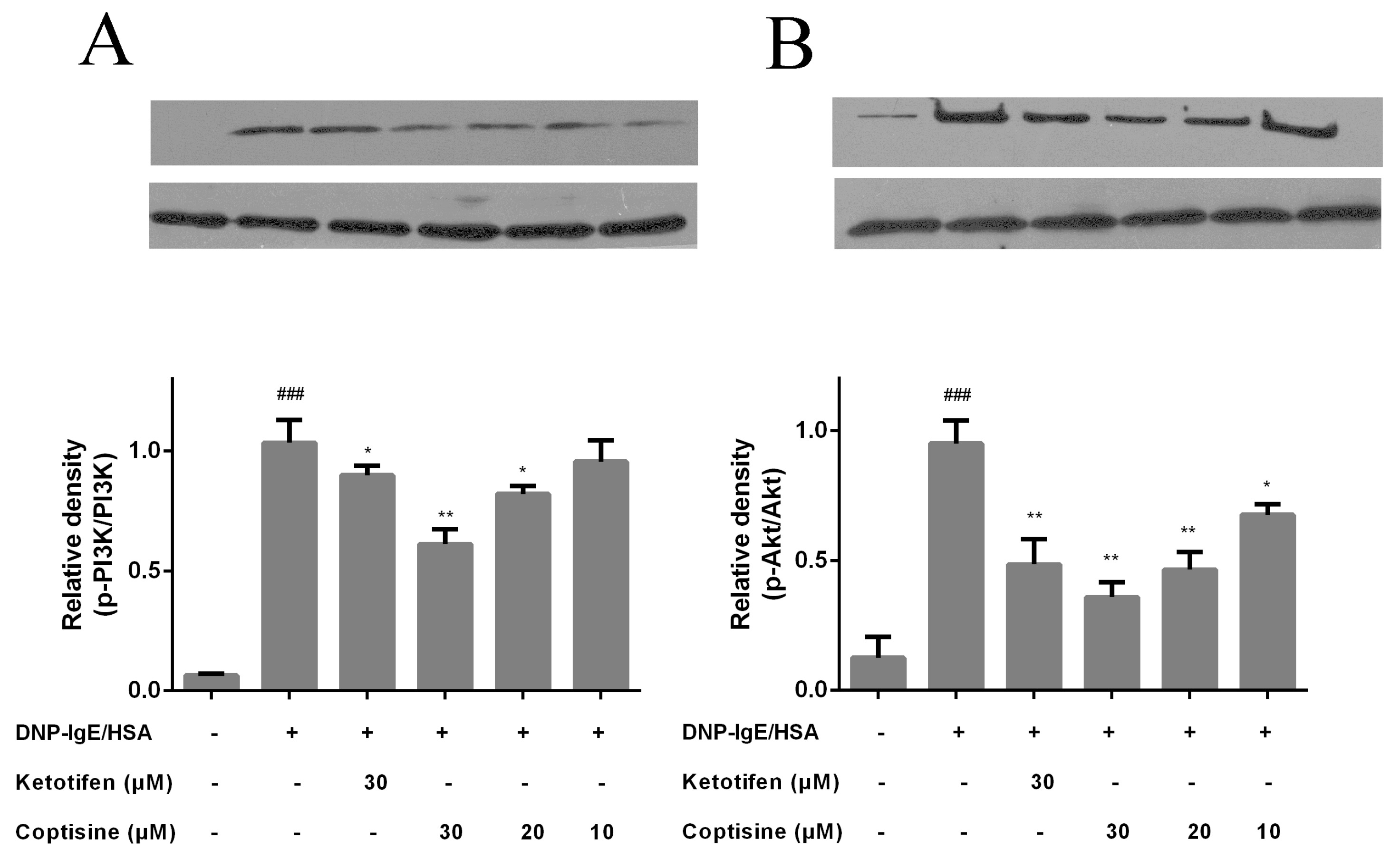
2.6. Effect of Coptisine on PI3K/Akt Signaling in RBL-2H3 Cells
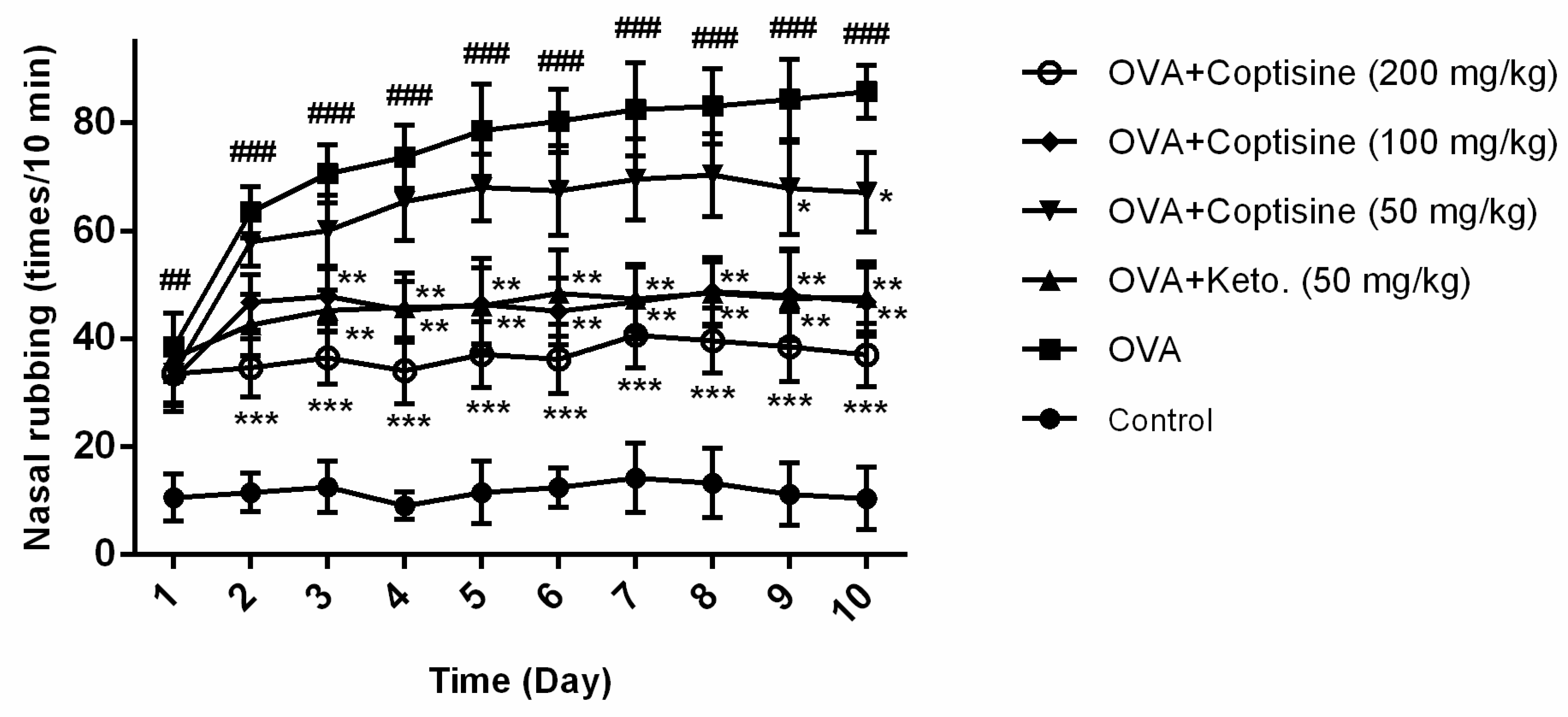
2.7. Effect of Coptisine on the Number of Occurrences of Nasal Rubbing in OVA Induced AR Mice
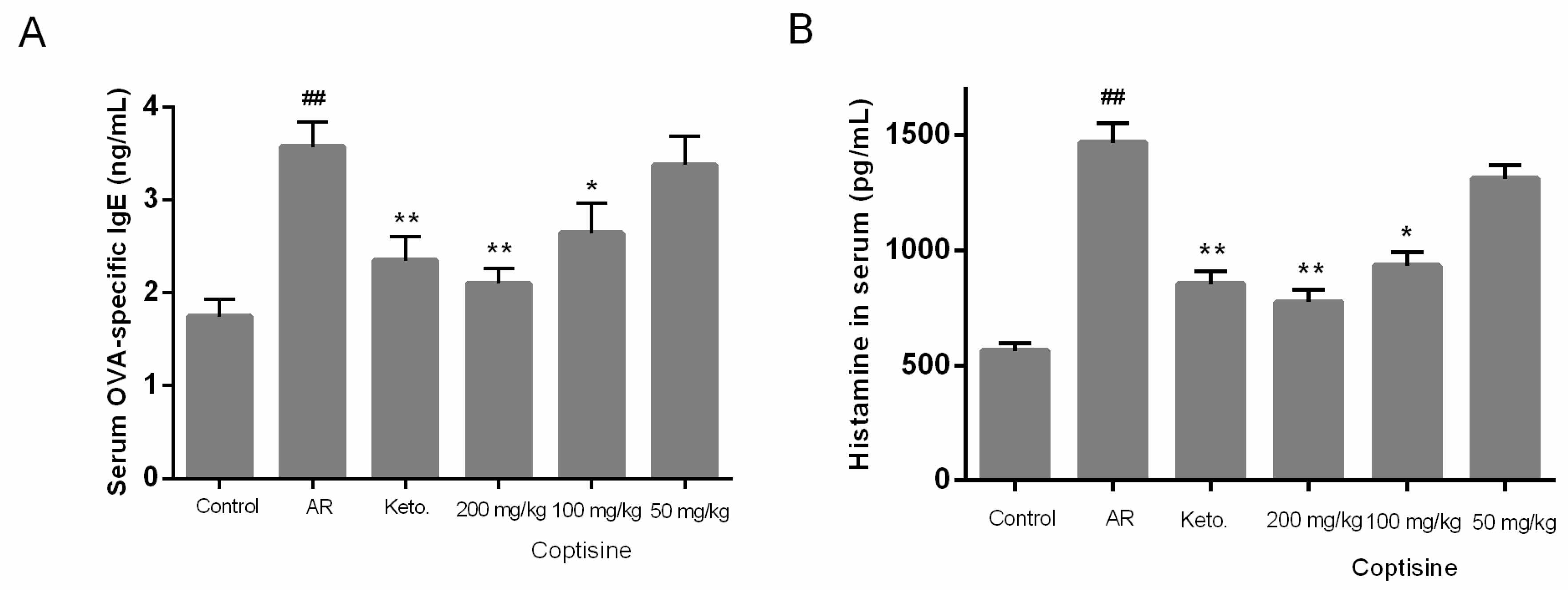
2.8. Effect of Coptisine on OVA-Specific IgE, Histamine Levels in OVA Induced AR Mice
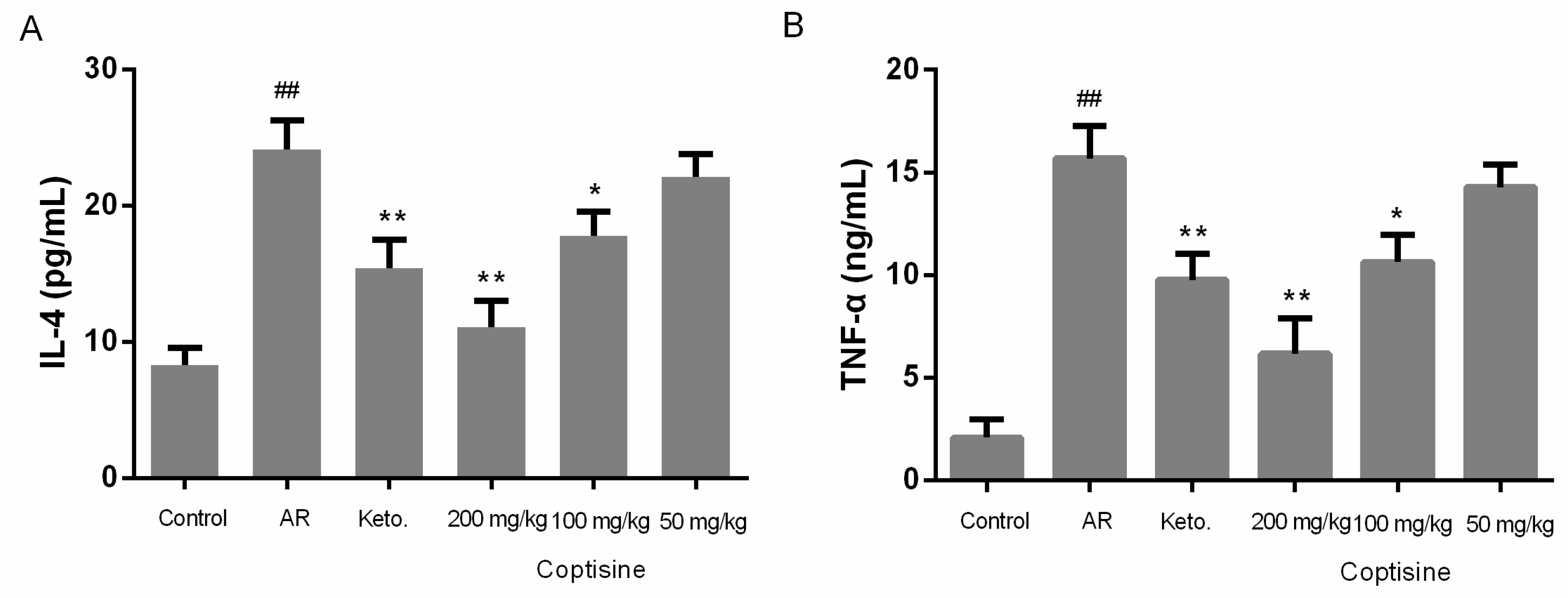
2.9. Effect of Coptisine on IL-4 and TNF-α Levels in OVA Induced AR Mice
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Animals
4.3. Cell Viability Assay
4.4. β-Hexosaminidase and Histamine Release Assay
4.5. Inflammatory Cytokines Assay
4.6. Toluidine Blue Staining
4.7. F-Actin Microfilament Staining
4.8. Western Blot
4.9. OVA-Induced AR in Mice
4.10. Statistical Analyses
Author Contributions
Conflicts of Interest
References
- Matsuoka, D.; Mizutani, N.; Sae-Wong, C.; Yoshino, S. Allergen-specific regulation of allergic rhinitis in mice by intranasal exposure to IgG1 monoclonal antibody Fab fragments against pathogenic allergen. Immunol. Lett. 2014, 161, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Nam, S.Y.; Chung, C.K.; Seo, J.H.; Rah, S.Y.; Kim, H.M.; Jeong, H.J. The therapeutic efficacy of α-pinene in an experimental mouse model of allergic rhinitis. Int. Immunopharmacol. 2014, 23, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Won, J.H.; Jung, J.K.; Weon, C.C.; Kang, J.S.; Park, Y.K. Antiallergic effect of KOB03, a polyherbal medicine, on mast cell-mediated allergic responses in ovalbumin-induced allergic rhinitis mouse and human mast cells. J. Ethnopharmacol. 2012, 142, 684–693. [Google Scholar]
- Fu, M.; Fu, S.; Ni, S.; Zou, L.; Liu, Y.; Hong, T. Anti-inflammatory effect of epigallocatechin gallate in a mouse model of ovalbumin-induced allergic rhinitis. Int. Immunopharmacol. 2017, 49, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.F.; Chan, Y.Y.; Shi, W.Y.; Jhong, M.T. Uncovering the Molecular Mechanism of Anti-Allergic Activity of Silkworm Pupa-Grown Cordyceps militaris Fruit Body. Am. J. Chin. Med. 2017, 45, 497–513. [Google Scholar] [CrossRef] [PubMed]
- Gelfand, E.W.; Joetham, A.; Wang, M.; Takeda, K.; Schedel, M. Spectrum of T-lymphocyte activities regulating allergic lung inflammation. Immunol. Rev. 2017, 278, 63–86. [Google Scholar] [CrossRef] [PubMed]
- Cardamone, C.; Parente, R.; Feo, G.D.; Triggiani, M. Mast cells as effector cells of innate immunity and regulators of adaptive immunity. Immunol. Lett. 2016, 178, 10–14. [Google Scholar] [CrossRef] [PubMed]
- Yi, Z.; Yi, Z.; Huang, K.; Cao, Y.; Xiao, C.; Li, Y.; Lu, Q.; Zhao, S.; Luo, W.; Liu, G. Propofol attenuates mast cell degranulation via inhibiting the miR-221/PI3K/Akt/Ca(2+) pathway. Exp. Ther. Med. 2018, 16, 1426–1432. [Google Scholar] [PubMed]
- Xu, H.; Gu, L.N.; Yang, Q.Y.; Zhao, D.Y.; Liu, F. MiR-221 promotes IgE-mediated activation of mast cells degranulation by PI3K/Akt/PLCγ/Ca2+ pathway. J. Bioenerg. Biomembr. 2016, 48, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Chai, F.N.; Ma, W.Y.; Zhang, J.; Xu, H.S.; Li, Y.F.; Zhou, Q.D.; Li, X.G.; Ye, X.L. Coptisine from Rhizoma coptidis exerts an anti-cancer effect on hepatocellular carcinoma by up-regulating miR-122. Biomed. Pharmacother. 2018, 103, 1002–1011. [Google Scholar] [CrossRef] [PubMed]
- Han, B.; Jiang, P.; Li, Z.; Yu, Y.; Huang, T.; Ye, X.; Li, X. Coptisine-induced apoptosis in human colon cancer cells (HCT-116) is mediated by PI3K/Akt and mitochondrial-associated apoptotic pathway. Phytomed. Int. J. Phytother. Phytopharmacol. 2018, 48, 152. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Yang, F.; Li, G.; Huang, J.; Liu, Y.; Zhang, Q.; Tang, Q.; Hu, C.; Zhang, R. Coptisine induces apoptosis in human hepatoma cells through activating 67-kDa laminin receptor/cGMP signaling. Front. Pharmacol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Zhang, H.; Hu, B.; Yang, L.; Wang, P.; Wang, F.; Meng, X. Coptisine from Coptis chinensis inhibits production of inflammatory mediators in lipopolysaccharide-stimulated RAW 264.7 murine macrophage cells. Eur. J. Pharmacol. 2016, 780, 106–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, K.; Hu, L.; Liao, W.; Yin, D.; Rui, F. Coptisine Prevented IL-β-Induced Expression of Inflammatory Mediators in Chondrocytes. Inflammation 2016, 39, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Nam, S.T.; Park, Y.H.; Kim, H.W.; Kim, H.S.; Lee, D.; Min, B.L.; Kim, Y.M.; Choi, W.S. Suppression of IgE-mediated mast cell activation and mouse anaphylaxis via inhibition of Syk activation by 8-formyl-7-hydroxy-4-methylcoumarin, 4μ8C. Toxicol. Appl. Pharmacol. 2017, 332, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.Z.; Ye, J.; Jin, G.Y.; Piao, H.M.; Cui, H.; Zheng, M.Y.; Yang, J.S.; Che, N.; Choi, Y.H.; Li, L.C. Asiaticoside Mitigates the Allergic Inflammation by Abrogating the Degranulation of Mast Cells. J. Agric. Food Chem. 2017, 65. [Google Scholar] [CrossRef] [PubMed]
- Puebla-Osorio, N.; Sarchio, S.N.E.; Ullrich, S.E.; Byrne, S.N. Detection of Infiltrating Mast Cells Using a Modified Toluidine Blue Staining. Methods Mol. Biol. 2017, 1627, 213. [Google Scholar] [PubMed]
- Ang, W.X.; Church, A.M.; Kulis, M.; Choi, H.W.; Burks, A.W.; Abraham, S.N. Mast cell desensitization inhibits calcium flux and aberrantly remodels actin. J. Clin. Invest. 2016, 126, 4103–4118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, N.R.; Moon, P.D.; Nam, S.Y.; Ryu, K.J.; Yoou, M.S.; Choi, J.H.; Hwang, S.Y.; Kim, H.M.; Jeong, H.J. Inhibitory effects of atractylone on mast cell-mediated allergic reactions. Chem. Biol. Interact. 2016, 258, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Zhao, D.Y.; Xu, H.; Zhou, H.; Yang, Q.Y.; Liu, F.; Zhou, G.P. Down-Regulation of MicroRNA-223 Promotes Degranulation via the PI3K/Akt Pathway by Targeting IGF-1R in Mast Cells. PLoS ONE 2015, 10, e0123575. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Piao, H.; Jiang, J.; Jin, G.; Zheng, M.; Yang, J.; Jin, X.; Sun, T.; Choi, Y.H.; Li, L. Polydatin inhibits mast cell-mediated allergic inflammation by targeting PI3K/Akt, MAPK, NF-κB and Nrf2/HO-1 pathways. Sci. Rep. 2017, 7, 11895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, H.A.; Han, N.R.; Kim, M.J.; Kim, H.M.; Jeong, H.J. Evaluation of the effect of kaempferol in a murine allergic rhinitis model. Eur. J. Pharmacol. 2013, 718, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Qu, S.; Min, L.; Ye, L.; Zhang, S.; Qin, T.; Hua, F. Effects of 1,25-dihydroxyvitamin D3 in an ovalbumin-induced allergic rhinitis model. Int. Immunopharmacol. 2017, 47, 182–189. [Google Scholar] [CrossRef] [PubMed]
- Falcone, F.H.; Wan, D.; Barwary, N.; Sagieisenberg, R. RBL cells as models for in vitro studies of mast cells and basophils. Immunol. Rev. 2018, 282, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Yoo, J.M.; Sok, D.E.; Kim, M.R. Effect of endocannabinoids on IgE-mediated allergic response in RBL-2H3 cells. Int. Immunopharmacol. 2013, 17, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Korinek, M.; Chen, K.M.; Jiang, Y.H.; El-Shazly, M.; Stocker, J.; Chou, C.K.; Hwang, T.L.; Wu, Y.C.; Chen, B.H.; Chang, F.R. Anti-allergic potential of Typhonium blumei: Inhibition of degranulation via suppression of PI3K/PLCγ2 phosphorylation and calcium influx. Phytomedicine 2016, 23, 1706–1715. [Google Scholar] [CrossRef] [PubMed]
- Hanieh, H.; Islam, V.I.H.; Saravanan, S.; Chellappandian, M.; Ragul, K.; Durga, A.; Venugopal, K.; Senthilkumar, V.; Senthilkumar, P.; Thirugnanasambantham, K. Pinocembrin, a novel histidine decarboxylase inhibitor with anti-allergic potential in in vitro. Eur. J. Pharmacol. 2017, 814, 178–186. [Google Scholar] [CrossRef] [PubMed]
- Gilfillan, A.M.; Tkaczyk, C. Integrated signalling pathways for mast-cell activation. Nat. Rev. Immunol. 2006, 6, 218–230. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Liu, Q.S.; Zhao, J.; Tang, H.P.; Sun, J. SIRT1 attenuates murine allergic rhinitis by downregulated HMGB 1/TLR4 pathway. Scand. J. Immunol. 2018, e12667. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Callaway, Z.; Kim, H.B.; Fujisawa, T.; Kim, C.K. The role of TNF-α in eosinophilic inflammation associated with RSV bronchiolitis. J. Allergy Clin. Immunol. 2008, 121, S122. [Google Scholar] [CrossRef]
- Yoo, J.M.; Kim, N.Y.; Seo, J.M.; Kim, S.J.; Lee, S.Y.; Kim, S.K.; Kim, H.D.; Lee, S.W.; Kim, M.R. Inhibitory effects of mulberry fruit extract in combination with naringinase on the allergic response in IgE-activated RBL-2H3 cells. Int. J. Mol. Med. 2014, 33, 469–477. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.W.; Israf Ali, D.A.; Harith, H.H.; Md Hashim, N.F.; Ng, C.H.; Shaari, K.; Tham, C.L. Anti-allergic activity of 2,4,6-trihydroxy-3-geranylacetophenone (tHGA) via attenuation of IgE-mediated mast cell activation and inhibition of passive systemic anaphylaxis. Toxicol. Appl. Pharmacol. 2017, 319, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Bement, W. Control of local actin assembly by membrane fusion-dependent compartment mixing. Nat. Cell Biol. 2007, 9, 149–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roth, K.; Chen, W.M.; Lin, T.J. Positive and negative regulatory mechanisms in high-affinity IgE receptor-mediated mast cell activation. Arch. Immunol. Ther. Exp. 2008, 56, 385–399. [Google Scholar] [CrossRef] [PubMed]
- Kitaura, J.; Asai, K.; Maeda-Yamamoto, M.; Kawakami, Y.; Kikkawa, U.; Kawakami, T. Akt-dependent cytokine production in mast cells. J. Exp. Med. 2000, 192, 729–740. [Google Scholar] [CrossRef] [PubMed]
- Bui, T.T.; Piao, C.H.; Song, C.H.; Chai, O.H. Skullcapflavone II attenuates ovalbumin-induced allergic rhinitis through the blocking of Th2 cytokine production and mast cell histamine release. Int. Immunopharmacol. 2017, 52, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.Y.; Zhou, Y.M.; Hu, M.; Li, J.Z.; Chen, C.J.; Wang, Y.J.; Shi, X.Y.; Wang, W.J.; Zhang, T.T. The Anti-Allergic Rhinitis Effect of Traditional Chinese Medicine of Shenqi by Regulating Mast Cell Degranulation and Th1/Th2 Cytokine Balance. Molecules 2017, 22, 504. [Google Scholar] [CrossRef] [PubMed]
- Sakat, M.S.; Kilic, K.; Kandemir, F.M.; Yildirim, S.; Sahin, A.; Kucukler, S.; Saglam, Y.S. The ameliorative effect of berberine and coenzyme Q10 in an ovalbumin-induced allergic rhinitis model. Eur. Arch. Oto-Rhino-Laryngol. 2018, 275, 2495–2505. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Feng, J.; Sun, J.; Wang, H.; Zhang, C.; Liu, H.; Zhang, H. H2-Eb1 expression is upregulated in the nasal mucosa of allergic rhinitis. Asian Pac. J. Allergy Immunol. 2014, 32, 308–315. [Google Scholar] [PubMed]
- Aswar, U.M.; Kandhare, A.D.; Vishwaraman, M.; Thakurdesai, P.A. Anti-allergic effect of intranasal administration of type-A procyanidin polyphenols based standardized extract of cinnamon bark in ovalbumin sensitized BALB/c mice. Phytotherapy Res. 2015, 29, 423–433. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |









© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fu, S.; Ni, S.; Wang, D.; Hong, T. Coptisine Suppresses Mast Cell Degranulation and Ovalbumin-Induced Allergic Rhinitis. Molecules 2018, 23, 3039. https://doi.org/10.3390/molecules23113039
Fu S, Ni S, Wang D, Hong T. Coptisine Suppresses Mast Cell Degranulation and Ovalbumin-Induced Allergic Rhinitis. Molecules. 2018; 23(11):3039. https://doi.org/10.3390/molecules23113039
Chicago/Turabian StyleFu, Shuilian, Saihong Ni, Danni Wang, and Tie Hong. 2018. "Coptisine Suppresses Mast Cell Degranulation and Ovalbumin-Induced Allergic Rhinitis" Molecules 23, no. 11: 3039. https://doi.org/10.3390/molecules23113039
APA StyleFu, S., Ni, S., Wang, D., & Hong, T. (2018). Coptisine Suppresses Mast Cell Degranulation and Ovalbumin-Induced Allergic Rhinitis. Molecules, 23(11), 3039. https://doi.org/10.3390/molecules23113039