Salvia elegans, Salvia greggii and Salvia officinalis Decoctions: Antioxidant Activities and Inhibition of Carbohydrate and Lipid Metabolic Enzymes

 ,
,  ,
, 
 and
and 
Abstract
:
1. Introduction
2. Results and Discussion
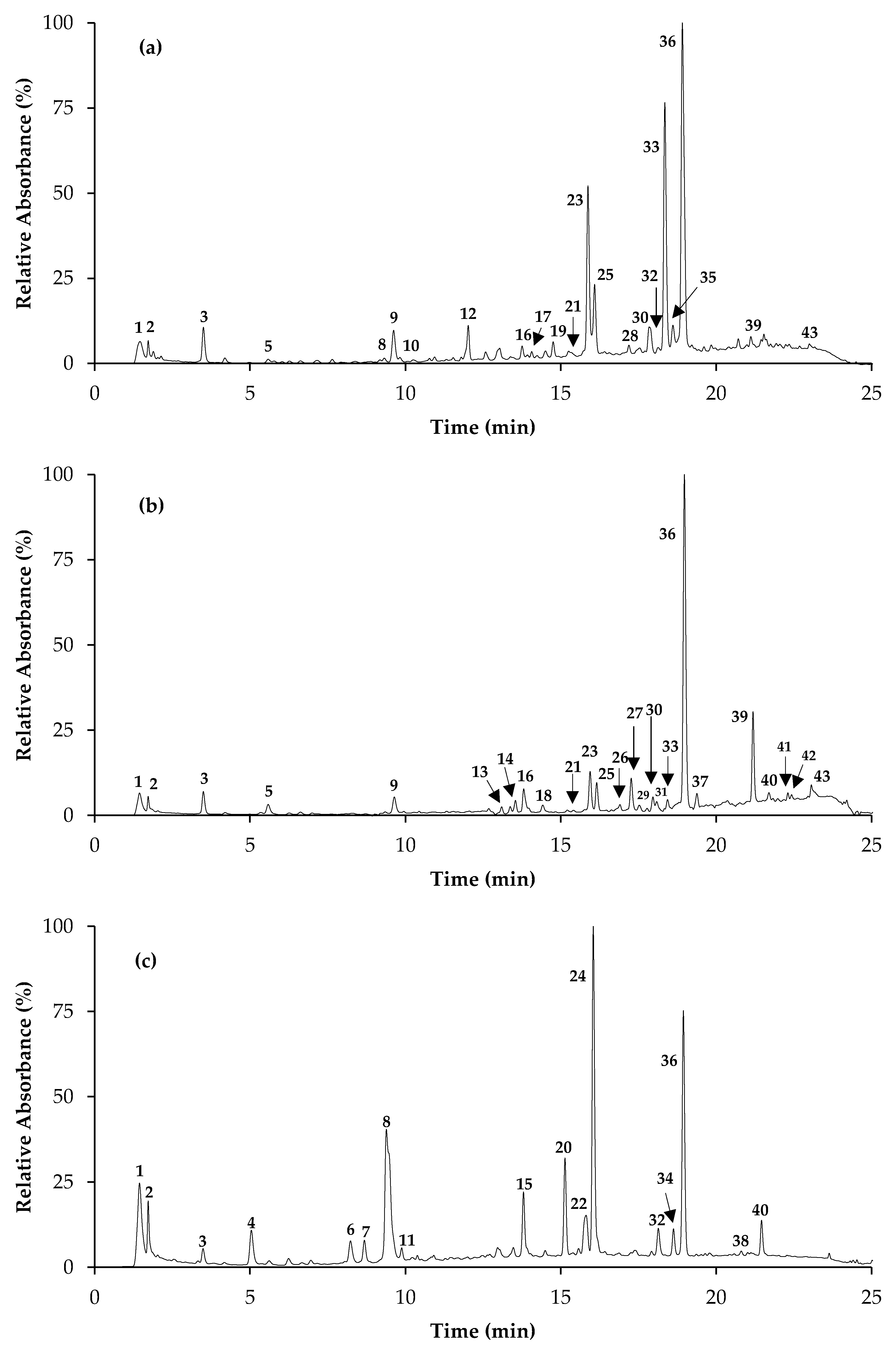
2.1. Phytochemical Composition
2.2. Biological Activities
2.2.1. Antioxidant Activity
2.2.2. Metabolic Enzyme Activity
3. Materials and Methods
3.1. Chemicals
3.2. Plant Sampling and Preparation of Extracts
3.3. Identification and Quantification of Phenolic Compounds
3.4. Antioxidant Activities
3.4.1. DPPH• Scavenging Assay
3.4.2. Ferric Reducing Antioxidant Power (FRAP) Assay
3.4.3. Oxygen Radical Absorbance Capacity (ORAC) Assay
3.4.4. NO• Scavenging Assay
3.4.5. Superoxide Anion (O2•–) Scavenging Assay
3.4.6. Inhibition of Xanthine Oxidase Activity
3.5. Inhibition of Enzymatic Activities
3.5.1. Inhibition of α-Glucosidase Activity
3.5.2. Inhibition of α-Amylase Activity
3.5.3. Inhibition of Pancreatic Lipase Activity
3.6. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Kintzios, S.E. Sage The Genus Salvia; Harwood Academic Publisher: Amsterdam, The Netherlands, 2000; ISBN 0203303660. [Google Scholar]
- Lu, Y.; Foo, L. Polyphenolics of Salvia—A review. Phytochemistry 2002, 59, 117–140. [Google Scholar] [CrossRef]
- Ghorbani, A.; Esmaeilizadeh, M. Pharmacological properties of Salvia officinalis and its components. J. Tradit. Complement. Med. 2017, 7, 433–440. [Google Scholar] [CrossRef] [PubMed]
- De Melo, G.A.N. Anti-inflammatory activity of Salvia officinalis L. J. Med. Plants Res. 2012, 6, 4934–4939. [Google Scholar] [CrossRef]
- Albano, S.M.; Miguel, M.G. Biological activities of extracts of plants grown in Portugal. Ind. Crops Prod. 2011, 33, 338–343. [Google Scholar] [CrossRef]
- Chohan, M.; Naughton, D.P.; Jones, L.; Opara, E.I. An investigation of the relationship between the anti-inflammatory activity, polyphenolic content, and antioxidant activities of cooked and in vitro digested culinary herbs. Oxidative Med. Cell. Longev. 2012, 2012, 627843. [Google Scholar] [CrossRef] [PubMed]
- Kozics, K.; Klusová, V.; Srančíková, A.; Mučaji, P.; Slameňová, D.; Hunáková, Ľ.; Kusznierewicz, B.; Horváthová, E. Effects of Salvia officinalis and Thymus vulgaris on oxidant-induced DNA damage and antioxidant status in HepG2 cells. Food Chem. 2013, 141, 2198–2206. [Google Scholar] [CrossRef] [PubMed]
- Ramos, A.A.; Azqueta, A.; Pereira-Wilson, C.; Collins, A.R. Polyphenolic compounds from Salvia species protect cellular DNA from oxidation and stimulate DNA repair in cultured human cells. J. Agric. Food Chem. 2010, 58, 7465–7471. [Google Scholar] [CrossRef]
- Walch, S.G.; Tinzoh, L.N.; Zimmermann, B.F.; Stühlinger, W.; Lachenmeier, D.W. Antioxidant capacity and polyphenolic composition as quality indicators for aqueous infusions of Salvia officinalis L. (sage tea). Front. Pharmacol. 2011, 2, 79. [Google Scholar] [CrossRef]
- Bettaieb, I.; Hamrouni-Sellami, I.; Bourgou, S.; Limam, F.; Marzouk, B. Drought effects on polyphenol composition and antioxidant activities in aerial parts of Salvia officinalis L. Acta Physiol. Plant. 2011, 33, 1103–1111. [Google Scholar] [CrossRef]
- Ozkan, G.; Kamiloglu, S.; Ozdal, T.; Boyacioglu, D.; Capanoglu, E. Potential use of Turkish medicinal plants in the treatment of various diseases. Molecules 2016, 21, 257. [Google Scholar] [CrossRef]
- Li, A.N.; Li, S.; Zhang, Y.J.; Xu, X.R.; Chen, Y.M.; Li, H.B. Resources and biological activities of natural polyphenols. Nutrients 2014, 6, 6020–6047. [Google Scholar] [CrossRef]
- Martins, N.; Barros, L.; Santos-Buelga, C.; Henriques, M.; Silva, S.; Ferreira, I.C.F.R. Evaluation of bioactive properties and phenolic compounds in different extracts prepared from Salvia officinalis L. Food Chem. 2014, 170, 378–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeshvaghani, Z.A.; Rahimmalek, M.; Talebi, M.; Goli, S.A.H. Comparison of total phenolic content and antioxidant activity in different Salvia species using three model systems. Ind. Crops Prod. 2015, 77, 409–414. [Google Scholar] [CrossRef]
- Farhat, M.B.; Landoulsi, A.; Chaouch-Hamada, R.; Sotomayor, J.A.; Jordán, M.J. Characterization and quantification of phenolic compounds and antioxidant properties of Salvia species growing in different habitats. Ind. Crops Prod. 2013, 49, 904–914. [Google Scholar] [CrossRef]
- Šulniūtė, V.; Ragažinskienė, O.; Venskutonis, P.R. Comprehensive Evaluation of Antioxidant Potential of 10 Salvia Species Using High Pressure Methods for the Isolation of Lipophilic and Hydrophilic Plant Fractions. Plant Foods Hum. Nutr. 2016, 71, 64–71. [Google Scholar] [CrossRef]
- Paun, G.; Neagu, E.; Moroeanu, V.; Ungureanu, O.; Cretu, R.; Ionescu, E.; Tebrencu, C.E. Phytochemical analysis and in vitro biological activity of Betonica officinalis and Salvia officinalis extracts. Rom. Biotechnol. Lett. 2017, 22, 12751–12761. [Google Scholar]
- Ullah, R.; Nadeem, M.; Khalique, A.; Imran, M.; Mehmood, S.; Javid, A.; Hussain, J. Nutritional and therapeutic perspectives of Chia (Salvia hispanica L.): A review. J. Food Sci. Technol. 2016, 53, 1750–1758, Epub 2015 Oct 1. [Google Scholar] [CrossRef]
- Min-hui, L.; Jian-min, C.; Yong, P.; Pei-gen, X. Distribution of Phenolic Acids in Chinese Salvia Plants. World Sci. Technol. 2008, 10, 46–52. [Google Scholar] [CrossRef]
- Zettel, V.; Hitzmann, B. Applications of chia (Salvia hispanica L.) in food products. Trends Food Sci. Technol. 2018, 80, 43–50. [Google Scholar] [CrossRef]
- Frett, J.F. Influence of nutrient salts, auxins and cytokinins on the in vitro growth of Salvia greggii. Plant Cell Tissue Organ Cult. 1987, 9, 89–93. [Google Scholar] [CrossRef]
- Starr, G. New World Salvias for Cultivation in Southern Arizona. Desert Plants 1985, 7, 184. [Google Scholar]
- Lim, T.K. Edible Medicinal and Non Medicinal Plants; Springer: Basel, Switzerland, 2014; ISBN 978-94-017-7276-1. [Google Scholar]
- Herrera-Ruiz, M.; García-Beltrán, Y.; Mora, S.; Díaz-Véliz, G.; Viana, G.S.B.; Tortoriello, J.; Ramírez, G. Antidepressant and anxiolytic effects of hydroalcoholic extract from Salvia elegans. J. Ethnopharmacol. 2006, 107, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Ferrer, E.; Badillo, F.H.; González-Cortazar, M.; Tortoriello, J.; Herrera-Ruiz, M. Antihypertensive activity of Salvia elegans Vahl. (Lamiaceae): ACE inhibition and angiotensin II antagonism. J. Ethnopharmacol. 2010, 130, 340–346. [Google Scholar] [CrossRef] [PubMed]
- Kawahara, N.; Tamura, T.; Inoue, M.; Hosoe, T.; Kawai, K.I.; Sekita, S.; Satake, M.; Goda, Y. Diterpenoid glucosides from Salvia greggii. Phytochemistry 2004, 65, 2577–2581. [Google Scholar] [CrossRef] [PubMed]
- Kawahara, N.; Inoue, M.; Kawai, K.I.; Sekita, S.; Satake, M.; Goda, Y. Diterpenoid from Salvia greggii. Phytochemistry 2003, 63, 859–862. [Google Scholar] [CrossRef]
- De Martino, L.; Roscigno, G.; Mancini, E.; De Falco, E.; De Feo, V. Chemical composition and antigerminative activity of the essential oils from five salvia species. Molecules 2010, 15, 735–746. [Google Scholar] [CrossRef] [PubMed]
- Pereira, O.R.; Afonso, A.F.; Cardoso, S.M. Advances in Analysis of Phenolic Compounds of Salvia, Thymus and Lavandula Plants. In Recent Progress in Medicinal Plants Series; Govil, J.N., Pathak, M., Eds.; Studium Press LLC: New Delhi, India, 2016; pp. 1–33. [Google Scholar]
- Cvetkovikj, I.; Stefkov, G.; Acevska, J.; Stanoeva, J.P.; Karapandzova, M.; Stefova, M.; Dimitrovska, A.; Kulevanova, S. Polyphenolic characterization and chromatographic methods for fast assessment of culinary Salvia species from South East Europe. J. Chromatogr. A 2013, 1282, 38–45. [Google Scholar] [CrossRef]
- Brahmi, N.; Scognamiglio, M.; Pacifico, S.; Mekhoukhe, A.; Madani, K.; Fiorentino, A.; Monaco, P. 1H NMR based metabolic profiling of eleven Algerian aromatic plants and evaluation of their antioxidant and cytotoxic properties. Food Res. Int. 2015, 76, 334–341. [Google Scholar] [CrossRef]
- Alimpić, A.; Knežević, A.; Milutinović, M.; Stević, T.; Šavikin, K.; Stajić, M.; Marković, S.; Marin, P.D.; Matevski, V.; Duletić-Laušević, S. Biological activities and chemical composition of Salvia amplexicaulis Lam. extracts. Ind. Crops Prod. 2017, 105, 1–9. [Google Scholar] [CrossRef]
- Alimpić, A.; Pljevljakušić, D.; Šavikin, K.; Knežević, A.; Ćurčić, M.; Veličković, D.; Stević, T.; Petrović, G.; Matevski, V.; Vukojević, J.; et al. Composition and biological effects of Salvia ringens (Lamiaceae) essential oil and extracts. Ind. Crops Prod. 2015, 76, 702–709. [Google Scholar] [CrossRef]
- Kostic, M.; Petrovic, M.B.; Jevtovic, T.; Jovic, M.; Petrovic, A.; Slavoljub, Ž. Anti-inflammatory effect of the Salvia sclarea L. ethanolic extract on lipopolysaccharide-induced periodontitis in rats. J. Ethnopharmacol. 2017, 199, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Bahadori, M.B.; Asghari, B.; Dinparast, L.; Zengin, G.; Sarikurkcu, C.; Abbas-Mohammadi, M.; Bahadori, S. Salvia nemorosa L.: A novel source of bioactive agents with functional connections. Food Sci. Thecnol. 2017, 75, 42–50. [Google Scholar] [CrossRef]
- Wang, B. Thousand Formulas and Thousand Herbs of Traditional Chinese Medicine; Heilongjiang Education Press: Harbin, China, 1993; Volume 2, ISBN 7531618990. [Google Scholar]
- Chen, L.; Kang, Y.H. Antioxidant and Enzyme Inhibitory Activities of Plebeian Herba (Salvia plebeia R. Br.) under Different Cultivation Conditions. J. Agric. Food Chem. 2014, 62, 2190–2197. [Google Scholar] [CrossRef] [PubMed]
- Mahesh, K.P.; Sasmal, D.; Papiya, M.M. In vitro antioxidant activity of methanolic extract of aerial parts of Salvia splendens (Scarlet sage). Pharmacogn. J. 2010, 2, 7–12. [Google Scholar]
- Liu, X.; Chen, R.; Shang, Y.; Jiao, B.; Huang, C. Superoxide radicals scavenging and xanthine oxidase inhibitory activity of magnesium lithospermate B from Salvia miltiorrhiza. J. Enzyme Inhib. Med. Chem. 2009, 24, 663–668. [Google Scholar] [CrossRef]
- Lin, D.; Xiao, M.; Zhao, J.; Li, Z.; Xing, B.; Li, X.; Kong, M.; Li, L.; Zhang, Q.; Liu, Y.; et al. An overview of plant phenolic compounds and their importance in human nutrition and management of type 2 diabetes. Molecules 2016, 21, 1374. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.K.; Kim, W.J.; Hyun, J.M.; Lee, J.S.; Kwon, J.G.; Seo, C.; Song, M.-J.; Choi, C.; Hong, S.S.; Park, K.; et al. Salvia plebeia Extract Inhibits Xanthine Oxidase Activity In Vitro and Reduces Serum Uric Acid in an Animal Model of Hyperuricemia. Planta Med. 2017, 83, 1335–1341. [Google Scholar] [CrossRef]
- Lin, C.-M.; Chen, C.-S.; Chen, C.-T.; Liang, Y.-C.; Lin, J.-K. Molecular modeling of flavonoids that inhibits XO. Biochem. Biophys. Res. Commun. 2002, 294, 167–172. [Google Scholar] [CrossRef]
- Jiao, R.H.; Ge, H.M.; Shi, D.H.; Tan, R.X. An apigenin-derived xanthine oxidase inhibitor from Palhinhaea cernua. J. Nat. Prod. 2006, 69, 1089–1091. [Google Scholar] [CrossRef]
- Vitaminol, J.N.S. Chemical Evidence for Potent Xanthine Oxidase Inhibitory Activity of Glechoma hederacea var. grandis leaves. J. Nutr. Sci. Vitaminol. 2013, 59, 570–575. [Google Scholar] [CrossRef]
- Moradabadi, L.; Kouhsari, S.M.; Sani, M.F. Hypoglycemic effects of three medicinal plants in experimental diabetes: Inhibition of rat intestinal α-glucosidase and enhanced pancreatic Insulin and cardiac Glut-4 mRNAs expression. Iran. J. Pharm. Res. 2013, 12, 385–397. [Google Scholar]
- Lunagariya, N.A.; Patel, N.K.; Jagtap, S.C.; Bhutani, K.K. Inhibitors of pancreatic lipase: State of the art and clinical perspectives. EXCLI J. 2014, 13, 897–921. [Google Scholar]
- Hamza, A.A.; Ksiksi, T.S.; Shamsi, O.A.A.; Balfaqh, S.A. α-Glucosidase Inhibitory Activity of Common Traditional Medicinal Plants Used for Diabetes Mellitus. J. Dev. Drugs 2015, 4, 2. [Google Scholar] [CrossRef]
- Loizzo, M.R.; Saab, A.M.; Tundis, R.; Menichini, F.; Bonesi, M.; Piccolo, V.; Statti, G.A.; de Cindio, B.; Houghton, P.J.; Menichini, F. In vitro inhibitory activities of plants used in Lebanon traditional medicine against angiotensin converting enzyme (ACE) and digestive enzymes related to diabetes. J. Ethnopharmacol. 2008, 119, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Asghari, B.; Salehi, P.; Sonboli, A.; Ebrahimi, S.N. Flavonoids from Salvia chloroleuca with alpha-Amylsae and alpha-Glucosidase Inhibitory Effect. Iran J. Pharm. Res. 2015, 14, 609–615. [Google Scholar] [PubMed]
- Kocak, M.S.; Sarikurkcu, C.; Cengiz, M.; Kocak, S.; Uren, M.C.; Tepe, B. Salvia cadmica: Phenolic composition and biological activity. Ind. Crops Prod. 2016, 85, 204–212. [Google Scholar] [CrossRef]
- Ma, H.Y.; Gao, H.Y.; Sun, L.; Huang, J.; Xu, X.M.; Wu, L.J. Constituents with α-glucosidase and advanced glycation end-product formation inhibitory activities from Salvia miltiorrhiza Bge. J. Nat. Med. 2011, 65, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, G.; Zamilpa, A.; Zavala, M.; Perez, J.; Morales, D.; Tortoriello, J. Chrysoeriol and other polyphenols from Tecoma stans with lipase inhibitory activity. J. Ethnopharmacol. 2016, 5, 1–8, Epub 2016 Mar 10. [Google Scholar] [CrossRef] [PubMed]
- Watcharachaisoponsiri, T.; Sornchan, P.; Charoenkiatkul, S.; Suttisansanee, U. The α-glucosidase and α-amylase inhibitory activity from different chili pepper extracts. Int. Food Res. J. 2016, 23, 1439–1445. [Google Scholar]
- Ninomiya, K.; Matsuda, H.; Shimoda, H.; Nishida, N.; Kasajima, N.; Yoshino, T.; Morikawa, T.; Yoshikawa, M. Carnosic acid, a new class of lipid absorption inhibitor from sage. Bioorg. Med. Chem. Lett. 2004, 14, 1943–1946. [Google Scholar] [CrossRef]
- Saad, B.; Zaid, H.; Shanak, S.; Kadan, S. Anti-Diabetes and Anti-Obesity Medicinal Plants and Phytochemicals; Springer: Basel, Switzerland, 2017; ISBN 978-3-319-54102-0. [Google Scholar]
- Arabiyat, S.; Al-Rabi’ee, A.; Zalloum, H.; Hudaib, M.; Mohammad, M.; Bustanji, Y. Antilipolytic and hypotriglyceridemic effects of dietary Salvia triloba Lf (Lamiaceae) in experimental rats. Trop. J. Pharm. Res. 2016, 15, 723–728. [Google Scholar] [CrossRef]
- Ferreira, F.M.; Dinis, L.T.; Azedo, P.; Galhano, C.I.C.; Simões, A.; Cardoso, S.M.; Domingues, M.R.M.; Pereira, O.R.; Palmeira, C.M.; Peixoto, F.P. Antioxidant capacity and toxicological evaluation of Pterospartum tridentatum flower extracts. CYTA J. Food 2012, 10, 92–102. [Google Scholar] [CrossRef]
- Afonso, A.F.; Pereira, O.R.; Neto, R.T.; Silva, A.M.S.; Cardoso, S.M. Health-promoting effects of Thymus herba-barona, Thymus pseudolanuginosus, and Thymus caespititius decoctions. Int. J. Mol. Sci. 2017, 18, 1879. [Google Scholar] [CrossRef] [PubMed]
- Catarino, M.D.; Silva, A.M.S.; Saraiva, S.C.; Sobral, A.J.F.N.; Cardoso, S.M. Characterization of phenolic constituents and evaluation of antioxidant properties of leaves and stems of Eriocephalus africanus. Arab. J. Chem. 2018, 11, 62–69. [Google Scholar] [CrossRef]
- Catarino, M.D.; Silva, A.M.S.; Cruz, M.T.; Cardoso, S.M. Antioxidant and anti-inflammatory activities of Geranium robertianum L. decoctions. Food Funct. 2017, 8, 3355–3365. [Google Scholar] [CrossRef] [PubMed]
- Yen, G.C.; Duh, P. Der. Scavenging Effect of Methanolic Extracts of Peanut Hulls on Free-Radical and Active-Oxygen Species. J. Agric. Food Chem. 1994, 42, 629–632. [Google Scholar] [CrossRef]
- Filha, Z.S.F.; Vitolo, I.F.; Fietto, L.G.; Lombardi, J.A.; Saúde-Guimarães, D.A. Xanthine oxidase inhibitory activity of Lychnophora species from Brazil (“Arnica”). J. Ethnopharmacol. 2006, 107, 79–82. [Google Scholar] [CrossRef]
- Neto, R.T.; Marçal, C.; Queirós, A.S.; Abreu, H.; Silva, A.M.S.; Cardoso, S.M. Screening of Ulva rigida, Gracilaria sp., Fucus vesiculosus and Saccharina latissima as Functional Ingredients. Int. J. Mol. Sci. 2018, 19, 2987. [Google Scholar] [CrossRef]
- Wickramaratne, M.N.; Punchihewa, J.C.; Wickramaratne, D.B.M. In-vitro alpha amylase inhibitory activity of the leaf extracts of Adenanthera pavonina. BMC Complement. Altern. Med. 2016, 16, 466. [Google Scholar] [CrossRef]
Sample Availability: Samples of extracts/compounds are available from the authors. |


{kind=link}
{kind=link}
| NP | RT (min) | λmax | [M − H]− | ESI-MS2 Main Fragments | Compound | S. off * | S. ele * | S. gre * |
|---|---|---|---|---|---|---|---|---|
| 1 | 1.5 | 275 | 149 | 103, 87, 131, 59 | 2,4-DimethylBA | 4.1 ± 0.2 | 5.1 ± 0.1 | 7.3 ± 0.2 |
| 2 | 1.7 | 205 | 191 | 111, 173 | Quinic acid | 0.6 ± 0.1 | 0.6 ± 0.1 | 0.5 ± 0.01 |
| 3 | 3.6 | 280 | 197 | 179, 73, 153 | Danshensu | D | D | D |
| 4 | 5.0 | 290, 324 | 353 | 191, 179, 135, 173 | cis 3-O-CQA | - | - | 3.1 ± 0.03 |
| 5 | 5.7 | 220, 278 | 137 | 109, 93, 119 | HydroxyBA | D | 2.0 ± 0.1 | - |
| 6 | 8.3 | 313 | 295 | 163 | p-Coum Ac Pent | - | - | 0.3 ± 0.02 |
| 7 | 8.8 | 290, 325 | 353 | 191, 179 | trans-5-O-CQA | - | - | 1.9 ± 0.1 |
| 8 | 9.4 | 313 | 265 | 177, 149, 119 | Coumaric Ac Der | - | - | 2.9 ± 0.05 |
| ND | 325 | 163, 119 | Caff Hex | D | - | - | ||
| 9 | 9.7 | 290, 323 | 179 | 135 | CaffAc | 1.8 ± 0.04 | 1.5 ± 0.02 | - |
| 10 | 9.8 | 314 | 325 | 265, 235, 163 | Coum Hex | D | - | - |
| 11 | 9.9 | 255, 265, 350 | 625 | 463, 301 | Querc diHex | - | - | D |
| 12 | 12.1 | 271, 336 | 593 | 473, 503, 353 | Api-6-C-Glc-7-O-Glc | 4.3 ± 0.1 | - | - |
| 13 | 13.1 | 291, 311 | 637 | 351, 285, 193 | Ferulic Ac Der | - | D | - |
| 14 | 13.5 | 274 | 571 | 527, 483, 439, 373 | YA E (isom1) | - | 1.9 ± 0.1 | - |
| 15 | 13.9 | 256, 267, 345 | 447 | 327, 357 | Lut-C-Hex | - | - | 4.6 ± 0.09 |
| 16 | 13.9 | 281, 345 | 477 | 301, 373, 343, 397 | Hydroxy-Lut-GlcA | D | 1.9 ± 0.2 | - |
| 17 | 14.1 | 276 | 571 | 527, 439, 553, 483 | YA E (isom2) | D | - | - |
| 18 | 14.4 | 269, 304 | 473 | 311, 293, 179, 135 | Cichoric acid | - | 1.6 ± 0.07 | - |
| 19 | 14.8 | 267, 345 | 621 | 351, 269 | Api-diGlcA | 4.6 ± 0.3 | - | - |
| 20 | 15.2 | 268, 336 | 431 | 311, 341, 269 | Api-C-Hex | - | - | 15.7 ± 0.3 |
| 21 | 15.4 | 274 | 555 | 313, 357 | SA K | 1.6 ± 0.2 | - | - |
| 571 | 527, 553, 509, 329 | YA E (isom3) | D | - | - | |||
| 22 | 15.8 | 255, 350 | 463 | 301 | Querc-O-Hex | - | - | 2.7 ± 0.2 |
| 23 | 15.9 | 280, 333 | 461 | 285 | Scut-O-GlcA | 13.4 ± 0.6 | 3.9 ± 0.1 | - |
| 24 | 16.0 | 255, 265, 348 | 447 | 285 | Lut-7-O-Glc | - | - | 26.1 ± 0.9 |
| 25 | 16.1 | 255, 266, 345 | 461 | 285 | Lut-7-O-GlcA (isom1) | 8.4 ± 0.3 | 5.1 ± 0.3 | - |
| 26 | 16.9 | 271, 306 | 521 | 359, 197, 179, 135 | Salviaflaside | - | D | - |
| 27 | 17.2 | 278 | 717 | 519, 475, 537, 339 | SA B (isom1) | - | 7.8 ± 0.4 | - |
| 28 | 17.3 | 279 | 571 | 527, 553, 329 | YA E (isom4) | 0.9 ± 0.1 | - | - |
| 29 | 17.7 | 279 | 717 | 537, 519, 339, 295 | SA B (isom2) | - | 1.7 ± 0.6 | - |
| 30 | 17.9 | 268, 334 | 577 | 269 | Api-rut | 4.5 ± 0.1 | D | - |
| 283 | 719 | 359, 539, 521, 341 | Sagerinic acid | 6.0 ± 0.3 | D | - | ||
| 31 | 18.1 | 271, 304 | 717 | 519, 607, 339, 537 | SA B (isom3) | - | 1.7 ± 0.1 | - |
| 32 | 18.1 | 269, 329 | 431 | 269 | Api-Hex | D | - | 3.4 ± 0.2 |
| 33 | 18.4 | 267, 337 | 445 | 269, 175 | Api-GlcA | 48.4 ± 1.3 | 3.2 ± 0.5 | - |
| 34 | 18.6 | 254, 266, 345 | 533 | 489, 447, 433 | Lut malonyl Hex | - | - | D |
| 35 | 18.6 | 270, 291, 326 | 717 | 555, 519, 475, 357 | SA B (isom4) | D | - | - |
| 36 | 19.0 | 218, 290, 328 | 359 | 161, 179, 197, 223 | RA | 28.3 ± 0.6 | 35.5 ± 0.8 | 10.9 ± 0.2 |
| 37 | 19.2 | 269, 307, 343 | 461 | 285 | Lut-O-GlcA (isom2) | - | 1.8 ± 0.1 | |
| 38 | 20.8 | 293, 328 | 373 | 343, 329, 311, 179 | Methyl rosmarinate | - | - | D |
| 39 | 21.2 | 290, 333 | 537 | 493, 359, 375 | CaffRA/SA I (isom1) | 1.2 ± 0.1 | 17.9 ± 0.1 | - |
| 40 | 21.4 | 293, 328 | 329 | 285, 314, 311, 161 | CaffAc derivative | - | - | 5.0 ± 0.03 |
| 239, 285, 330 | 537 | 456, 493, 375, 359 | CaffRA (isom2) | - | 1.3 ± 0.04 | - | ||
| 41 | 22.1 | 295, 325 | 713 | 493, 359, 375 | CaffAc der | - | D | - |
| 42 | 22.3 | 280 | 537 | 456, 493, 359, 161 | CaffRA (isom3) | - | 0.7 ± 0.04 | - |
| 43 | 23.0 | 289, 327 | 717 | 519, 357, 555 | SA B isomer | D | 2.4 ± 0.04 | - |
| Caffeic acid and derivatives | 39.8 ± 0.9 | 74.1 ± 0.5 | 20.8 ± 0.3 | |||||
| Coumaric acid derivatives | - | - | 3.2 ± 0.06 | |||||
| Flavones | 83.5 ± 2.3 | 15.9 ± 0.9 | 49.7 ± 1.3 | |||||
| Flavonols | - | - | 2.7 ± 0.2 | |||||
| S. officinalis | S. elegans | S. greggii | Standard | |
|---|---|---|---|---|
| DPPH• (EC50 μg/mL) (1) | 34.8 ± 3.3a | 10.7 ± 2.1b | 21.1 ± 2.5c | 6.69 ± 0.7b |
| Reducing Power (EC50 μg/mL) (2) | 40.0 ± 11.2a | 31.3 ± 5.0a,c | 77.9 ± 5.6b | 16.30 ± 1.5c |
| NO• (EC50 μg/mL) (1) | 118.2 ± 16.4a | 91.5 ± 14.5a | 167.8 ± 23.9b | 212.1 ± 9.7c |
| O2•– (EC50 μg/mL) (3) | 32.8 ± 0.6a | 30.6 ± 1.3a | 61.7 ± 3.4b | 7.8 ± 0.5c |
| ORAC (μM TE/mg ext) (4) | 404.4 ± 1.80a | 373.1 ± 28.1a | 335.6 ± 69.6a | - |
| Xanthine oxidase (EC50 μg/mL) (5) | 55.1 ± 10.6a | 71.8 ± 3.8b | 70.1 ± 4.0a,b | 0.09 ± 0.01c |
| DPPH | RP | ORAC | NO | O2 | XO | AG | L | |
|---|---|---|---|---|---|---|---|---|
| Flavones | − 0.971 | − 0.357 | 0.454 | − 0.498 | − 0.123 | 0.901 | − 0.551 | − 0.367 |
| Flavonols | − 0.239 | − 0.934 | − 0.891 | − 0.868 | − 0.992 | − 0.434 | − 0.835 | 0.930 |
| CafAcD | 0.801 | 0.948 | 0.400 | 0.986 | 0.844 | − 0.237 | 0.995 | − 0.485 |
| CouAcD | − 0.239 | − 0.934 | − 0.891 | − 0.868 | − 0.992 | − 0.434 | − 0.835 | 0.930 |
| DPPH | 0.570 | − 0.228 | 0.690 | 0.356 | − 0.771 | 0.734 | 0.134 | |
| RP | 0.670 | 0.988 | 0.971 | 0.084 | 0.976 | − 0.738 | ||
| ORAC | 0.547 | 0.829 | 0.796 | 0.493 | − 0.995 | |||
| NO | 0.922 | − 0.071 | 0.998 * | − 0.624 | ||||
| O2 | 0.321 | 0.996 | − 0.878 | |||||
| XO | − 0.134 | − 0.735 | ||||||
| AG | − 0.574 |
| S. officinalis | S. elegans | S. greggii | Standard | |
|---|---|---|---|---|
| α-Glucosidase (EC50 μg/mL) (1) | 71.2 ± 5.0a | 36.0 ± 2.7b | 345.3 ± 6.4c | 357.8 ± 12.3c |
| α-Amylase (2) | - | - | 6.5 ± 3.0 | 0.7 ± 0.2 |
| Pancreatic lipase (3) | 4.6 ± 3.6a | 8.2 ± 0.3a | 14.4 ± 7.4a | 1.8 ± 0.4 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pereira, O.R.; Catarino, M.D.; Afonso, A.F.; Silva, A.M.S.; Cardoso, S.M. Salvia elegans, Salvia greggii and Salvia officinalis Decoctions: Antioxidant Activities and Inhibition of Carbohydrate and Lipid Metabolic Enzymes. Molecules 2018, 23, 3169. https://doi.org/10.3390/molecules23123169
Pereira OR, Catarino MD, Afonso AF, Silva AMS, Cardoso SM. Salvia elegans, Salvia greggii and Salvia officinalis Decoctions: Antioxidant Activities and Inhibition of Carbohydrate and Lipid Metabolic Enzymes. Molecules. 2018; 23(12):3169. https://doi.org/10.3390/molecules23123169
Chicago/Turabian StylePereira, Olívia R., Marcelo D. Catarino, Andrea F. Afonso, Artur M. S. Silva, and Susana M. Cardoso. 2018. "Salvia elegans, Salvia greggii and Salvia officinalis Decoctions: Antioxidant Activities and Inhibition of Carbohydrate and Lipid Metabolic Enzymes" Molecules 23, no. 12: 3169. https://doi.org/10.3390/molecules23123169
APA StylePereira, O. R., Catarino, M. D., Afonso, A. F., Silva, A. M. S., & Cardoso, S. M. (2018). Salvia elegans, Salvia greggii and Salvia officinalis Decoctions: Antioxidant Activities and Inhibition of Carbohydrate and Lipid Metabolic Enzymes. Molecules, 23(12), 3169. https://doi.org/10.3390/molecules23123169