Effect of Sterilization Process and Storage on the Antioxidative Properties of Runner Bean


Abstract
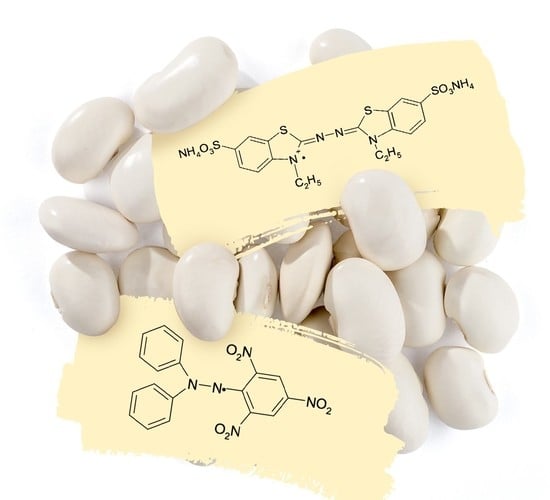
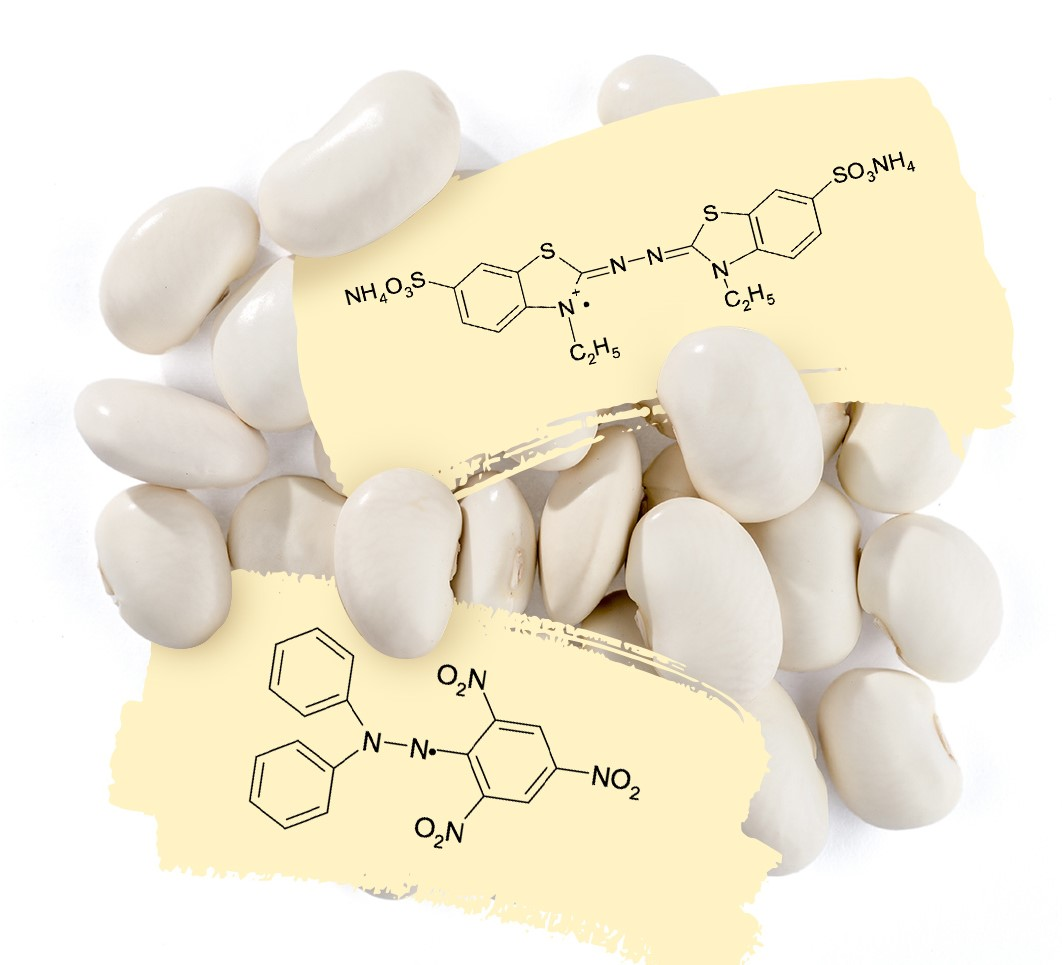
:
1. Introduction
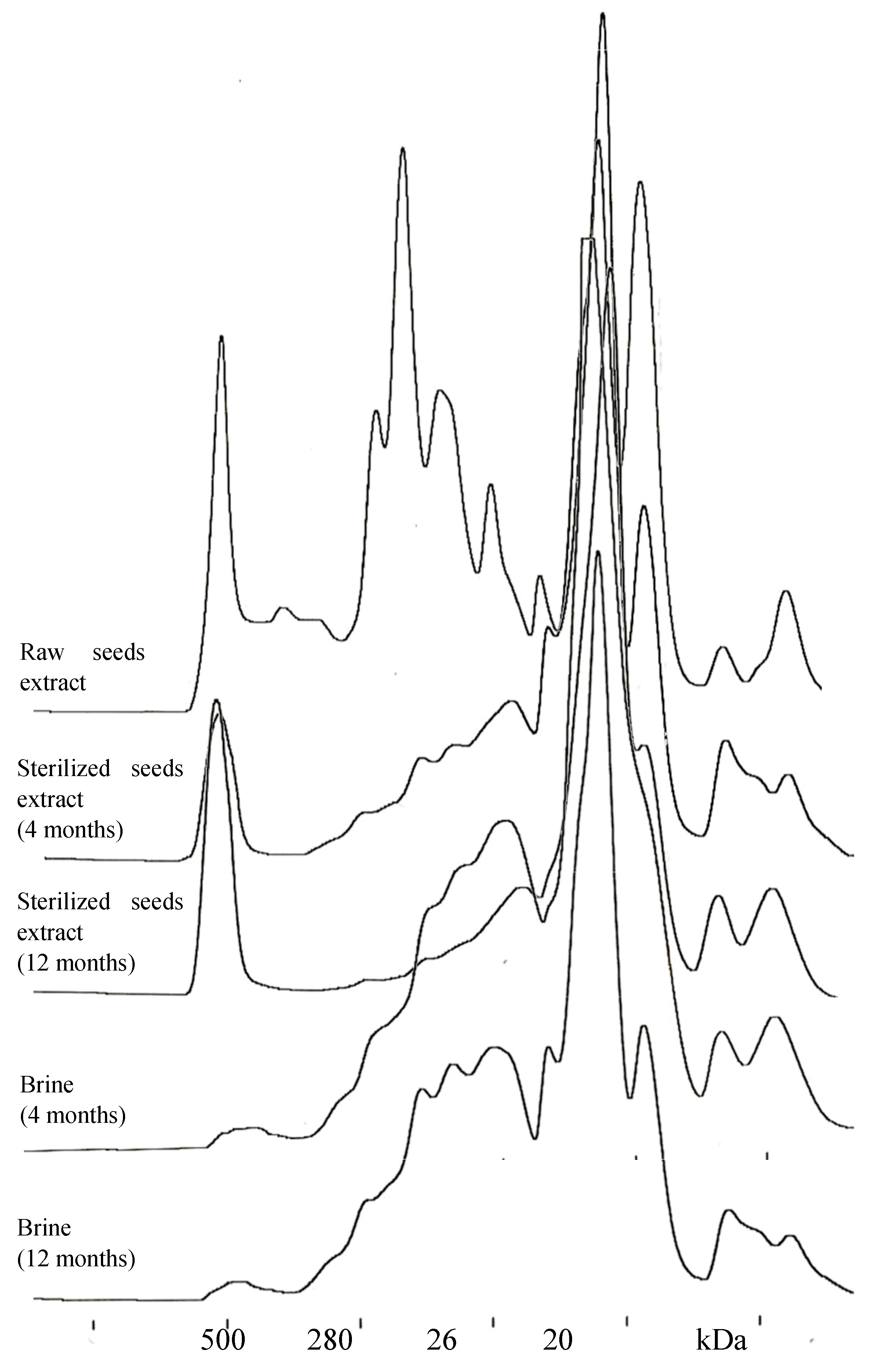
2. Results
3. Materials and Methods
3.1. Plant Material
3.2. Material Preparation
3.3. Chemical Composition
3.4. Determination of Condensed Tannins Content (CTC)
3.5. Determination of Total Polyphenols Content (TPC)
3.6. Size-Exclusion HPLC
3.7. Chelating Ability
3.8. Antiradical and Antioxidant Activity
3.9. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Vaz-Patto, M.; Amarowicz, R.; Aryee, A.N.A.; Boye, J.I.; Chung, H.J.; Martin-Cabrejas, M.A.; Domoney, C. Achievements and challengers in improving the nutritional quality of food legumes. Crit. Rev. Plant Sci. 2015, 34, 105–143. [Google Scholar] [CrossRef]
- Marathe, S.A.; Rajalakshimi, V.; Jamdar, S.N.; Sharma, A. Comparative study on antioxiadant activity of different varieties of commonly consumed legumes in India. Food Chem. Toxicol. 2011, 49, 2005–2012. [Google Scholar] [CrossRef] [PubMed]
- Akond, G.M.; Khandaker, L.; Berthold, J.; Gates, L.; Peters, K.; Delong, H.; Hossain, K. Anthocyanin, total polyphenols and antioxidant activity of common bean. Am. J. Food Technol. 2011, 6, 385–394. [Google Scholar] [CrossRef]
- Druzynska, B. The use of fluorometric method to study the antioxidative properties of legume seeds—Short report. Pol. J. Food Nutr. Sci. 2000, 50, 47–51. [Google Scholar]
- Wolosiak, R. Pea and bean albumins as antioxidants in linoleic acid model system. In Proceedings of the 4th European Conference on Grain Legumes, Cracow, Poland, 8–10 July 2001; pp. 405–406. [Google Scholar]
- Worobiej, E. Antioxidative activity of broad bean proteins and changes in proteins induced by radicals. In Proceedings of the 4th European Conference on Grain Legumes, Cracow, Poland, 8–10 July 2001; pp. 406–407. [Google Scholar]
- Amiri, A.; Amiri, A. Antioxidants and heart disease; current knowledge. Ann. Res. Antioxid. 2017, 2, 1–2. [Google Scholar]
- Cai, Y.; Luo, Q.; Sun, M.; Corke, H. Antioxidant activity in phenolic compounds of 112 traditional Chinese medical plants associated with anticancer. Life Sci. 2004, 74, 2157–2184. [Google Scholar] [CrossRef] [PubMed]
- Ganesan, K.; Xu, B.J. Polyphenol-rich dry common beans (Phaseolus vulgaris L.) and their health benefits. Int. J. Mol. Sci. 2017, 18, 2331. [Google Scholar] [CrossRef] [PubMed]
- Umeda, W.M.; Luzia, D.M.M.; Jorge, N. Evaluation of bioactive compounds in bean oils (Phaseolus vulgaris L.), Perola and BRS Valente varieties. Curr. Nutr. Food Sci. 2018, 14, 40–46. [Google Scholar] [CrossRef]
- Duenas, M.; Sarmento, T.; Aguilera, Y.; Benitez, V.; Molla, E.; Esteban, R.M.; Martin-Cabarejas, M. Impact of cooking and germination on phenolic composition and dietary fibre fractions in dark bens (Phaseolus Vulgaris L.) and lentils (Lens culinaris L.). LWT-Food Sci. Technol. 2016, 66, 72–78. [Google Scholar] [CrossRef]
- Labuda, H. Runner bean (Phaseolus coccineus L.)—Biology and use. Acta Sci. Pol. Hortorum 2010, 9, 117–132. [Google Scholar]
- Schwember, A.R.; Carrasco, B.; Gepts, P. Unraveling agronomic and genetic aspects of runner bean (Phaseolus coccineus L.). Field Crop Res. 2017, 206, 86–94. [Google Scholar] [CrossRef]
- Avasilcai, L.; Teliban, G.; Morariu, D.I.; Stoleru, V.; Bibire, N.; Vieriu, M.; Panainte, A.D.; Munteanu, N. Parameters of chemical composition of Phaseolus coccineus L. pods grown in protected areas. Rev. Chim. 2017, 68, 2955–2958. [Google Scholar]
- Hamburdă, S.B.; Teliban, G.C.; Munteanu, N.; Stoleru, V. Effect of intercropping system on the quality and quantity of runner bean (Phaseolus coccineus L.). Not. Bot. Horti Agrobot. 2016, 44, 613–618. [Google Scholar] [CrossRef]
- Pan, W.L.; Ng, T.B. A dimeric Phaseolus coccineus lectin with anti-oxidative, anti-proliferative and cytokine-inducing activities. Int. J. Biol. Macromol. 2015, 81, 960–966. [Google Scholar] [CrossRef] [PubMed]
- Worobiej, E.; Wołosiak, R.; Drużyńska, B. Antioxidant properties of globulin preparations from the seeds of chosen leguminous species. In Food Quality and Safety; Krasnowska, G., Pęksa, A., Eds.; Wydawnictwo Uniwersytetu Przyrodniczego we Wrocławiu: Wrocław, Poland, 2009; pp. 125–132. ISSN 1898-1151. [Google Scholar]
- Piecyk, M.; Drużyńska, B.; Worobiej, E.; Wołosiak, R.; Ostrowska-Ligęza, E. Effect of hydrothermal treatment of runner bean (Phaseolus coccineus) seeds and starch isolation on starch digestibility. Food Res. Int. 2013, 50, 428–437. [Google Scholar] [CrossRef]
- Ovando-Martinez, M.; Osorio-Diaz, P.; Whitney, K.; Bello-Pérez, L.A.; Simsek, S. Effect of the cooking on physicochemical and starch digestibility properties of two varieties of common bean (Phaseolus vulgaris L.) grown under different water regimes. Food Chem. 2011, 129, 358–365. [Google Scholar] [CrossRef]
- Piecyk, M.; Wołosiak, R.; Drużyńska, B.; Worobiej, E. Chemical composition and starch digestibility in flour from Polish processed legume seeds. Food Chem. 2012, 135, 1057–1064. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Hatcher, D.W.; Tyler, R.T.; Toews, R.; Gawalko, E.J. Effect of cooking on the composition of beans (Phaseolus vulgaris L.) and chickpeas (Cicer arietinum L.). Food Res. Int. 2010, 43, 589–594. [Google Scholar] [CrossRef]
- Carbonaro, M.; Nicoli, S.; Musci, G. Heat-induced aggregation of Phaseolus vulgaris L. proteins: An electron spin resonance study. J. Agric. Food Chem. 1999, 47, 2188–2192. [Google Scholar] [CrossRef] [PubMed]
- Gueguen, J. Developments in Food Proteins-6; Elsevier Applied Science: London, UK; New York, NY, USA, 1991. [Google Scholar]
- Lawrence, M.C.; Izard, T.; Beuchat, M.; Blagrove, R.J.; Colman, P.M. Structure of Phaseolin at 2.2 Å Resolution: Implications for a Common Vicilin/Legumin Structure and the Genetic Engineering of Seed Storage Proteins. J. Mol. Biol. 1994, 238, 748–776. [Google Scholar] [CrossRef] [PubMed]
- Nyau, V.; Prakash, S.; Rodrigues, J.; Farrant, J. Domestic cooking effects of bambara groundnuts and common beans in the antioxidant properties and polyphenol profiles. J. Food Res. 2017, 6, 1–14. [Google Scholar] [CrossRef]
- Parmar, N.; Singh, N.; Kaur, A.; Virdi, A.S.; Thakur, S. Effect of canning on color, protein and phenolic profile of grains from kidney bean, field pea and chickpea. Food Res. Int. 2016, 89, 526–532. [Google Scholar] [CrossRef] [PubMed]
- Ismail, A.; Marjan, Z.M.; Foong, C. Total antioxidant activity and phenolic content in selected vegetables. Food Chem. 2004, 87, 581–586. [Google Scholar] [CrossRef]
- Repo-Carrasco-Valencia, R.; de la Cruz, A.A.; Alvarez, J.C.I.; Kallio, H. Chemical and functional characterization of kaniwa (Chenopodium pallidicaule) grain, extrudate and bran. Plant Foods Hum. Nutr. 2009, 64, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Wolosiak, R.; Druzynska, B.; Piecyk, M.; Worobiej, E.; Majewska, E.; Lewicki, P.P. Influence of industrial sterilization, freezing and steam cooking on antioxidant properties of green peas and string beans. Int. J. Food Sci. Technol. 2011, 46, 93–100. [Google Scholar] [CrossRef]
- Arcan, I.; Yemenicioğlu, A. Antioxidant activity of protein extracts from heat-treated or thermal processed chickpeas and white beans. Food Chem. 2007, 103, 301–312. [Google Scholar] [CrossRef]
- Lisiewska, Z.; Kmiecik, W.; Korus, A. Content of nitrogen compounds in raw and preserved seeds of grass pea (Lathyrus sativus L.). Eur. Food Res. Technol. 2000, 213, 343–348. [Google Scholar] [CrossRef]
- Wolosiak, R.; Worobiej, E.; Piecyk, M.; Druzynska, B.; Nowak, D.; Lewicki, P.P. Activities of amine and phenolic antioxidants and their changes in broad beans (Vicia faba) after freezing and steam cooking. Int. J. Food Sci. Technol. 2010, 45, 29–37. [Google Scholar] [CrossRef]
- Siddhuraju, P.; Becker, K. The antioxidant and free radical scavenging activities of processed cowpea (Vigna unguiculata (L.) Walp.) seed extracts. Food Chem. 2007, 101, 10–19. [Google Scholar] [CrossRef]
- Nithiyanantham, S.; Selvakumar, S.; Siddhuraju, P. Total phenolic content and antioxidant activity of two different solvent extracts from raw and processed legumes, Cicer arietinum L. and Pisum sativum L. J. Food Compos. Anal. 2012, 27, 52–60. [Google Scholar] [CrossRef]
- Xu, B.J.; Chang, S.K.C. Effect of soaking, boiling, and steaming on total phenolic content and antioxidant activities of cool season food legumes. Food Chem. 2008, 110, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Association of Official Analytical Chemists (AOAC). Official Methods of Analysis of the Association of Official Analytical Chemists, 15th ed.; Association of Official Analytical Chemists (AOAC): Arlington, VA, USA, 1990. [Google Scholar]
- Price, M.L.; van Scoyoc, S.; Butler, L.G. A critical evaluation of the vanillin reaction as an assay for tannin in sorgum grain. J. Agric. Food Chem. 1978, 26, 1214–1218. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybolic-phosphotungstic acid reagents. Am. J. Enol. Vitac. 1965, 16, 144–158. [Google Scholar]
- Andrews, P. Estimation of molecular weights of proteins by Sephadex gel-filtration. Biochem. J. 1964, 91, 222–233. [Google Scholar] [CrossRef] [PubMed]
- Lai, L.S.; Chou, S.T.; Chao, W.W. Studies on the antioxidative activities of hsian-tsao leaf gum. J. Agric. Food Chem. 2001, 49, 963–968. [Google Scholar] [CrossRef] [PubMed]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Rad. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Yen, G.C.; Chen, H.Y. Antioxidant activity of various tea extracts in relation to their antimutagenicity. J. Agric. Food Chem. 1995, 43, 27–32. [Google Scholar] [CrossRef]
- Kuo, J.M.; Yeh, D.B.; Pan, B.S. Rapid photometric assay evaluating antioxidative activity in edible plant material. J. Agric. Food Chem. 1999, 47, 3206–3209. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the investigated compounds are not available from the authors. |


{kind=link}
{kind=link}
| Sample | Total N g/100 g d.m. | Ash g/100 g d.m. | Dry Matter g/100 g | |
|---|---|---|---|---|
| Raw seeds | 3.8 ± 0.1 b | 4.12 ± 0.01 b | 89.2 ± 0.1 e | |
| Stored 4 months | sterilized seeds | 5.0 ± 0.1 c | 1.29 ± 0.04 a | 30.2 ± 0.3 c |
| brine | 2.9 ± 0.3 a | nd | 7.8 ± 0.1 b | |
| Stored 12 months | sterilized seeds | 3.7 ± 0.1 b | 1.28 ± 0.01 a | 32.3 ± 0.1 d |
| brine | 3.6 ± 0.1 b | nd | 6.7 ± 0.2 a | |
| Sample | Soluble N g/100 g d.m. | Non-Protein N g/100 g d.m. | Chelating Ability µmol Fe/g d.m. | |
|---|---|---|---|---|
| Raw seeds | 1.52 ± 0.02 c | 0.38 ± 0.0 b | 1.94 ± 0.02 a | |
| Stored 4 months | sterilized seeds | 0.64 ± 0.01 b | 0.61 ± 0.0 c | 5.84 ± 0.01 c |
| brine | nd | 1.67 ± 0.0 e | 2.17 ± 0.03 b | |
| Stored 12 months | sterilized seeds | 0.26 ± 0.01 a | 0.17 ± 0.0 a | 6.04 ± 0.03 c |
| brine | nd | 1.11 ± 0.0 d | 5.98 ± 0.01 c | |
| Sample | Total Polyphenols g GAE/100 g d.m. | Condensed Tannins g CAE/100 g d.m. | Chelating Ability µmol Fe/g d.m. | |
|---|---|---|---|---|
| Raw seeds | 1.62 ± 1.54 c | 0.65 ± 1.21 d | 1.88 ± 0.03 a | |
| Stored 4 months | sterilized seeds | 1.14 ± 1.87 b | 0.56 ± 1.26 c | 5.30 ± 0.06 b |
| brine | nd | 0.01 ± 0.01 a | nd | |
| Stored 12 months | sterilized seeds | 0.88 ± 1.45 a | 0.46 ± 1.14 b | 5.73 ± 0.09 b |
| brine | nd | 0.01 ± 0.01 a | nd | |
| Sample | Water Extract | 70% Acetone Extract | |||
|---|---|---|---|---|---|
| mg Trolox/100 g | mg Trolox/100 g d.m. | mg Trolox/100 g | mg Trolox/100 g d.m. | ||
| Raw seeds | 355.4 ± 3.7 e | 398.4 ± 4.2 d | 192.0 ± 1.2 c | 215.3 ± 1.4 c | |
| Stored 4 months | sterilized seeds | 50.9 ± 1.8 b | 168.7 ± 6.1 a | 33.9 ± 4.1 a | 112.3 ± 13.4 a |
| brine | 23.7 ± 0.4 a | 304.4 ± 5.7 c | nd | nd | |
| Stored 12 months | sterilized seeds | 65.7 ± 3.3 d | 203.4 ± 10.1 b | 44.7 ± 1.4 b | 138.3 ± 4.2 b |
| brine | 61.0 ± 0.6 c | 910.5 ± 8.2 e | nd | nd | |
| Sample | Water Extract | 70% Acetone Extract | |||
|---|---|---|---|---|---|
| mg Trolox /100 g | mg Trolox/100 g d.m. | mg Trolox/100 g | mg Trolox/100 g d.m. | ||
| Raw seeds | 38.0 ± 0.1 e | 42.7 ± 0.5 b | 402.9 ± 27.1 b | 451.7 ± 30.3 b | |
| Stored 4 months | sterilized seeds | 4.6 ± 0.1 b | 15.3 ± 0.2 a | 5.8 ± 0.1 a | 19.2 ± 0.2 a |
| brine | 7.1 ± 0.2 c | 91.0 ± 2.1 c | nd | nd | |
| Stored 12 months | sterilized seeds | 4.0 ± 0.2 a | 12.5 ± 0.7 a | 6.1 ± 0.2 a | 19.0 ± 0.7 a |
| brine | 7.6 ± 0.4 d | 112.8 ± 5.9 d | nd | nd | |
| Sample | Water Extract | 70% Acetone Extract | |||
|---|---|---|---|---|---|
| LOOH | LOX | LOOH | LOX | ||
| Raw seeds | 36.6 ± 0.8 c | 17.0 ± 4.5 c | 5.3 ± 1.0 b | 32.2 ± 9.7 b | |
| Stored 4 months | sterilized seeds | 4.8 ± 0.4 a | 6.8 ± 2.1 b | 1.6 ± 1.4 a | 23.2 ± 7.1 b |
| brine | 3.5 ± 1.6 a | 0.8 ± 2.7 a | nd | nd | |
| Stored 12 months | sterilized seeds | 3.9 ± 3.1 a | 1.4 ± 2.0 a | 4.8 ± 1.4 b | 8.9 ± 3.5 a |
| brine | 10.1 ± 3.2 b | 3.3 ± 1.9 a,b | nd | nd | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wołosiak, R.; Drużyńska, B.; Piecyk, M.; Majewska, E.; Worobiej, E. Effect of Sterilization Process and Storage on the Antioxidative Properties of Runner Bean. Molecules 2018, 23, 1409. https://doi.org/10.3390/molecules23061409
Wołosiak R, Drużyńska B, Piecyk M, Majewska E, Worobiej E. Effect of Sterilization Process and Storage on the Antioxidative Properties of Runner Bean. Molecules. 2018; 23(6):1409. https://doi.org/10.3390/molecules23061409
Chicago/Turabian StyleWołosiak, Rafał, Beata Drużyńska, Małgorzata Piecyk, Ewa Majewska, and Elwira Worobiej. 2018. "Effect of Sterilization Process and Storage on the Antioxidative Properties of Runner Bean" Molecules 23, no. 6: 1409. https://doi.org/10.3390/molecules23061409
APA StyleWołosiak, R., Drużyńska, B., Piecyk, M., Majewska, E., & Worobiej, E. (2018). Effect of Sterilization Process and Storage on the Antioxidative Properties of Runner Bean. Molecules, 23(6), 1409. https://doi.org/10.3390/molecules23061409