
Functional Characterization of the Saccharomyces cerevisiae Equilibrative Nucleoside Transporter 1 (ScENT1)

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
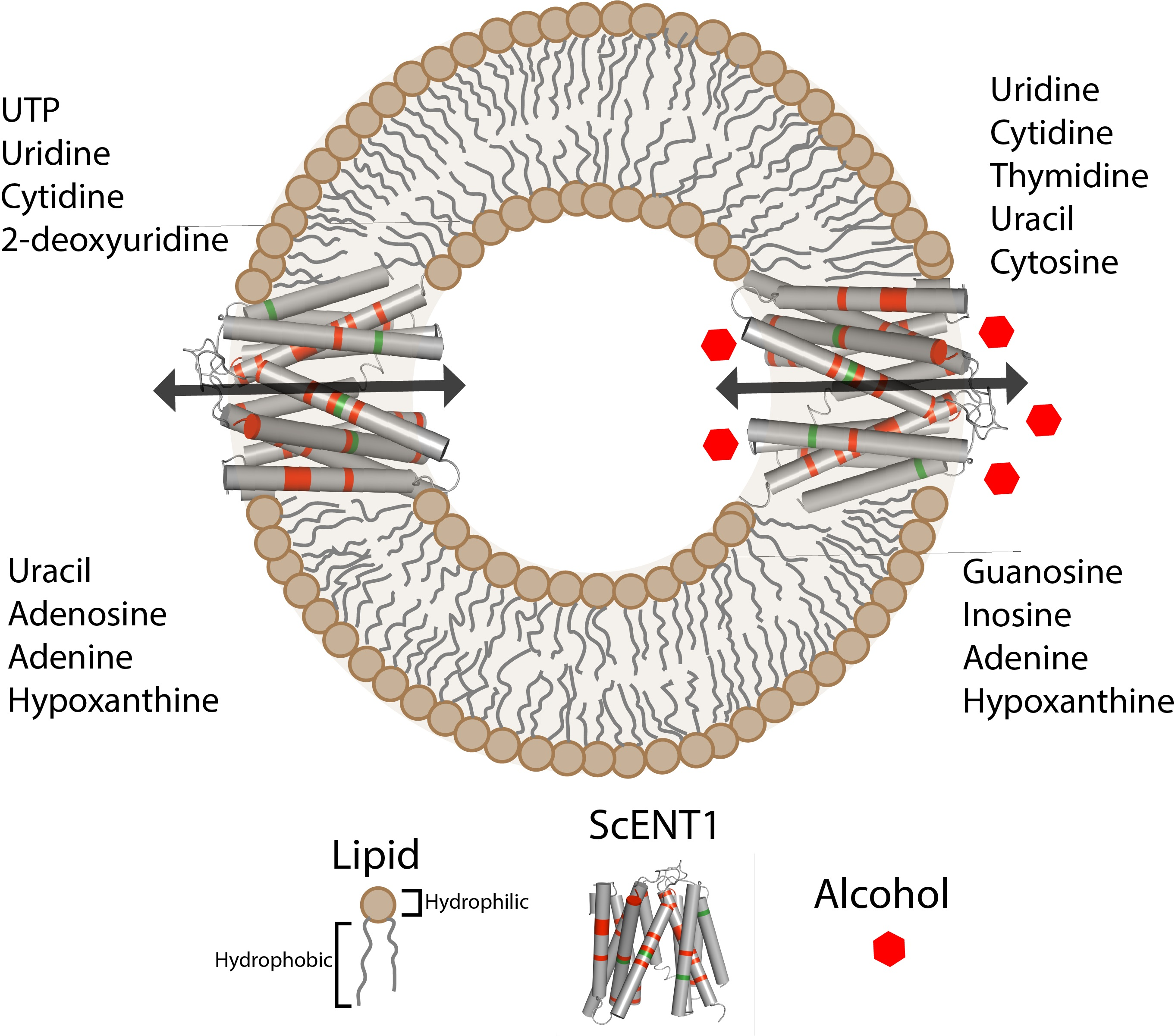
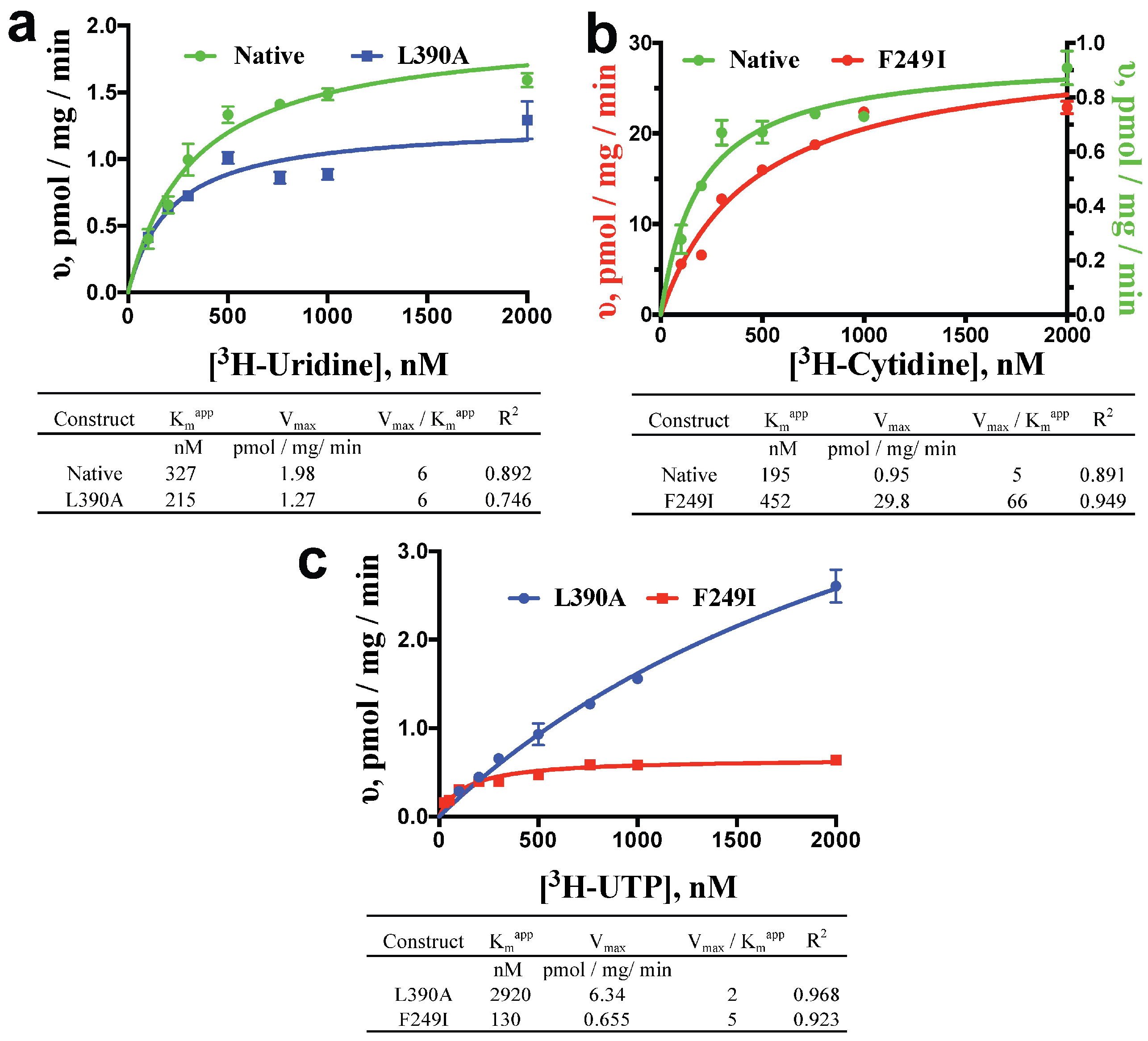
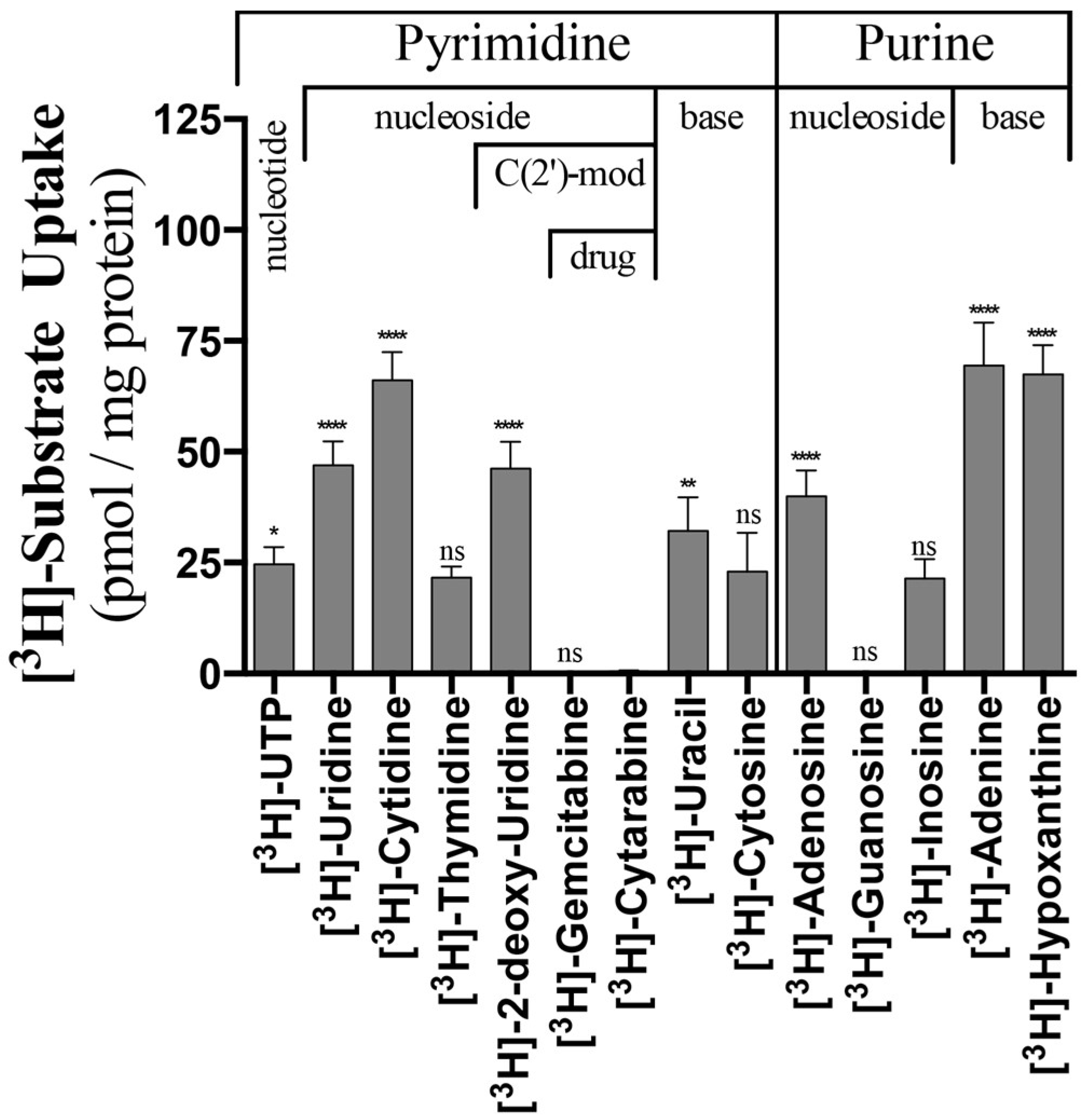
2.1. Nucleoside, Nucleobase and Nucleotide Uptake by the L390A and F249I Mutant Proteoliposomes
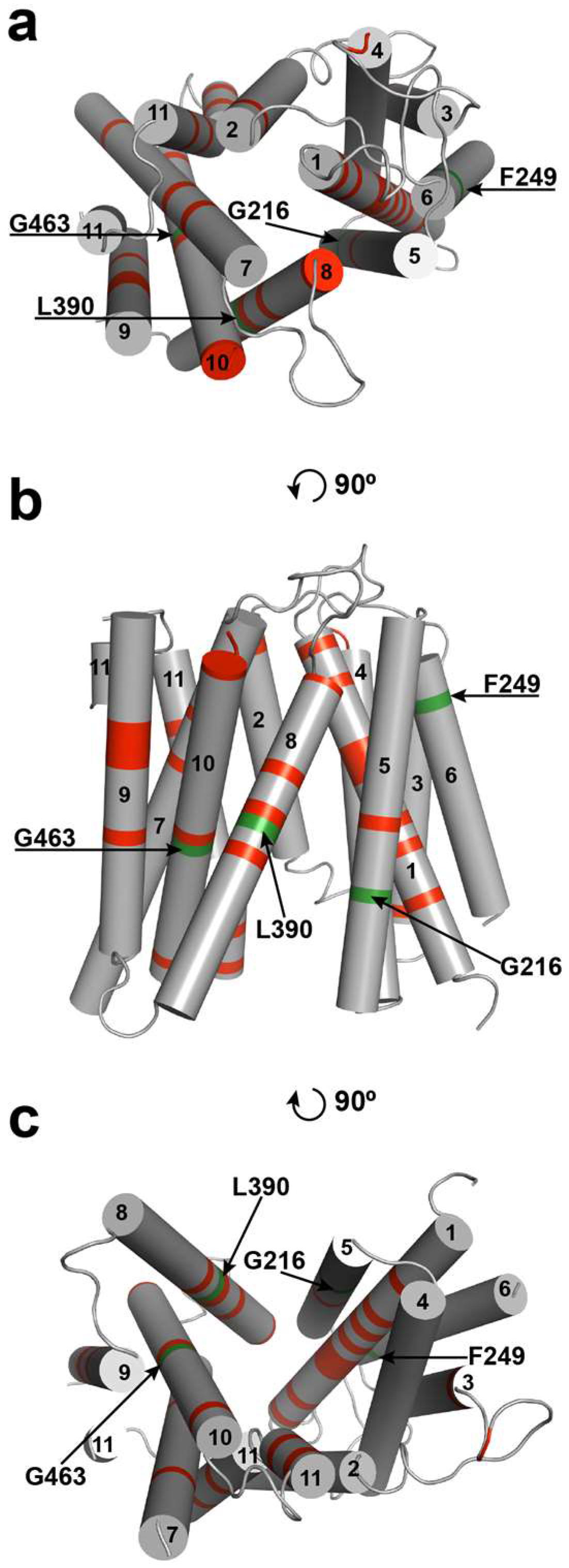
2.2. Ab Initio Modeling of FUN26
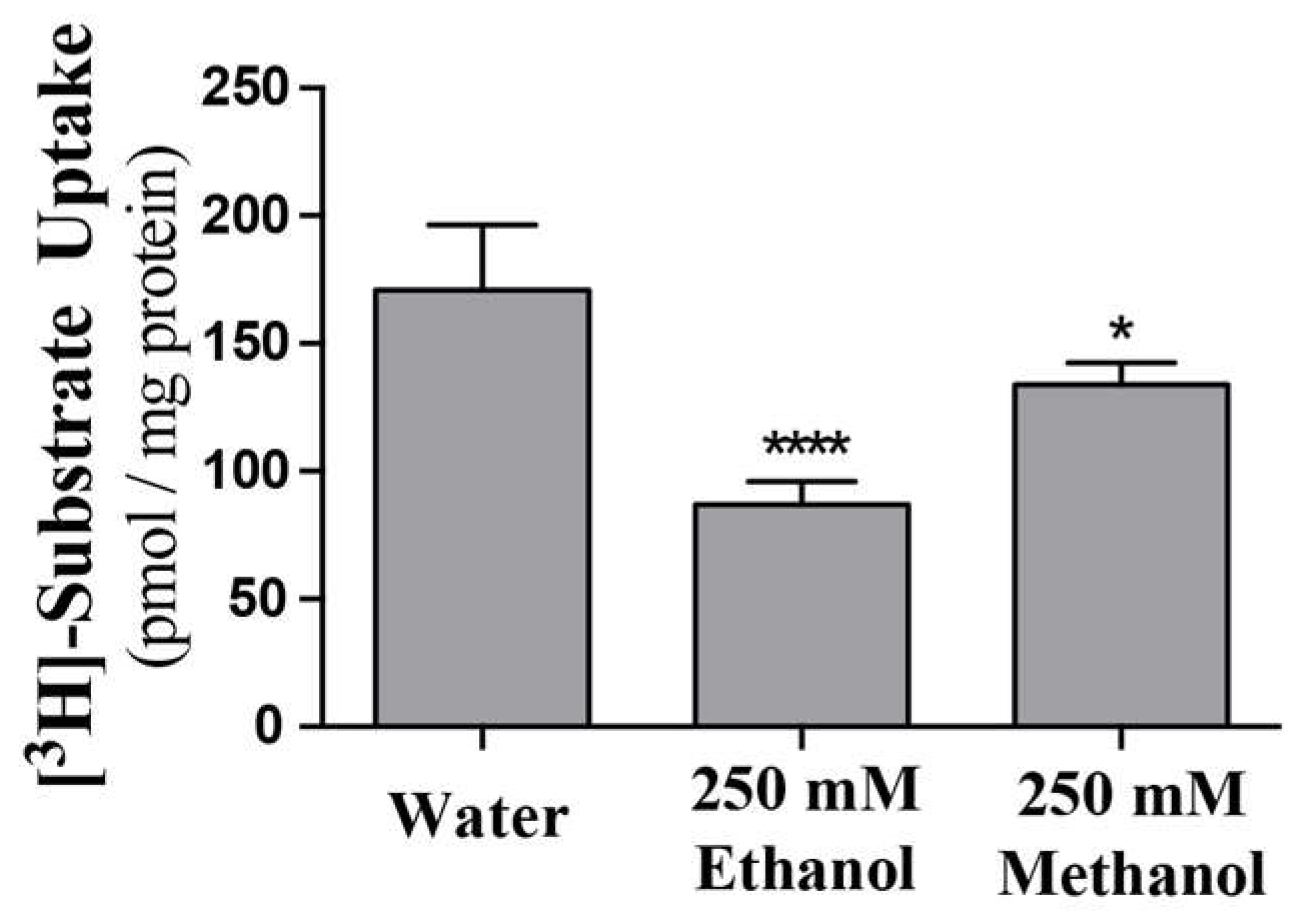
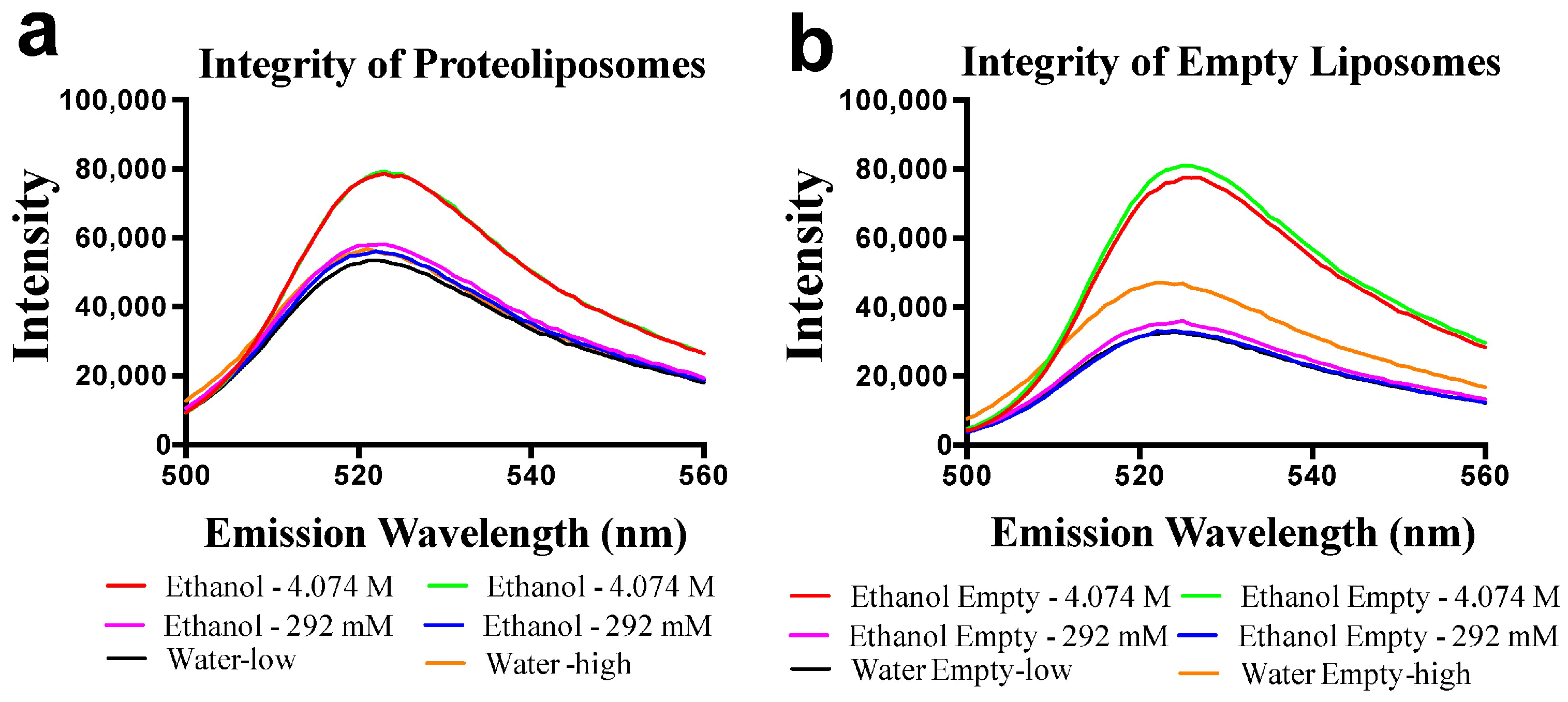
2.3. ScENT1-Mediated Transport is Attenuated by Alcohols
3. Discussion
4. Materials and Methods
4.1. Molecular Cloning and Protein Expression
4.2. Membrane Preparation, Solubilization and Purification
4.3. Gaussian Modeling
4.4. Ab Initio Model of the ScENT1 Inward-Open Configuration
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| ENT | Equilibrative nucleoside transporter |
| FUN26 | Function unknown now 26 |
| ScENT1 | Saccharomyces cerevisiae equilibrative nucleoside transporter 1 |
| IMP | Integral membrane protein |
| FDA | Food and Drug Administration |
| EMA | European Medicines Agency |
| hENT | Human equilibrative nucleoside transporter |
| TMD | Transmembrane domain |
| MFS | Major facilitator superfamily |
| PL | Proteoliposome |
| SC-His | Synthetic complete histidine dropout media |
| DDM | n-Dodecyl-β-d-maltoside |
| CF | 5(6)-Carboxyfluorescein |
| LacY | Lactose permease |
| GlpT | Glycerol 3-phosphate transporter |
| PHID syndrome | Pigmentary hypertrichosis and non-autoimmune insulin-dependent diabetes mellitus |
| Mhp1 | MAP-Homologous Protein 1 |
| LdNT1.1 | Leishmania donovani nucleoside transporter 1.1 |
| LUSH | General odorant binding protein lush |
| SEC | Size exclusion chromatography |
| NmR | N-ribosylnicotinamide |
| ns | Not significant |
References
- Jacobson, K.A. Introduction to adenosine receptors as therapeutic targets. Handb. Exp. Pharmacol. 2009, 193, 1–24. [Google Scholar]
- Li, R.W.; Yang, C.; Sit, A.S.; Lin, S.Y.; Ho, E.Y.; Leung, G.P. Physiological and pharmacological roles of vascular nucleoside transporters. J. Cardiovasc. Pharmacol. 2012, 59, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Young, J.D.; Yao, S.Y.; Baldwin, J.M.; Cass, C.E.; Baldwin, S.A. The human concentrative and equilibrative nucleoside transporter families, SLC28 and SLC29. Mol. Asp. Med. 2013, 34, 529–547. [Google Scholar] [CrossRef] [PubMed]
- Greenhalf, W.; Ghaneh, P.; Neoptolemos, J.P.; Palmer, D.H.; Cox, T.F.; Lamb, R.F.; Garner, E.; Campbell, F.; Mackey, J.R.; Costello, E.; et al. Pancreatic cancer hent1 expression and survival from gemcitabine in patients from the ESPAC-3 trial. J. Natl. Cancer Inst. 2014, 106, djt347. [Google Scholar] [CrossRef] [PubMed]
- Jordheim, L.P.; Durantel, D.; Zoulim, F.; Dumontet, C. Advances in the development of nucleoside and nucleotide analogues for cancer and viral diseases. Nat. Rev. Drug Discov. 2013, 12, 447–464. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Xu, P.; Chen, Y.; Liu, J.; Zhang, Y.; Lv, Y.; Luo, J.; Fang, M.; Zhang, J.; Wang, J.; et al. ENT1 inhibition attenuates epileptic seizure severity via regulation of glutamatergic neurotransmission. Neuromol. Med. 2015, 17, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Karpyak, V.M.; Biernacka, J.M.; Nam, H.W.; Lee, M.R.; Preuss, U.W.; Zill, P.; Yoon, G.; Colby, C.; Mrazek, D.A.; et al. Functional role of the polymorphic 647 T/C variant of ENT1 (SLC29A1) and its association with alcohol withdrawal seizures. PLoS ONE 2011, 6, e16331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nam, H.W.; Hinton, D.J.; Kang, N.Y.; Kim, T.; Lee, M.R.; Oliveros, A.; Adams, C.; Ruby, C.L.; Choi, D.S. Adenosine transporter ENT1 regulates the acquisition of goal-directed behavior and ethanol drinking through A2A receptor in the dorsomedial striatum. J. Neurosci. 2013, 33, 4329–4338. [Google Scholar] [CrossRef] [PubMed]
- Sundaram, M.; Yao, S.Y.; Ingram, J.C.; Berry, Z.A.; Abidi, F.; Cass, C.E.; Baldwin, S.A.; Young, J.D. Topology of a human equilibrative, nitrobenzylthioinosine (NBMPR)-sensitive nucleoside transporter (HENT1) implicated in the cellular uptake of adenosine and anti-cancer drugs. J. Biol. Chem. 2001, 276, 45270–45275. [Google Scholar] [CrossRef] [PubMed]
- Yan, N. Structural biology of the major facilitator superfamily transporters. Annu. Rev. Biophys. 2015, 44, 257–283. [Google Scholar] [CrossRef] [PubMed]
- Girke, C.; Arutyunova, E.; Syed, M.; Traub, M.; Mohlmann, T.; Lemieux, M.J. High yield expression and purification of equilibrative nucleoside transporter 7 (ENT7) from Arabidopsis thaliana. Biochim. Biophys. Acta 2015, 1850, 1921–1929. [Google Scholar] [CrossRef] [PubMed]
- Boswell-Casteel, R.C.; Johnson, J.M.; Duggan, K.D.; Roe-Zurz, Z.; Schmitz, H.; Burleson, C.; Hays, F.A. FUN26 (function unknown now 26) protein from saccharomyces cerevisiae is a broad selectivity, high affinity, nucleoside and nucleobase transporter. J. Biol. Chem. 2014, 289, 24440–24451. [Google Scholar] [CrossRef] [PubMed]
- Vickers, M.F.; Yao, S.Y.; Baldwin, S.A.; Young, J.D.; Cass, C.E. Nucleoside transporter proteins of saccharomyces cerevisiae. Demonstration of a transporter (FUI1) with high uridine selectivity in plasma membranes and a transporter (FUN26) with broad nucleoside selectivity in intracellular membranes. J. Biol. Chem. 2000, 275, 25931–25938. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.P.; Lin, S.J. Phosphate-responsive signaling pathway is a novel component of NAD+ metabolism in Saccharomyces cerevisiae. J. Biol. Chem. 2011, 286, 14271–14281. [Google Scholar] [CrossRef] [PubMed]
- Kang, N.; Jun, A.H.; Bhutia, Y.D.; Kannan, N.; Unadkat, J.D.; Govindarajan, R. Human equilibrative nucleoside transporter-3 (HENT3) spectrum disorder mutations impair nucleoside transport, protein localization, and stability. J. Biol. Chem. 2010, 285, 28343–28352. [Google Scholar] [CrossRef] [PubMed]
- SenGupta, D.J.; Lum, P.Y.; Lai, Y.; Shubochkina, E.; Bakken, A.H.; Schneider, G.; Unadkat, J.D. A single glycine mutation in the equilibrative nucleoside transporter gene, HENT1, alters nucleoside transport activity and sensitivity to nitrobenzylthioinosine. Biochemistry 2002, 41, 1512–1519. [Google Scholar] [CrossRef] [PubMed]
- Ramadan, A.; Naydenova, Z.; Stevanovic, K.; Rose, J.B.; Coe, I.R. The adenosine transporter, ENT1, in cardiomyocytes is sensitive to inhibition by ethanol in a kinase-dependent manner: implications for ethanol-dependent cardioprotection and nucleoside analog drug cytotoxicity. Purinergic Sign. 2014, 10, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Nam, H.W.; Lee, M.R.; Hinton, D.J.; Choi, S.; Kim, T.; Kawamura, T.; Janak, P.H.; Choi, D.S. Altered glutamatergic neurotransmission in the striatum regulates ethanol sensitivity and intake in mice lacking ENT1. Behav Brain Res. 2010, 208, 636–642. [Google Scholar] [CrossRef] [PubMed]
- Krauss, S.W.; Ghirnikar, R.B.; Diamond, I.; Gordon, A.S. Inhibition of adenosine uptake by ethanol is specific for one class of nucleoside transporters. Mol. Pharmacol. 1993, 44, 1021–1026. [Google Scholar] [PubMed]
- Jiang, X.; Loo, D.D.F.; Hirayama, B.A.; Wright, E.M. The importance of being aromatic: Pi interactions in sodium symporters. Biochemistry 2012, 51, 9480–9487. [Google Scholar] [CrossRef] [PubMed]
- Coe, I.R.; Dohrman, D.P.; Constantinescu, A.; Diamond, I.; Gordon, A.S. Activation of cyclic AMP-dependent protein kinase reverses tolerance of a nucleoside transporter to ethanol. J. Pharmacol. Exp. Ther. 1996, 276, 365–369. [Google Scholar] [PubMed]
- Lum, P.Y.; Ngo, L.Y.; Bakken, A.H.; Unadkat, J.D. Human intestinal es nucleoside transporter: Molecular characterization and nucleoside inhibitory profiles. Cancer Chemother. Pharmacol. 2000, 45, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Cass, C.E.; Paterson, A.R. Mediated transport of nucleosides by human erythrocytes. Specificity toward purine nucleosides as permeants. Biochim. Biophys. Acta 1973, 291, 734–746. [Google Scholar] [CrossRef]
- Abramson, J.; Smirnova, I.; Kasho, V.; Verner, G.; Kaback, H.R.; Iwata, S. Structure and mechanism of the lactose permease of escherichia coli. Science 2003, 301, 610–615. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Lemieux, M.J.; Song, J.; Auer, M.; Wang, D.N. Structure and mechanism of the glycerol-3-phosphate transporter from escherichia coli. Science 2003, 301, 616–620. [Google Scholar] [CrossRef] [PubMed]
- Yan, N. Structural advances for the major facilitator superfamily (MFS) transporters. Trends Biochem. Sci. 2013, 38, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Valdes, R.; Arastu-Kapur, S.; Landfear, S.M.; Shinde, U. An ab initio structural model of a nucleoside permease predicts functionally important residues. J. Biol. Chem. 2009, 284, 19067–19076. [Google Scholar] [CrossRef] [PubMed]
- Valdes, R.; Elferich, J.; Shinde, U.; Landfear, S.M. Identification of the intracellular gate for a member of the equilibrative nucleoside transporter (ENT) family. J. Biol. Chem. 2014, 289, 8799–8809. [Google Scholar] [CrossRef] [PubMed]
- Law, C.J.; Enkavi, G.; Wang, D.N.; Tajkhorshid, E. Structural basis of substrate selectivity in the glycerol-3-phosphate: Phosphate antiporter GlpT. Biophys. J. 2009, 97, 1346–1353. [Google Scholar] [CrossRef] [PubMed]
- Sanguinetti, M.; Amillis, S.; Pantano, S.; Scazzocchio, C.; Ramon, A. Modelling and mutational analysis of Aspergillus nidulans urea, a member of the subfamily of urea/H(+) transporters in fungi and plants. Open Biol. 2014, 4, 140070. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Jiang, X.; Kaback, H.R. Role of the irreplaceable residues in the LacY alternating access mechanism. Proc. Natl. Acad. Sci. USA 2012, 109, 12438–12442. [Google Scholar] [CrossRef] [PubMed]
- Boswell-Casteel, R.C.; Johnson, J.M.; Roe-Zurz, Z.; Duggan, K.D.; Schmitz, H.; Hays, F.A. Expression and purification of human and saccharomyces cerevisiae equilibrative nucleoside transporters. Protein Expr. Purif. 2018, 142, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Hammond, J.R. Enhancement of the functional stability of solubilized nucleoside transporters by substrates and inhibitors. Biochem. Pharmacol. 1997, 53, 623–629. [Google Scholar] [CrossRef]
- Hammond, J.R.; Zarenda, M. Effect of detergents on ligand binding and translocation activities of solubilized/reconstituted nucleoside transporters. Arch. Biochem. Biophys. 1996, 332, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Shimamura, T.; Weyand, S.; Beckstein, O.; Rutherford, N.G.; Hadden, J.M.; Sharples, D.; Sansom, M.S.; Iwata, S.; Henderson, P.J.; Cameron, A.D. Molecular basis of alternating access membrane transport by the sodium-hydantoin transporter Mhp1. Science 2010, 328, 470–473. [Google Scholar] [CrossRef] [PubMed]
- Yao, S.Y.; Ng, A.M.; Vickers, M.F.; Sundaram, M.; Cass, C.E.; Baldwin, S.A.; Young, J.D. Functional and molecular characterization of nucleobase transport by recombinant human and rat equilibrative nucleoside transporters 1 and 2. Chimeric constructs reveal a role for the ENT2 helix 5–6 region in nucleobase translocation. J. Biol. Chem. 2002, 277, 24938–24948. [Google Scholar] [CrossRef] [PubMed]
- Papageorgiou, I.; De Koning, H.P.; Soteriadou, K.; Diallinas, G. Kinetic and mutational analysis of the trypanosoma brucei NBT1 nucleobase transporter expressed in Saccharomyces cerevisiae reveals structural similarities between ent and mfs transporters. Int. J. Parasitol. 2008, 38, 641–653. [Google Scholar] [CrossRef] [PubMed]
- Swinnen, S.; Goovaerts, A.; Schaerlaekens, K.; Dumortier, F.; Verdyck, P.; Souvereyns, K.; Van Zeebroeck, G.; Foulquie-Moreno, M.R.; Thevelein, J.M. Auxotrophic mutations reduce tolerance of saccharomyces cerevisiae to very high levels of ethanol stress. Eukaryot. Cell 2015, 14, 884–897. [Google Scholar] [CrossRef] [PubMed]
- Cramer, A.C.; Vlassides, S.; Block, D.E. Kinetic model for nitrogen-limited wine fermentations. Biotechnol. Bioeng. 2002, 77, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Varela, C.; Pizarro, F.; Agosin, E. Biomass content governs fermentation rate in nitrogen-deficient wine musts. Appl. Environ. Microbiol. 2004, 70, 3392–3400. [Google Scholar] [CrossRef] [PubMed]
- Henderson, C.M.; Block, D.E. Examining the role of membrane lipid composition in determining the ethanol tolerance of Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2014, 80, 2966–2972. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Kawamata, T.; Horie, T.; Tsugawa, H.; Nakayama, Y.; Ohsumi, Y.; Fukusaki, E. Bulk RNA degradation by nitrogen starvation-induced autophagy in yeast. EMBO J. 2015, 34, 154–168. [Google Scholar] [CrossRef] [PubMed]
- Long, C.W.; Pardee, A.B. Cytidine triphosphate synthetase of Escherichia coli B. I. Purification and Kinetics. J. Biol. Chem. 1967, 242, 4715–4721. [Google Scholar] [PubMed]
- Ostrander, D.B.; O’Brien, D.J.; Gorman, J.A.; Carman, G.M. Effect of CTP synthetase regulation by ctp on phospholipid synthesis in Saccharomyces cerevisiae. J. Biol. Chem. 1998, 273, 18992–19001. [Google Scholar] [CrossRef] [PubMed]
- Ader, L.; Jones, D.N.; Lin, H. Alcohol binding to the odorant binding protein LUSH: Multiple factors affecting binding affinities. Biochemistry 2010, 49, 6136–6142. [Google Scholar] [CrossRef] [PubMed]
- Kumar, H.; Finer-Moore, J.S.; Kaback, H.R.; Stroud, R.M. Structure of LacY with an alpha-substituted galactoside: Connecting the binding site to the protonation site. Proc. Natl. Acad. Sci. USA 2015, 112, 9004–9009. [Google Scholar] [CrossRef] [PubMed]
- Gerlsma, S.Y.; Stuur, E.R. The effect of polyhydric and monohydric alcohols on the heat-induced reversible denaturation of lysozyme and ribonuclease. Int. J. Pept. Protein Res. 1972, 4, 377–383. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Bolen, D.W. The peptide backbone plays a dominant role in protein stabilization by naturally occurring osmolytes. Biochemistry 1995, 34, 12884–12891. [Google Scholar] [CrossRef] [PubMed]
- Thomas, P.D.; Dill, K.A. Local and nonlocal interactions in globular proteins and mechanisms of alcohol denaturation. Protein Sci. 1993, 2, 2050–2065. [Google Scholar] [CrossRef] [PubMed]
- Ma, M.; Liu, Z.L. Mechanisms of ethanol tolerance in Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2010, 87, 829–845. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.H.; Wang, M.H.; Tan, T.; Li, J.R.; Yang, H.; Leach, L.; Zhang, R.M.; Luo, Z.W. Genetic dissection of ethanol tolerance in the budding yeast Saccharomyces cerevisiae. Genetics 2007, 175, 1479–1487. [Google Scholar] [CrossRef] [PubMed]
- Stanley, D.; Bandara, A.; Fraser, S.; Chambers, P.J.; Stanley, G.A. The ethanol stress response and ethanol tolerance of Saccharomyces cerevisiae. J. Appl. Microbiol. 2010, 109, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Leao, C.; van Uden, N. Effects of ethanol and other alkanols on the general amino acid permease of Saccharomyces cerevisiae. Biotechnol. Bioeng. 1984, 26, 403–405. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, K.; Tanaka, T.; Furusawa, C.; Nagahisa, K.; Hirasawa, T.; Shimizu, H. Comprehensive phenotypic analysis for identification of genes affecting growth under ethanol stress in Saccharomyces cerevisiae. FEMS Yeast Res. 2009, 9, 32–44. [Google Scholar] [CrossRef] [PubMed]
- VanderSluis, B.; Hess, D.C.; Pesyna, C.; Krumholz, E.W.; Syed, T.; Szappanos, B.; Nislow, C.; Papp, B.; Troyanskaya, O.G.; Myers, C.L.; et al. Broad metabolic sensitivity profiling of a prototrophic yeast deletion collection. Genome Biol. 2014, 15, R64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belenky, P.A.; Moga, T.G.; Brenner, C. Saccharomyces cerevisiae YOR071C encodes the high affinity nicotinamide riboside transporter Nrt1. J. Biol. Chem. 2008, 283, 8075–8079. [Google Scholar] [CrossRef] [PubMed]
- Allen-Gipson, D.S.; Jarrell, J.C.; Bailey, K.L.; Robinson, J.E.; Kharbanda, K.K.; Sisson, J.H.; Wyatt, T.A. Ethanol blocks adenosine uptake via inhibiting the nucleoside transport system in bronchial epithelial cells. Alcohol. Clin. Exp. Res. 2009, 33, 791–798. [Google Scholar] [CrossRef] [PubMed]
- Choi, D.S.; Cascini, M.G.; Mailliard, W.; Young, H.; Paredes, P.; McMahon, T.; Diamond, I.; Bonci, A.; Messing, R.O. The type 1 equilibrative nucleoside transporter regulates ethanol intoxication and preference. Nat. Neurosci. 2004, 7, 855–861. [Google Scholar] [CrossRef] [PubMed]
- Osato, D.H.; Huang, C.C.; Kawamoto, M.; Johns, S.J.; Stryke, D.; Wang, J.; Ferrin, T.E.; Herskowitz, I.; Giacomini, K.M. Functional characterization in yeast of genetic variants in the human equilibrative nucleoside transporter, ENT1. Pharmacogenetics 2003, 13, 297–301. [Google Scholar] [CrossRef] [PubMed]
- Marechal, R.; Bachet, J.B.; Mackey, J.R.; Dalban, C.; Demetter, P.; Graham, K.; Couvelard, A.; Svrcek, M.; Bardier-Dupas, A.; Hammel, P.; et al. Levels of gemcitabine transport and metabolism proteins predict survival times of patients treated with gemcitabine for pancreatic adenocarcinoma. Gastroenterology 2012, 143, 664–674. [Google Scholar] [CrossRef] [PubMed]
- Murata, Y.; Hamada, T.; Kishiwada, M.; Ohsawa, I.; Mizuno, S.; Usui, M.; Sakurai, H.; Tabata, M.; Ii, N.; Inoue, H.; et al. Human equilibrative nucleoside transporter 1 expression is a strong independent prognostic factor in UICC T3-T4 pancreatic cancer patients treated with preoperative gemcitabine-based chemoradiotherapy. J. Hepatobiliary Pancreat. Sci. 2012, 19, 413–425. [Google Scholar] [CrossRef] [PubMed]
- Morinaga, S.; Nakamura, Y.; Watanabe, T.; Mikayama, H.; Tamagawa, H.; Yamamoto, N.; Shiozawa, M.; Akaike, M.; Ohkawa, S.; Kameda, Y.; et al. Immunohistochemical analysis of human equilibrative nucleoside transporter-1 (hENT1) predicts survival in resected pancreatic cancer patients treated with adjuvant gemcitabine monotherapy. Ann. Surg. Oncol. 2012, 19 (Suppl 3), S558–S564. [Google Scholar] [CrossRef] [PubMed]
- Fujita, H.; Ohuchida, K.; Mizumoto, K.; Itaba, S.; Ito, T.; Nakata, K.; Yu, J.; Kayashima, T.; Souzaki, R.; Tajiri, T.; et al. Gene expression levels as predictive markers of outcome in pancreatic cancer after gemcitabine-based adjuvant chemotherapy. Neoplasia 2010, 12, 807–817. [Google Scholar] [CrossRef] [PubMed]
- Marechal, R.; Mackey, J.R.; Lai, R.; Demetter, P.; Peeters, M.; Polus, M.; Cass, C.E.; Young, J.; Salmon, I.; Deviere, J.; et al. Human equilibrative nucleoside transporter 1 and human concentrative nucleoside transporter 3 predict survival after adjuvant gemcitabine therapy in resected pancreatic adenocarcinoma. Clin. Cancer Res. 2009, 15, 2913–2919. [Google Scholar] [CrossRef] [PubMed]
- Farrell, J.J.; Elsaleh, H.; Garcia, M.; Lai, R.; Ammar, A.; Regine, W.F.; Abrams, R.; Benson, A.B.; Macdonald, J.; Cass, C.E.; et al. Human equilibrative nucleoside transporter 1 levels predict response to gemcitabine in patients with pancreatic cancer. Gastroenterology 2009, 136, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Giovannetti, E.; Del Tacca, M.; Mey, V.; Funel, N.; Nannizzi, S.; Ricci, S.; Orlandini, C.; Boggi, U.; Campani, D.; Del Chiaro, M.; et al. Transcription analysis of human equilibrative nucleoside transporter-1 predicts survival in pancreas cancer patients treated with gemcitabine. Cancer Res. 2006, 66, 3928–3935. [Google Scholar] [CrossRef] [PubMed]
- Spratlin, J.; Sangha, R.; Glubrecht, D.; Dabbagh, L.; Young, J.D.; Dumontet, C.; Cass, C.; Lai, R.; Mackey, J.R. The absence of human equilibrative nucleoside transporter 1 is associated with reduced survival in patients with gemcitabine-treated pancreas adenocarcinoma. Clin. Cancer Res. 2004, 10, 6956–6961. [Google Scholar] [CrossRef] [PubMed]
- Indiveri, C. Studying amino acid transport using liposomes. Methods Mol. Biol. 2010, 606, 55–68. [Google Scholar] [PubMed]
- Yarov-Yarovoy, V.; Schonbrun, J.; Baker, D. Multipass membrane protein structure prediction using Rosetta. Proteins 2006, 62, 1010–1025. [Google Scholar] [CrossRef] [PubMed]
- Dang, S.; Sun, L.; Huang, Y.; Lu, F.; Liu, Y.; Gong, H.; Wang, J.; Yan, N. Structure of a fucose transporter in an outward-open conformation. Nature 2010, 467, 734–738. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.T. Do transmembrane protein superfolds exist? FEBS Lett. 1998, 423, 281–285. [Google Scholar] [CrossRef]
- Sali, A.; Blundell, T.L. Comparative protein modelling by satisfaction of spatial restraints. J. Mol. Biol. 1993, 234, 779–815. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Zhang, Y. LOMETS: A local meta-threading-server for protein structure prediction. Nucleic Acids Res. 2007, 35, 3375–3382. [Google Scholar] [CrossRef] [PubMed]
- Mackey, J.R.; Mani, R.S.; Selner, M.; Mowles, D.; Young, J.D.; Belt, J.A.; Crawford, C.R.; Cass, C.E. Functional nucleoside transporters are required for gemcitabine influx and manifestation of toxicity in cancer cell lines. Cancer Res. 1998, 58, 4349–4357. [Google Scholar] [PubMed]
- Achiwa, H.; Oguri, T.; Sato, S.; Maeda, H.; Niimi, T.; Ueda, R. Determinants of sensitivity and resistance to gemcitabine: The roles of human equilibrative nucleoside transporter 1 and deoxycytidine kinase in non-small cell lung cancer. Cancer Sci. 2004, 95, 753–757. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.P.; Wen, J.; Bang, S.; Park, S.; Song, S.Y. CD44-positive cells are responsible for gemcitabine resistance in pancreatic cancer cells. Int. J. Cancer 2009, 125, 2323–2331. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Radioactive substrates are available from sources indicated. Purified protein is not available from the authors. |






© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boswell-Casteel, R.C.; Johnson, J.M.; Hays, F.A. Functional Characterization of the Saccharomyces cerevisiae Equilibrative Nucleoside Transporter 1 (ScENT1). Molecules 2018, 23, 732. https://doi.org/10.3390/molecules23040732
Boswell-Casteel RC, Johnson JM, Hays FA. Functional Characterization of the Saccharomyces cerevisiae Equilibrative Nucleoside Transporter 1 (ScENT1). Molecules. 2018; 23(4):732. https://doi.org/10.3390/molecules23040732
Chicago/Turabian StyleBoswell-Casteel, Rebba C., Jennifer M. Johnson, and Franklin A. Hays. 2018. "Functional Characterization of the Saccharomyces cerevisiae Equilibrative Nucleoside Transporter 1 (ScENT1)" Molecules 23, no. 4: 732. https://doi.org/10.3390/molecules23040732
APA StyleBoswell-Casteel, R. C., Johnson, J. M., & Hays, F. A. (2018). Functional Characterization of the Saccharomyces cerevisiae Equilibrative Nucleoside Transporter 1 (ScENT1). Molecules, 23(4), 732. https://doi.org/10.3390/molecules23040732