Red and Blue Light Promote the Accumulation of Artemisinin in Artemisia annua L.


Abstract
:1. Introduction
2. Results and Discussion
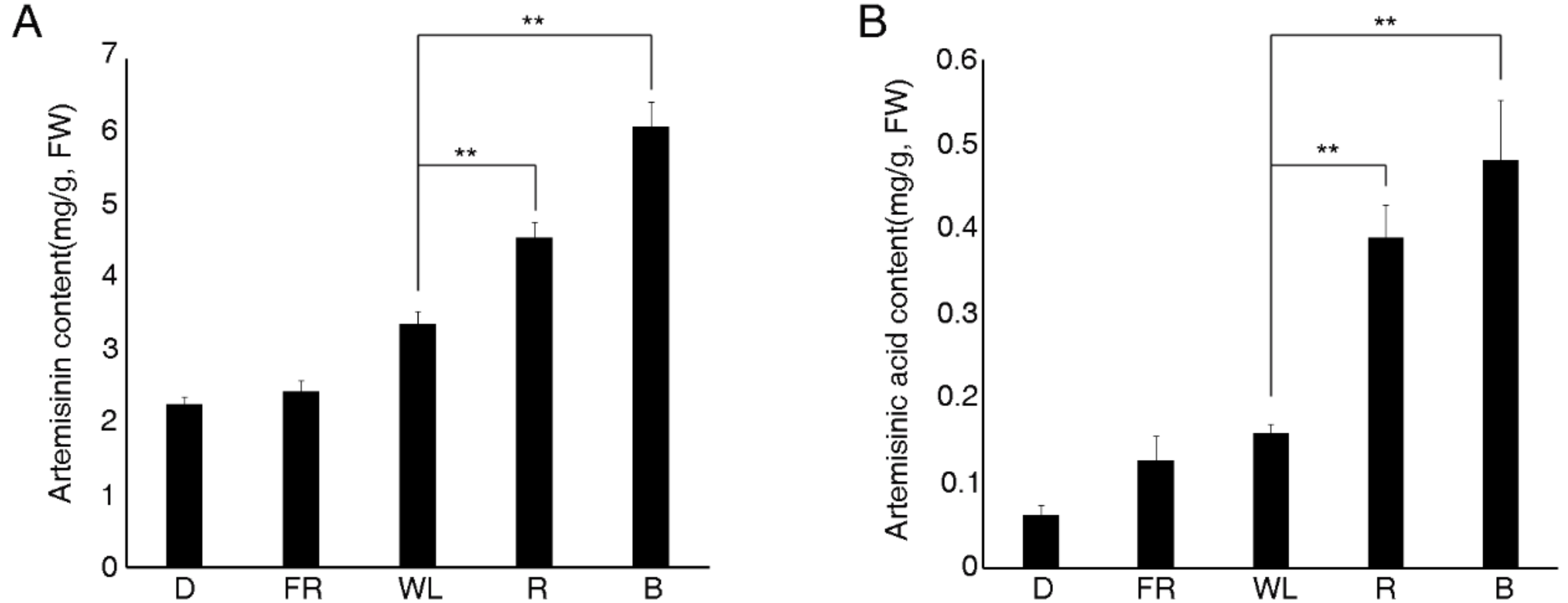
2.1. Analysis of Artemisinin Content under Various Treatments
2.2. Transcriptome Sequencing and Assembly
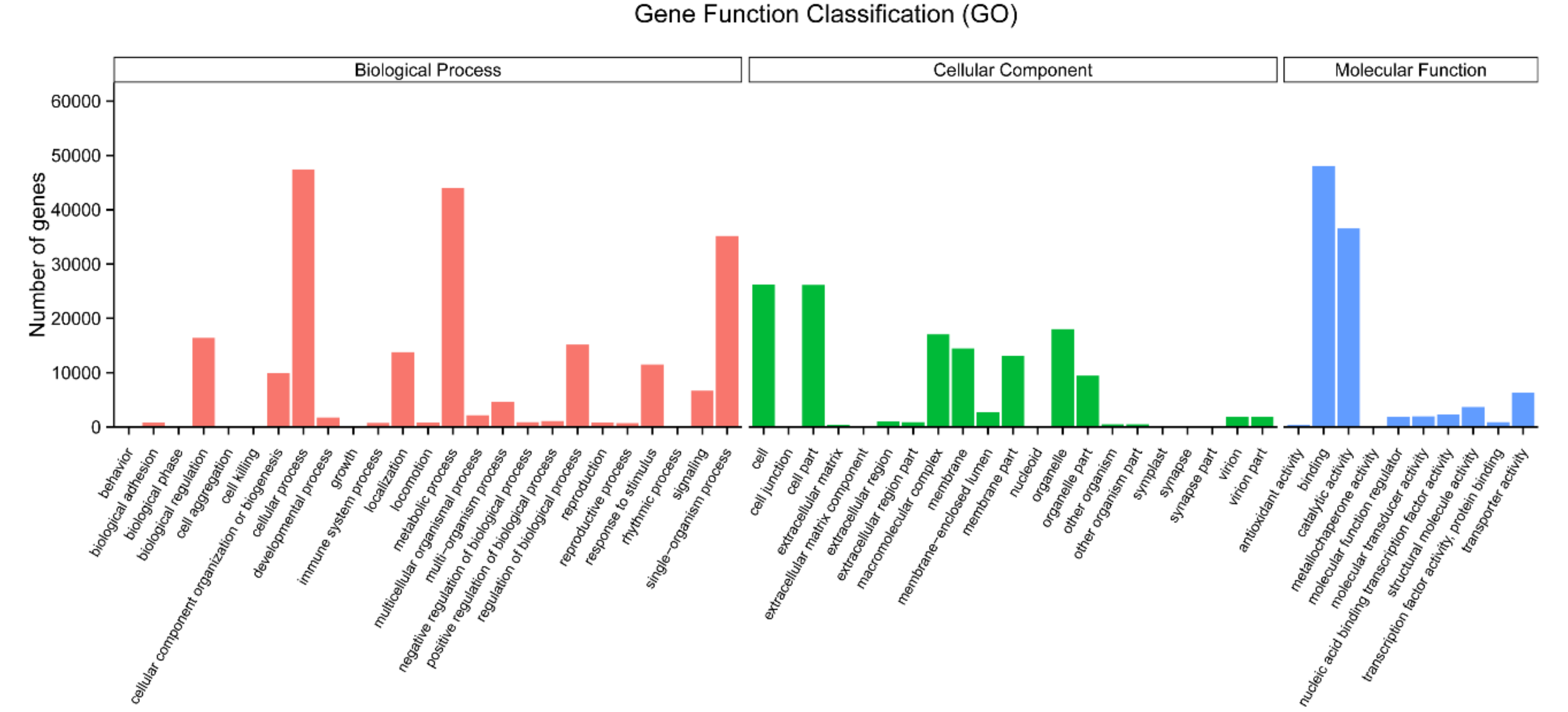
2.3. Functional Annotation and Classification
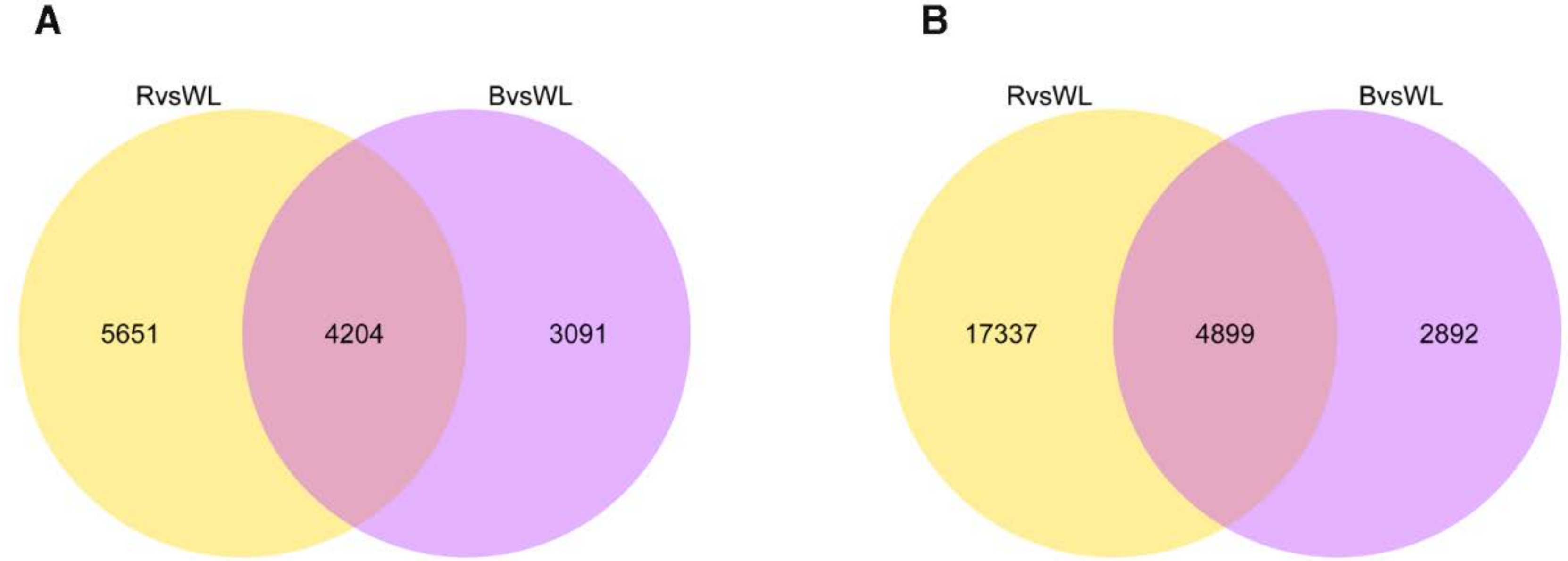
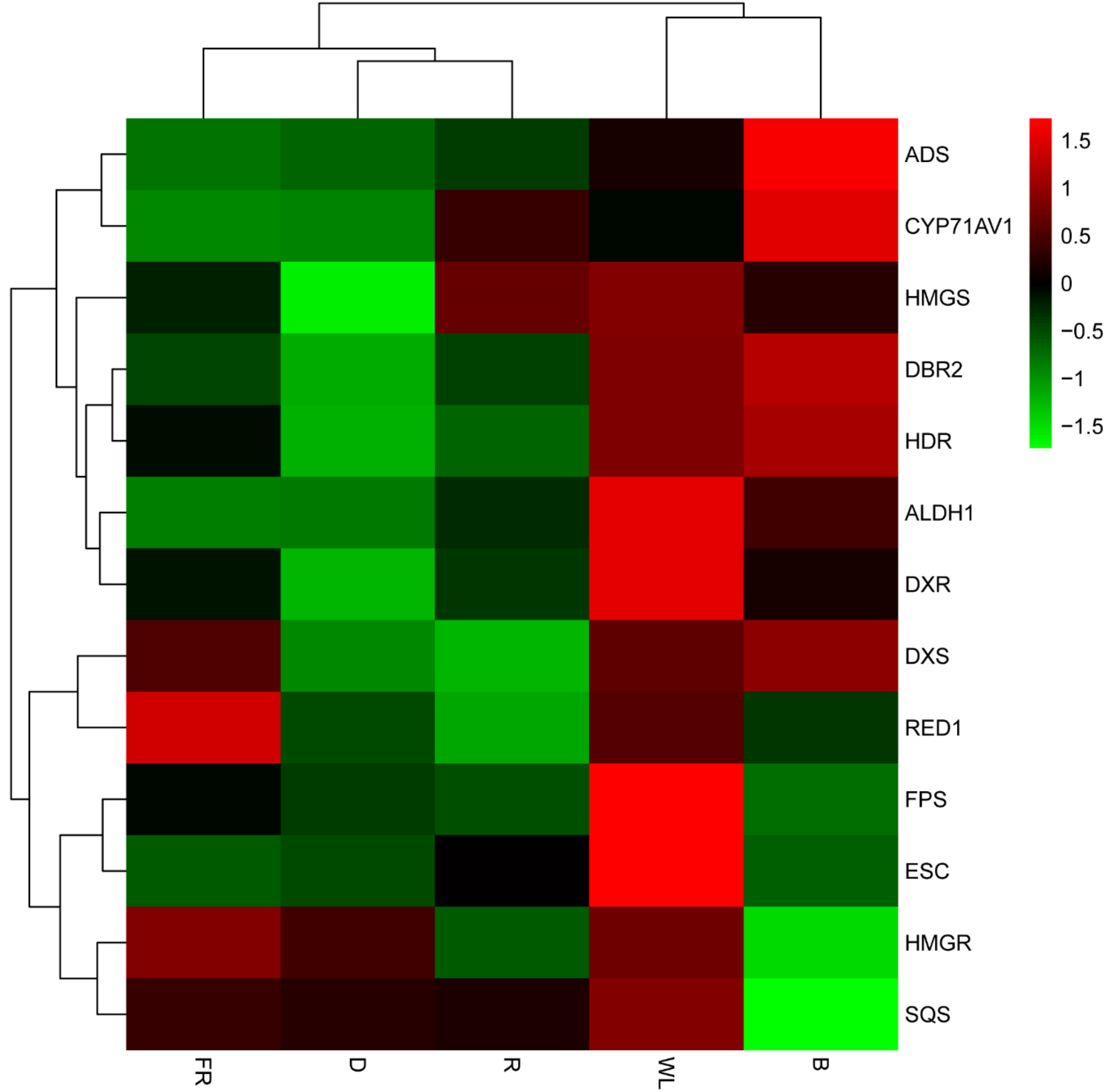
2.5. Comparative Analysis of Transcriptional Profiles
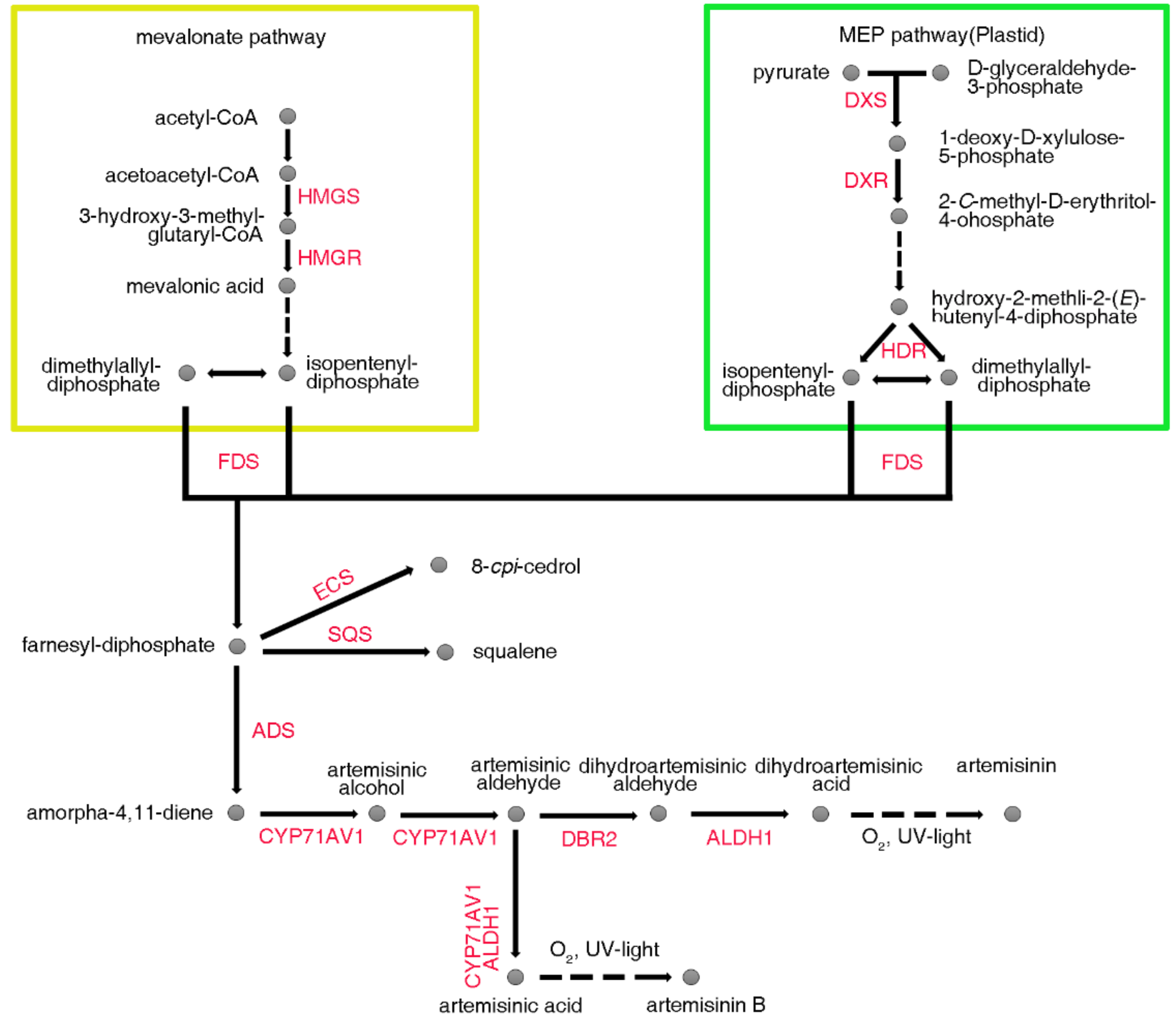
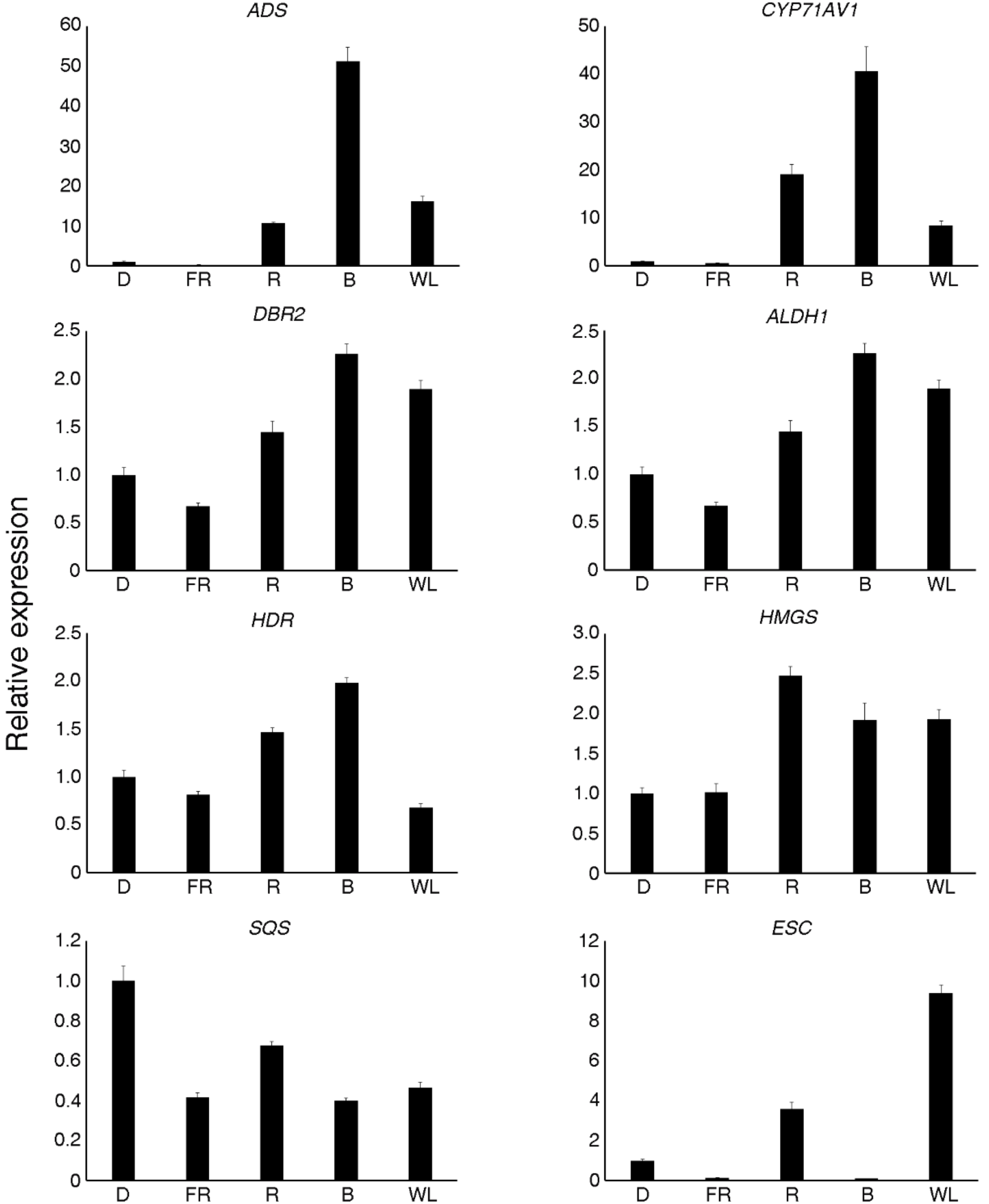
2.6. Expression Analysis of the Genes Involved in Artemisinin Biosynthesis
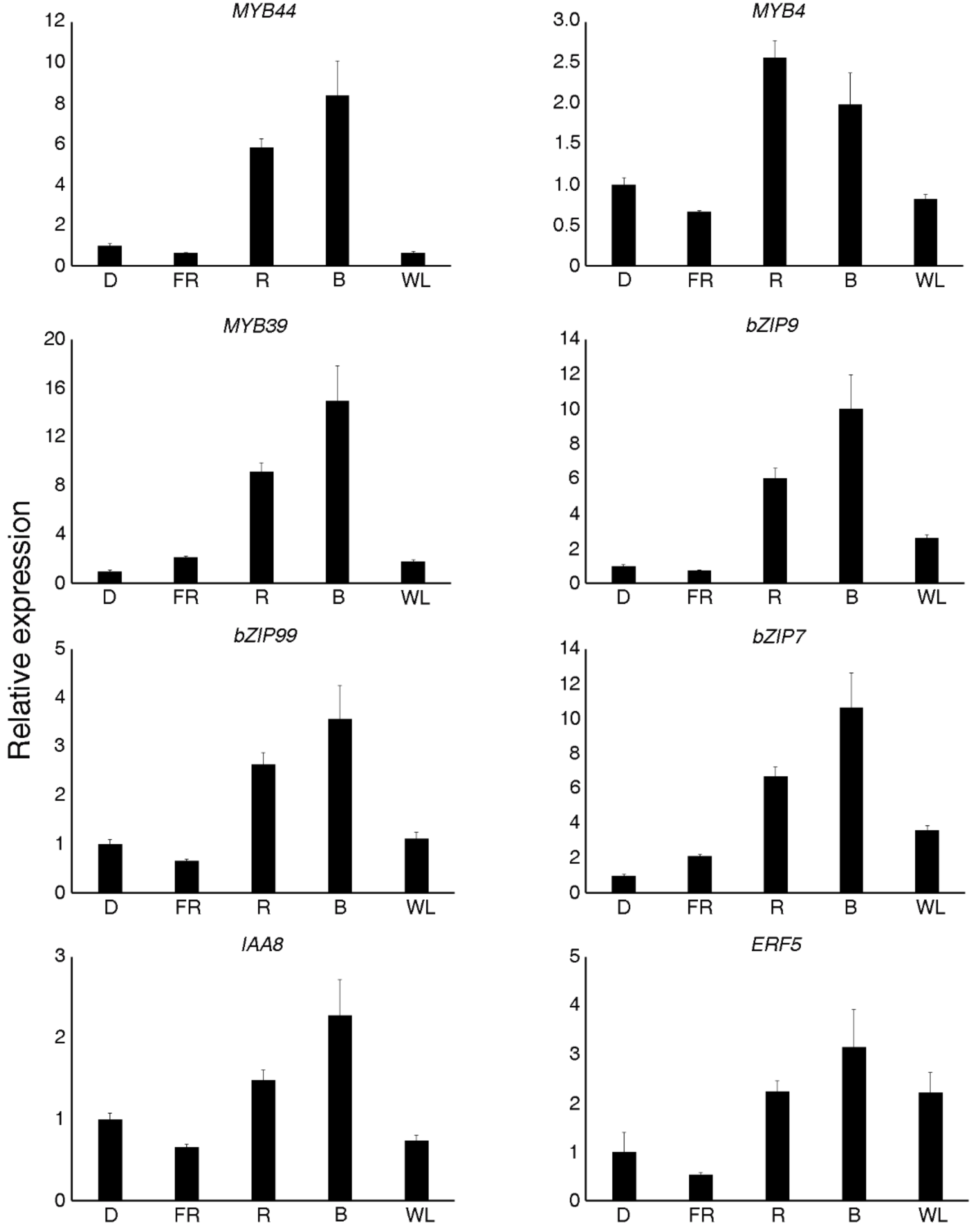
2.7. Validation of Differential Expression via Quantitative Real-Time PCR (qRT-PCR)
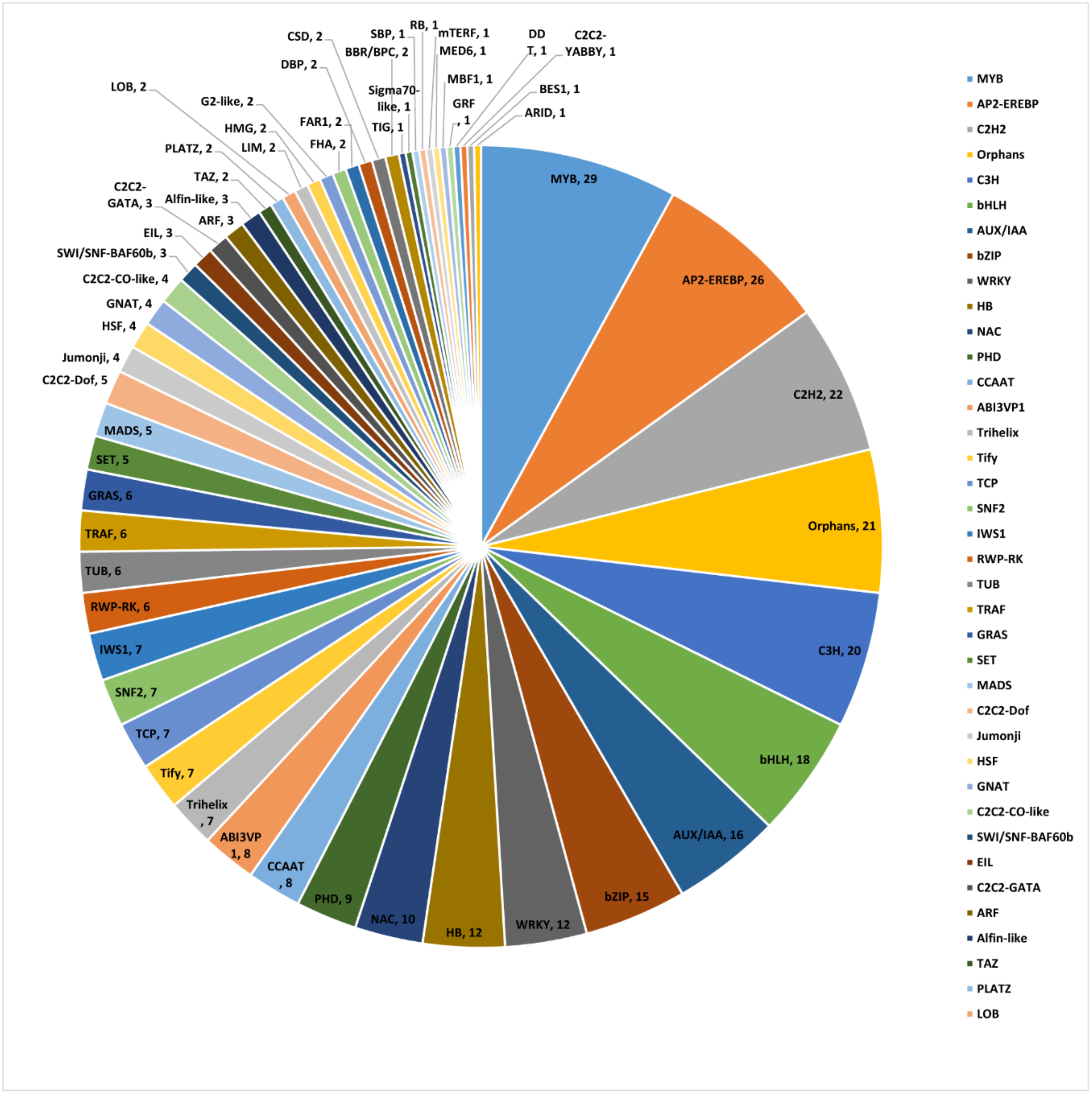
2.8. Identification of Co-Expressed TFs and Artemisinin Biosynthesis Genes
3. Materials and Methods
3.1. Plant Material and Growth Conditions
3.2. Chlorophyll Measurement
3.3. Transcriptome Sequencing
3.4. De Novo Assembly
3.5. Functional Annotation
3.6. Transcription Factor Identification and Co-Expression Analysis
3.7. Quantitative Real-Time PCR (qRT-PCR) Analysis of Gene Expression
3.8. Liquid Chromatography–Mass Spectrometry (LC–MS) Analysis of Secondary Metabolites
3.9. High-Performance Liquid Chromatography (HPLC) Conditions
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- World Health Organization. World Malaria Report 2015. 2015. Available online: http://www.who.int/mediacentre/factsheets/fs094/en/ (accessed on 9 December 2015).
- Dondorp, A.M.; Nosten, F.; Yi, P.; Das, D.; Phyo, A.P.; Tarning, J.; Lwin, K.M.; Ariey, F.; Hanpithakpong, W.; Lee, S.J.; et al. Artemisinin Resistance in Plasmodium falciparum Malaria. N. Engl. J. Med. 2009, 361, 455–467. [Google Scholar] [CrossRef] [PubMed]
- Bin, L.; Hong, Z. Advances on the study of the pharmacological effects of artemisinin and its derivatives. Chin. J. Clin. Pharmacol. Ther. 2010, 15, 572–576. [Google Scholar]
- Peplow, M. Malaria drug made in yeast causes market ferment. Nature 2013, 494, 160–161. [Google Scholar] [CrossRef] [PubMed]
- Paddon, C.J.; Westfall, P.J.; Pitera, D.J.; Benjamin, K.; Fisher, K.; McPhee, D.; Leavell, M.D.; Tai, A.; Main, A.; Eng, D.; et al. High-level semi-synthetic production of the potent antimalarial artemisinin. Nature 2013, 496, 528–532. [Google Scholar] [CrossRef] [PubMed]
- Aziz, A.; Poinssot, B.; Daire, X.; Adrian, M.; Bézier, A.; Lambert, B.; Joubert, J.M.; Pugin, A. Laminarin elicits defense responses in grapevine and induces protection against Botrytis cinerea and Plasmopara viticola. Mol. Plant-Microbe. Interact. 2003, 16, 1118–1128. [Google Scholar] [CrossRef] [PubMed]
- Korn, M.; Peterek, S.; Mock, H.P.; Heyer, A.G.; Hincha, D.K. Heterosis in the freezing tolerance, and sugar and flavonoid contents of crosses between Arabidopsis thaliana accessions of widely varying freezing tolerance. Plant Cell Environ. 2008, 31, 813–827. [Google Scholar] [CrossRef] [PubMed]
- Tsurunaga, Y.; Takahashi, T.; Katsube, T.; Kudo, A.; Kuramitsu, O.; Ishiwata, M.; Matsumoto, S. Effects of UV-B irradiation on the levels of anthocyanin, rutin and radical scavenging activity of buckwheat sprouts. Food Chem. 2013, 141, 552–556. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.Y.; Deng, X.W. Dissecting the phytochrome A-dependent signaling network in higher plants. Trends Plant Sci. 2003, 8, 172–178. [Google Scholar] [CrossRef]
- Jiao, Y.; Lau, O.S.; Deng, X.W. Light-regulated transcriptional networks in higher plants. Genetics 2007, 8, 217–230. [Google Scholar] [CrossRef] [PubMed]
- Lau, O.S.; Deng, X.W. Plant hormone signaling lightens up: Integrators of light and hormones. Curr. Opin. Plant Biol. 2010, 13, 571–577. [Google Scholar] [CrossRef] [PubMed]
- Wilson, M.I.; Greenberg, B.M. Protection of the D1 photosystem II reaction center protein from degradation in ultraviolet radiation following adaptation of Brassica napus L. to growth in ultraviolet radiation. Photochem. Photobiol. 1993, 57, 556–563. [Google Scholar] [CrossRef]
- Landrey, L.G.; Chapple, C.C.S.; Last, R.L. Arabidopsis mutants lacking phenolic sunscreens exhibit enhanced ultraviolet-B injury and oxidative damage. Plant Physiol. 1995, 109, 1159–1166. [Google Scholar] [CrossRef]
- Adrian, M.; Jeandet, P.; Douillet-Breuil, A.C.; Tesson, L.; Bessis, R. Stilbene content of 3 mature Vitis vinifera berries in response to UV-C elicitation. J. Agric. Food Chem. 2000, 48, 6103–6105. [Google Scholar] [CrossRef] [PubMed]
- Versari, A.; Parpinello, G.P.; Tornielli, G.B.; Ferrarini, R.; Giulivo, C. Stilbene compounds and stilbene synthase expression during ripening, wilting, and UV treatment in grape cv. Corvina. J. Agric. Food Chem. 2001, 49, 5531–5536. [Google Scholar] [CrossRef] [PubMed]
- Takayanagi, T.; Okuda, T.; Mine, Y.; Yokotsuka, K. Induction of resveratrol biosynthesis in skins of three grape cultivars by ultraviolet irradiation. J. Jpn. Soc. Hortic. Sci. 2004, 73, 193–199. [Google Scholar] [CrossRef]
- Nishikawa, Y.; Tominori, S.; Wada, K.; Kondo, H. Effect of cultivation practices on resveratrol content in grape berry skins. Hortic. Res. 2011, 10, 249–253. [Google Scholar] [CrossRef]
- Rai, R.; Meena, R.P.; Smita, S.S.; Shukla, A.; Rai, K.; Pandey-Rai, S. UV-B and UV-C pre-treatments induce physiological changes and artemisinin biosynthesis in Artemisia annua L.—An antimalarial plant. J. Photochem. Photobiol. B Biol. 2011, 105, 216–225. [Google Scholar] [CrossRef] [PubMed]
- Schramek, N.; Wang, H.; Römisch-Margl, W.; Keil, B.; Radykewicz, T.; Winzenhörlein, B.; Beerhues, L.; Bacher, A.; Rohdich, F.; Gershenzon, J.; et al. Artemisinin biosynthesis in growing plants of Artemisia annua. A 13CO2 study. Phytochemistry 2010, 71, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, K.T.; Arsenault, P.R.; Weathers, P.J. Trichomes + roots + ROS = artemisinin: Regulating artemisinin biosynthesis in Artemisia annua L. In Vitro Cell. Dev. Biol.-Plant 2011, 47, 329–338. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Han, J.; Kanagarajan, S.; Lundgren, A.; Brodelius, P.E. Trichome-specific expression of the amorpha-4,11-diene 12-hydroxylase (cyp71av1) gene, encoding a key enzyme of artemisinin biosynthesis in Artemisia annua, as reported by a promoter-GUS fusion. Plant Mol. Biol. 2013, 81, 119–138. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yang, K.; Jing, F.; Li, M.; Deng, T.; Huang, R.; Wang, B.; Wang, G.; Sun, X.; Tang, K.X. Cloning and Characterization of Trichome Specific Promoter of cpr71av1 Gene Involved in Artemisinin Biosynthesis in Artemisia annua L. Mol. Biol. 2011, 45, 751–758. [Google Scholar] [CrossRef]
- Teoh, K.H.; Polichuk, D.R.; Reed, D.W.; Nowak, G.; Covello, P.S. Artemisia annua L. (Asteraceae) trichome-specific cDNAs reveal CYP71AV1, a cytochrome P450 with a key role in the biosynthesis of the antimalarial sesquiterpene lactone artemisinin. FEBS Lett. 2006, 580, 1411–1416. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.; Lu, S.; Yu, Z.X.; Chen, X.Y. Research in biosynthesis of artemisinin (in Chinese). Sci. Technol. Rev. 2015, 33, 31–35. [Google Scholar]
- Yang, C.Q.; Fang, X.; Wu, X.M.; Mao, Y.; Wang, L.; Chen, X. Transcriptional regulation of plant secondary metabolism. J. Integr. Plant Biol. 2012, 54, 703–712. [Google Scholar] [CrossRef] [PubMed]
- Patra, B.; Schluttenhofer, C.; Wu, Y.; Pattanaik, S.; Yuan, L. Transcriptional regulation of secondary metabolite biosynthesis in plants. BBA-Gene Regul. Mech. 2013, 1829, 1236–1247. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.; Pu, G.; Lei, C.; Ma, L.; Wang, H.; Guo, Y.; Chen, J.; Du, Z.; Wang, H.; Li, G.; et al. Isolation and characterization of AaWRKY1, an Artemisia annua transcription factor that regulates the amorpha-4,11-diene synthase gene, a key gene of artemisinin biosynthesis. Plant Cell Physiol. 2009, 50, 2146–2161. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Zhang, L.; Zhang, F.; Jiang, W.; Shen, Q.; Zhang, L.; Lv, Z.; Wang, G.; Tang, K. AaORA, a trichome-specific AP2/ERF transcription factor of Artemisia annua, is a positive regulator in the artemisinin biosynthetic pathway and in disease resistance to Botrytis cinerea. New Phytol. 2013, 198, 1191–1202. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.; Xiao, J.; Shen, Y.; Ma, D.; Li, Z.; Pu, G.; Li, X.; Huang, L.; Liu, B.; Ye, H.; et al. Cloning and Characterization of AabHLH1, a bHLH Transcription Factor that Positively Regulates Artemisinin Biosynthesis in Artemisia annua. Plant Cell Physiol. 2014, 55, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Tang, K.X.; Shen, Q.; Lu, X.; Fu, X.Q.; Yan, T.X.; Sun, X.F.; Wang, G.F. Sweet Wormwood MYC2 Transcription Factor Protein Coding Sequence and Applications Thereof. China Patent ZL201310413155.3, 11 September 2013. [Google Scholar]
- Zhang, F.; Fu, X.; Lv, Z.; Lu, X.; Shen, Q.; Zhang, L.; Zhu, M.; Wang, G.; Sun, X.; Liao, Z.; et al. A Basic Leucine Zipper Transcription Factor, AabZIP1, Connects Abscisic Acid Signaling with Artemisinin Biosynthesis in Artemisia annua L. Mol. Plant 2015, 8, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Rockwell, N.C.; Su, Y.S.; Lagarias, J.C. Phytochrome structure and signaling mechanisms. Annu. Rev. Plant Biol. 2006, 57, 837–858. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Robertson, D.E.; Ahmad, M.; Raibekas, A.A.; Jorns, M.S.; Dutton, P.L.; Cashmore, A.R. Association of flavin adenine dinucleotide with the Arabidopsis blue light receptor CRY1. Science 1995, 269, 968–970. [Google Scholar] [CrossRef] [PubMed]
- Malhotra, K.; Kim, S.T.; Batschauer, A.; Dawut, L.; Sancar, A. Putative blue-light photoreceptors from Arabidopsis thaliana and Sinapis alba with a high degree of sequence homology to DNA photolyase contain the two photolyase cofactors but lack DNA repair activity. Biochemistry 1995, 34, 6892–6899. [Google Scholar] [CrossRef] [PubMed]
- Hsu, D.S.; Zhao, X.; Zhao, S.; Kazantsev, A.; Wang, R.P.; Todo, T.; Wei, Y.F.; Sancar, A. Putative human blue-light photoreceptors hCRY1 and hCRY2 are flavoproteins. Biochemistry 1996, 35, 13871–13877. [Google Scholar] [CrossRef] [PubMed]
- Brown, B.A.; Cloix, C.; Jiang, G.H.; Kaiserli, E.; Herzyk, P.; Kliebenstein, D.J.; Jenkins, G.I. A UV-B-specific signaling component orchestrates plant UV protection. Proc. Natl. Acad. Sci. USA 2005, 102, 18225–18230. [Google Scholar] [CrossRef] [PubMed]
- Moore, R.; Clark, W.D.; Kingsley, R.S.; Vodopich, D. Plant physiology and growth. In Botany; Wm. C. Brown Pub: Dubuque, IA, USA, 1995; Chapter 7; ISBN 9780697037756. [Google Scholar]
- Nafis, T.; Akmal, M.; Ram, M.; Alam, P.; Ahlawat, S.; Mohd, A.; Abdin, M.Z. Enhancement of artemisinin content by constitutive expression of the HMG-CoA reductase gene in highyielding strain of Artemisia annua L. Plant Biotechnol. Rep. 2011, 5, 53–60. [Google Scholar] [CrossRef]
- Cordoba, E.; Salmi, M.; León, P. Unravelling the regulatory mechanisms that modulate the MEP pathway in higher plants. J. Exp. Bot. 2009, 60, 2933–2943. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.; Li, G.; Zhu, Y.; Xie, D.Y. Overexpression and Suppression of Artemisia annua 4-Hydroxy- 3-Methylbut-2-enyl Diphosphate Reductase 1 Gene (AaHDR1) Differentially Regulate Artemisinin and Terpenoid Biosynthesis. Front. Plant Sci. 2017, 8, 77. [Google Scholar] [CrossRef] [PubMed]
- Banyai, W.; Kirdmanee, C.; Mii, M.; Supaibulwatana, K. Overexpression of farnesyl pyrophosphate synthase (FPS) gene affected artemisinin content and growth of Artemisia annua L. Plant Cell Tissue Organ. Cult. 2010, 103, 255–265. [Google Scholar] [CrossRef]
- Jing, F.; Zhang, L.; Li, M.; Tang, K. Over-expressing CYP71AV1 and CPR genes enhances artemisinin content in Artemisia annua L. J. Agric. Sci. Technol. 2008, 10, 64–70. [Google Scholar]
- Yuan, Y.; Liu, W.; Zhang, Q.; Xiang, L.; Liu, X.; Chen, M.; Lin, Z.; Wang, Q.; Liao, Z. Overexpression of artemisinic aldehyde Δ11(13) reductase gene-enhanced artemisinin and its relative metabolite biosynthesis in transgenic Artemisia annua L. Biotechnol. Appl. Biochem. 2015, 62, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Jing, F.Y.; Li, F.P.; Li, M.Y.; Wang, Y.L.; Wang, G.F.; Sun, X.F.; Tang, K.X. Development of transgenic Artemisia annua (Chinese wormwood) plants with an enhanced content of artemisinin, an effective anti-malarial drug, by hairpin-RNA-mediated gene silencing. Biotechnol. Appl. Biochem. 2009, 52, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Osterlund, M.T.; Hardtke, C.S.; Wei, N.; Deng, X.W. Targeted destabilization of HY5 during light-regulated development of Arabidopsis. Nature 2000, 405, 462–466. [Google Scholar] [CrossRef] [PubMed]
- Saijo, Y.; Sullivan, J.A.; Wang, H.; Yang, J.; Shen, Y.; Rubio, V.; Ma, L.; Hoecker, U.; Deng, X.W. The COP1-SPA1 interaction defines a critical step in phytochrome A-mediated regulation of HY5 activity. Genes Dev. 2003, 17, 2642–2647. [Google Scholar] [CrossRef] [PubMed]
- Heim, M.A.; Jakoby, M.; Werber, M.; Martin, C.; Weisshaar, B.; Bailey, P.C. The basic helix-loop-helix transcription factor family in plants: A genome-wide study of protein structure and functional diversity. Mol. Biol. Evol. 2003, 20, 735–747. [Google Scholar] [CrossRef] [PubMed]
- Rushton, D.L.; Tripathi, P.; Rabara, R.; Liu, J.; Ringler, P.; Boken, A.K.; Langum, T.J.; Smidt, L.; Boomsma, D.D.; Emme, N.J.; et al. WRKY transcription factors: Key components in abscisic acid signalling. Plant Biotechnol. J. 2012, 10, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Lin, R.; Ding, L.; Casola, C.; Ripoll, D.R.; Feschotte, C.; Wang, H. Transposase-derived transcription factors regulate light signaling in Arabidopsis. Science 2007, 318, 1302–1305. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Song, C.; Song, L.; Shang, Z.; Yang, S.; Zhang, D.; Sun, W.; Shen, Q.; Zhao, D. RNA Sequencing and Coexpression Analysis Reveal Key Genes Involved in α-Linolenic Acid Biosynthesis in Perilla frutescens Seed. Int. J. Mol. Sci. 2017, 18, 2433. [Google Scholar] [CrossRef] [PubMed]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q. Trinity: Reconstructing a full-length transcriptome without a genome from RNA-Seq data. Nat. Biotechnol. 2013, 29, 644–652. [Google Scholar] [CrossRef] [PubMed]
- Götz, S.; García-Gómez, J.M.; Terol, J.; Williams, T.D.; Nagaraj, S.H.; Nueda, M.J.; Robles, M.; Talón, M.; Dopazo, J.; Conesa, A. High-throughput functional annotation and data mining with the Blast2GO suite. Nucleic Acids Res. 2008, 36, 3420–3435. [Google Scholar] [CrossRef] [PubMed]
- Ariani, A.; Gepts, P. Genome-wide identification and characterization of aquaporin gene family in common bean (Phaseolus vulgaris L.). Mol. Genet. Genom. 2015, 290, 1771. [Google Scholar] [CrossRef] [PubMed]
- Smoot, M.; Ono, K.; Ideker, T.; Maere, S. PiNGO: A Cytoscape plugin to find candidate genes in biological networks. Bioinformatics 2011, 27, 1030–1031. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are available from the authors. |









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Clean Reads (n) | Clean Bases (n) | Q20 (%) | Q30 (%) | GC Content (%) |
|---|---|---|---|---|---|
| D1 | 41,315,654 | 39,620,944 | 97.63 | 93.66 | 42.76 |
| D2 | 44,4941,98 | 42,460,928 | 97.32 | 92.96 | 42.92 |
| D3 | 45,997,162 | 44,196,856 | 97.62 | 93.63 | 42.85 |
| FR1 | 44,583,758 | 42,993,870 | 97.60 | 93.58 | 42.92 |
| FR2 | 46,616,436 | 44,924,628 | 97.65 | 93.80 | 42.92 |
| FR3 | 53,563,708 | 51,433,966 | 97.16 | 92.79 | 42.93 |
| WL1 | 48,061,222 | 46,239,326 | 97.56 | 93.62 | 42.67 |
| WL2 | 41,232,122 | 39,771,772 | 97.63 | 93.77 | 42.75 |
| WL3 | 51,498,242 | 49,412,744 | 97.68 | 93.87 | 42.82 |
| R1 | 46,278,940 | 44,459,758 | 96.90 | 92.12 | 42.62 |
| R2 | 50,660,678 | 48,157,446 | 97.32 | 93.01 | 42.55 |
| R3 | 50,543,012 | 47,291,920 | 97.21 | 92.90 | 42.61 |
| B1 | 63,937,458 | 61,502,154 | 97.95 | 94.64 | 42.50 |
| B2 | 60,062,102 | 57,833,940 | 97.91 | 94.54 | 42.52 |
| B3 | 55,204,498 | 52,916,964 | 97.59 | 93.60 | 42.51 |
| Database | Number of Unigenes | Percentage (%) |
|---|---|---|
| NR | 90,049 | 33.11 |
| NT | 67,311 | 24.75 |
| KEGG | 37,459 | 13.77 |
| Swiss-Prot | 83,506 | 30.70 |
| PFAM | 85,702 | 31.51 |
| GO | 87,236 | 32.08 |
| KOG | 36,681 | 13.48 |
| Annotated in all of the databases | 13,326 | 4.90 |
| Annotated in at least one database | 136,365 | 50.14 |
| Total Unigenes | 271,932 | 100.00 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, D.; Sun, W.; Shi, Y.; Wu, L.; Zhang, T.; Xiang, L. Red and Blue Light Promote the Accumulation of Artemisinin in Artemisia annua L. Molecules 2018, 23, 1329. https://doi.org/10.3390/molecules23061329
Zhang D, Sun W, Shi Y, Wu L, Zhang T, Xiang L. Red and Blue Light Promote the Accumulation of Artemisinin in Artemisia annua L. Molecules. 2018; 23(6):1329. https://doi.org/10.3390/molecules23061329
Chicago/Turabian StyleZhang, Dong, Wei Sun, Yuhua Shi, Lan Wu, Tianyuan Zhang, and Li Xiang. 2018. "Red and Blue Light Promote the Accumulation of Artemisinin in Artemisia annua L." Molecules 23, no. 6: 1329. https://doi.org/10.3390/molecules23061329
APA StyleZhang, D., Sun, W., Shi, Y., Wu, L., Zhang, T., & Xiang, L. (2018). Red and Blue Light Promote the Accumulation of Artemisinin in Artemisia annua L. Molecules, 23(6), 1329. https://doi.org/10.3390/molecules23061329