Physicochemical Properties of C-Type Starch from Root Tuber of Apios fortunei in Comparison with Maize, Potato, and Pea Starches



Abstract
:1. Introduction
2. Results and Discussion
2.1. Starch and Soluble Sugar Contents in Root Tuber of A. fortunei
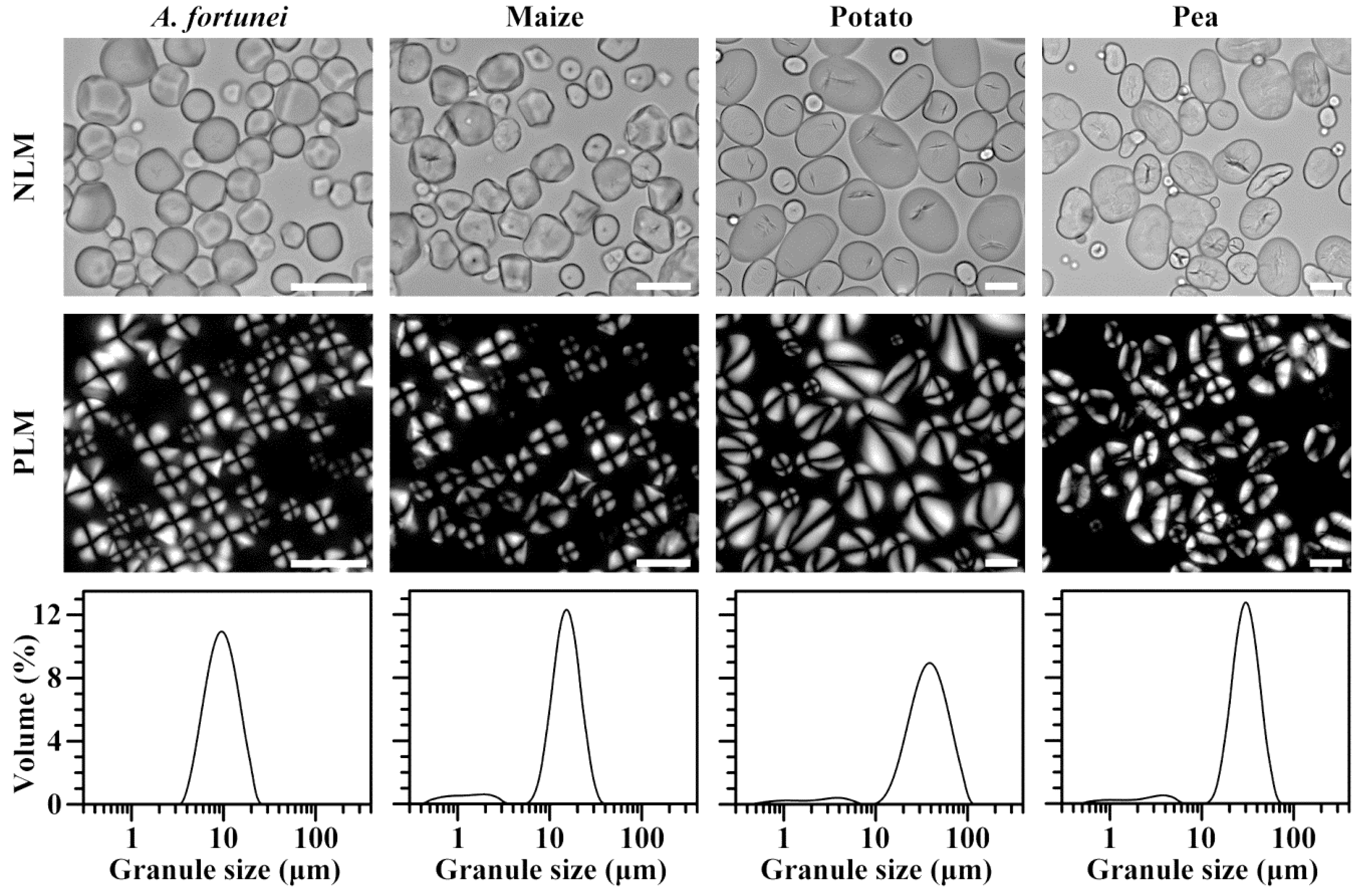
2.2. Morphology and Granule Sizes of Starch
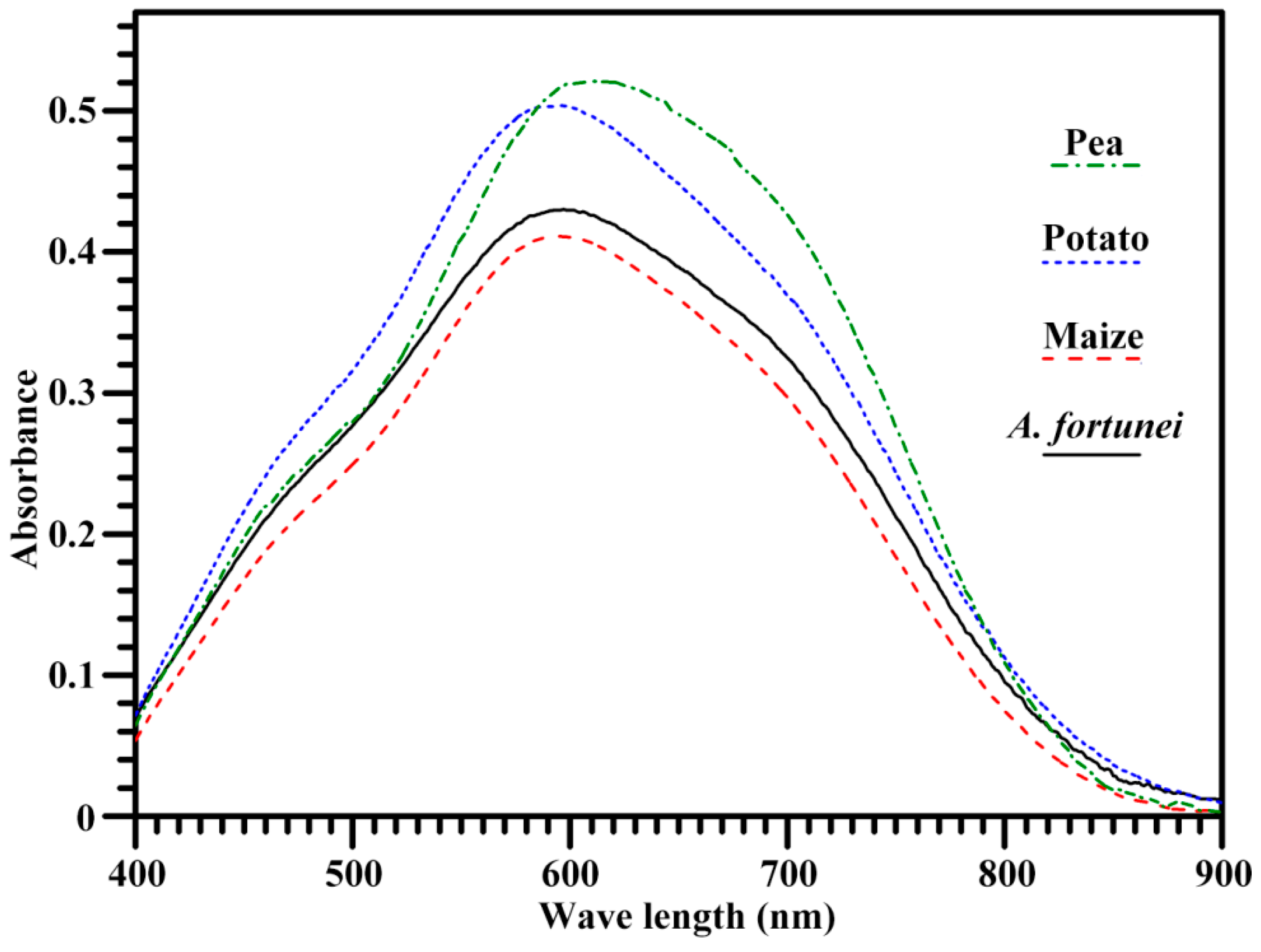
2.3. Iodine Absorption Spectrum and Apparent Amylose Content of Starch
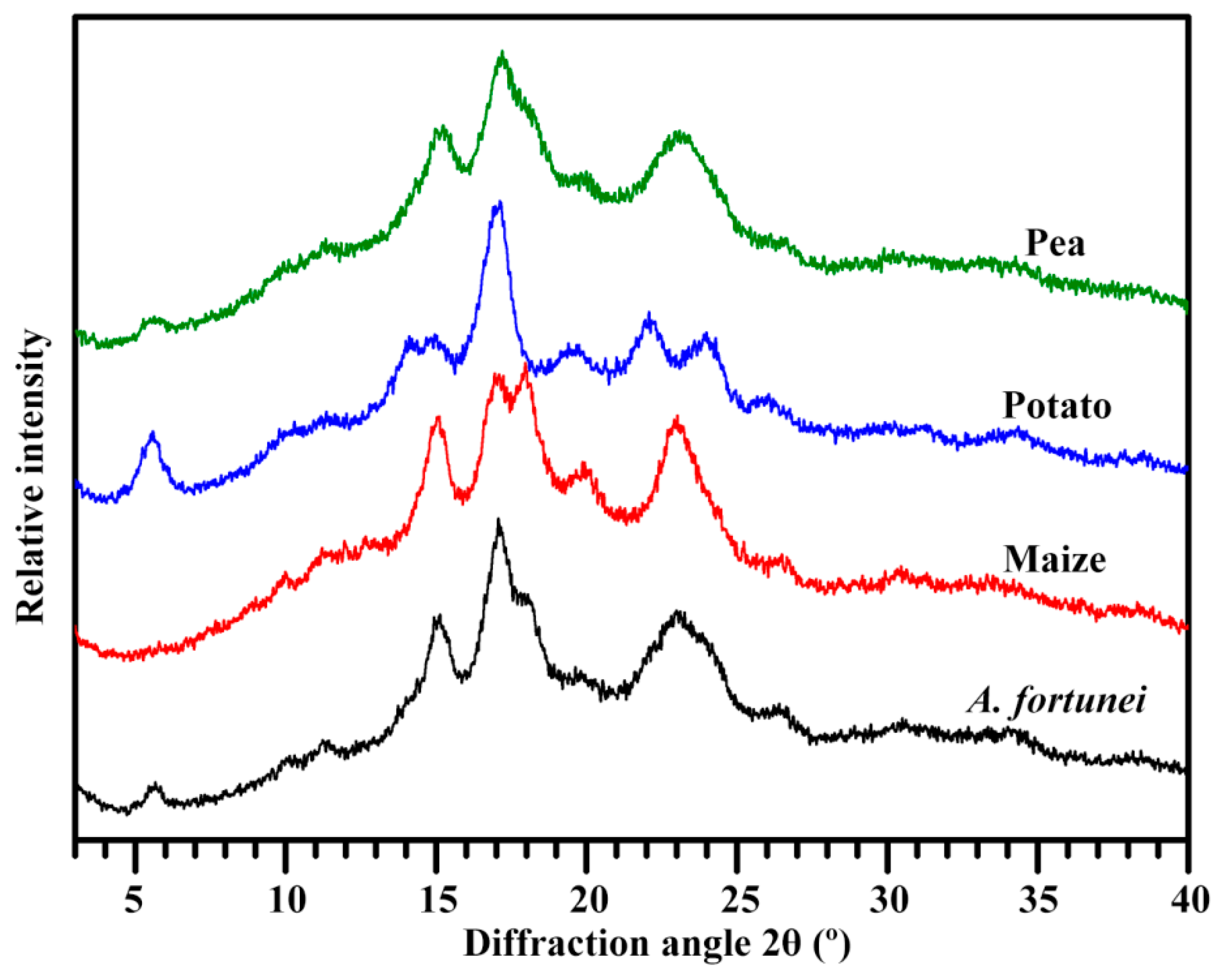
2.4. Crystalline Structure of Starch
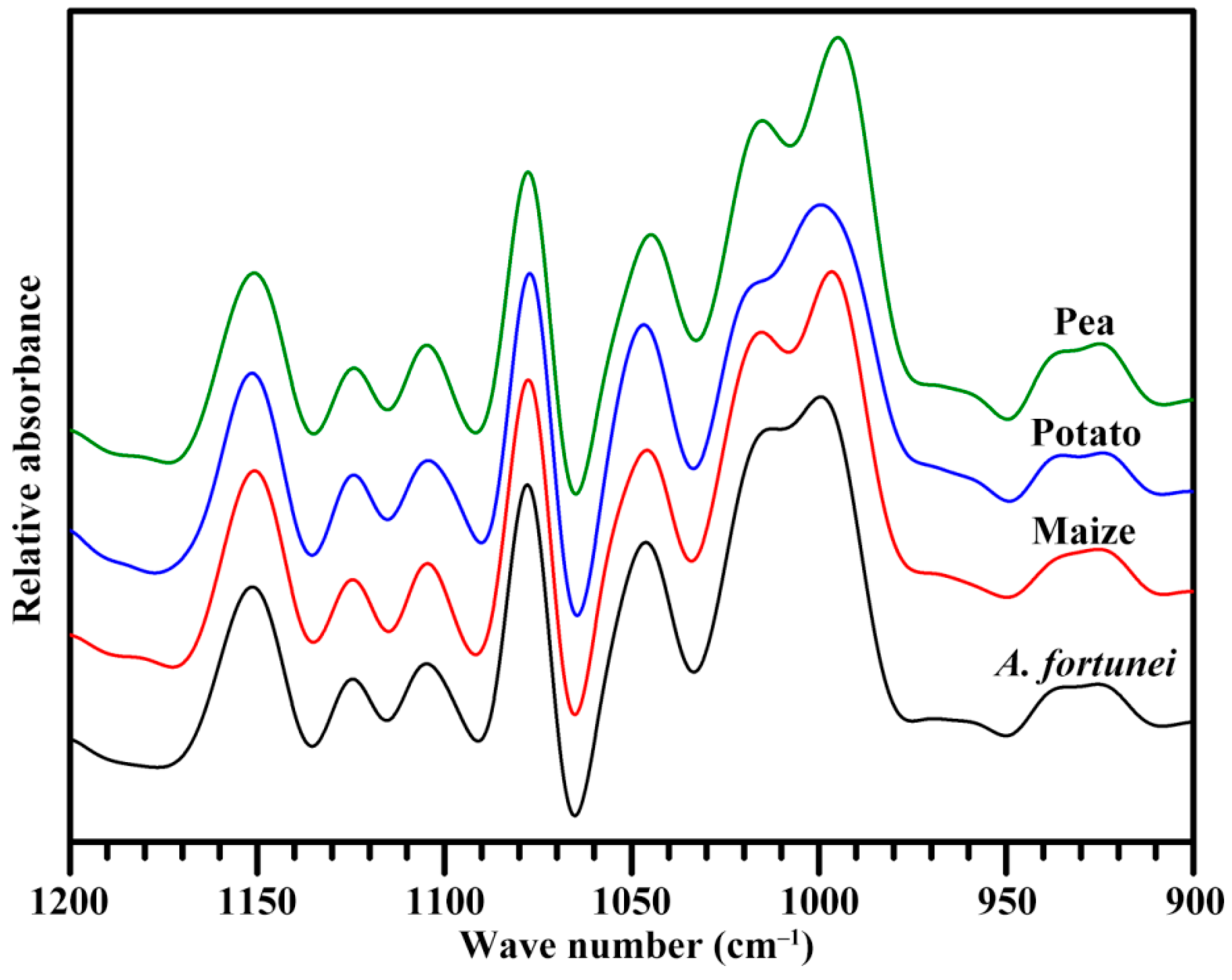
2.5. Short-Range Ordered Structure of Starch
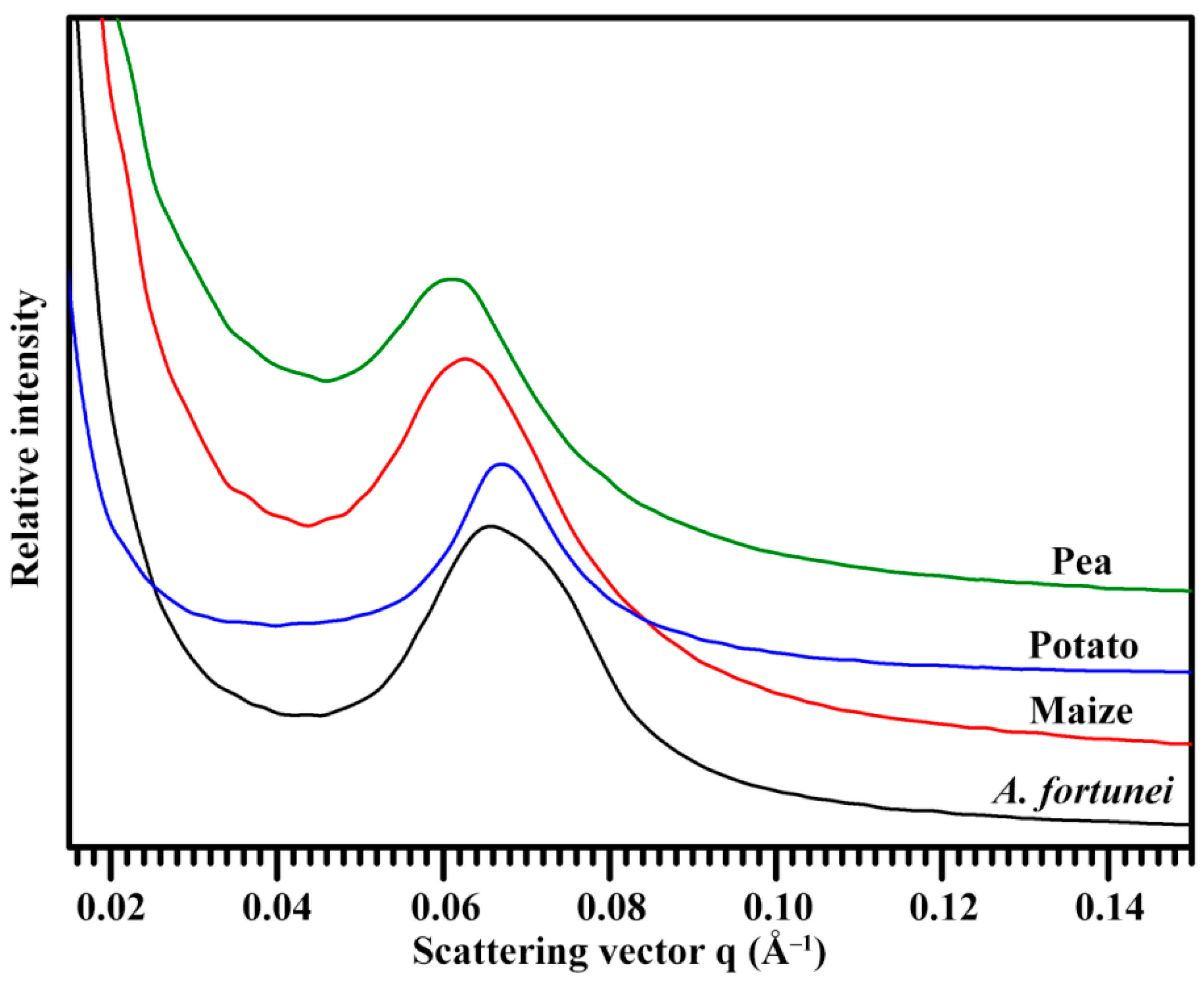
2.6. Lamellar Structure of Starch
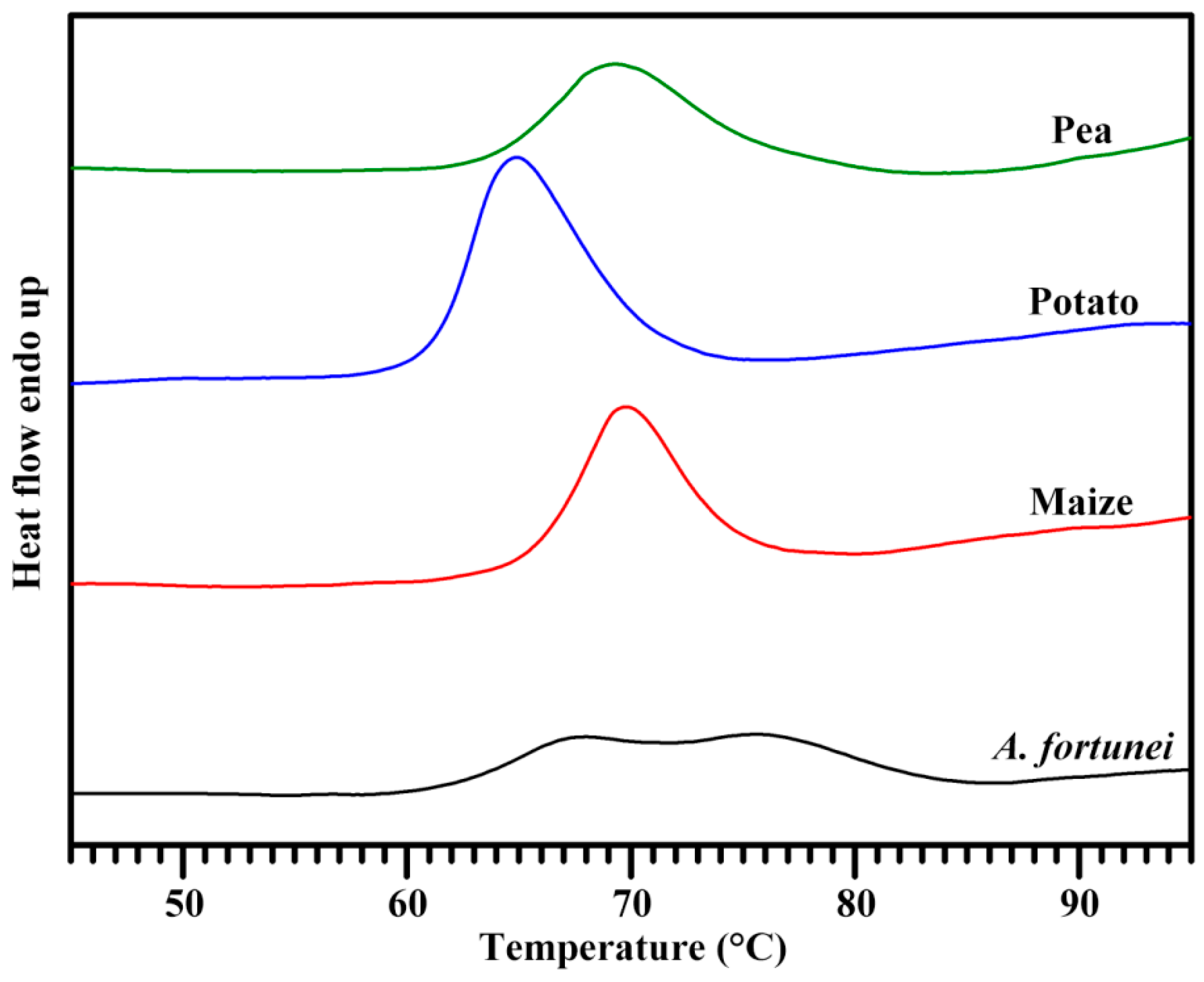
2.7. Thermal Properties of Starch
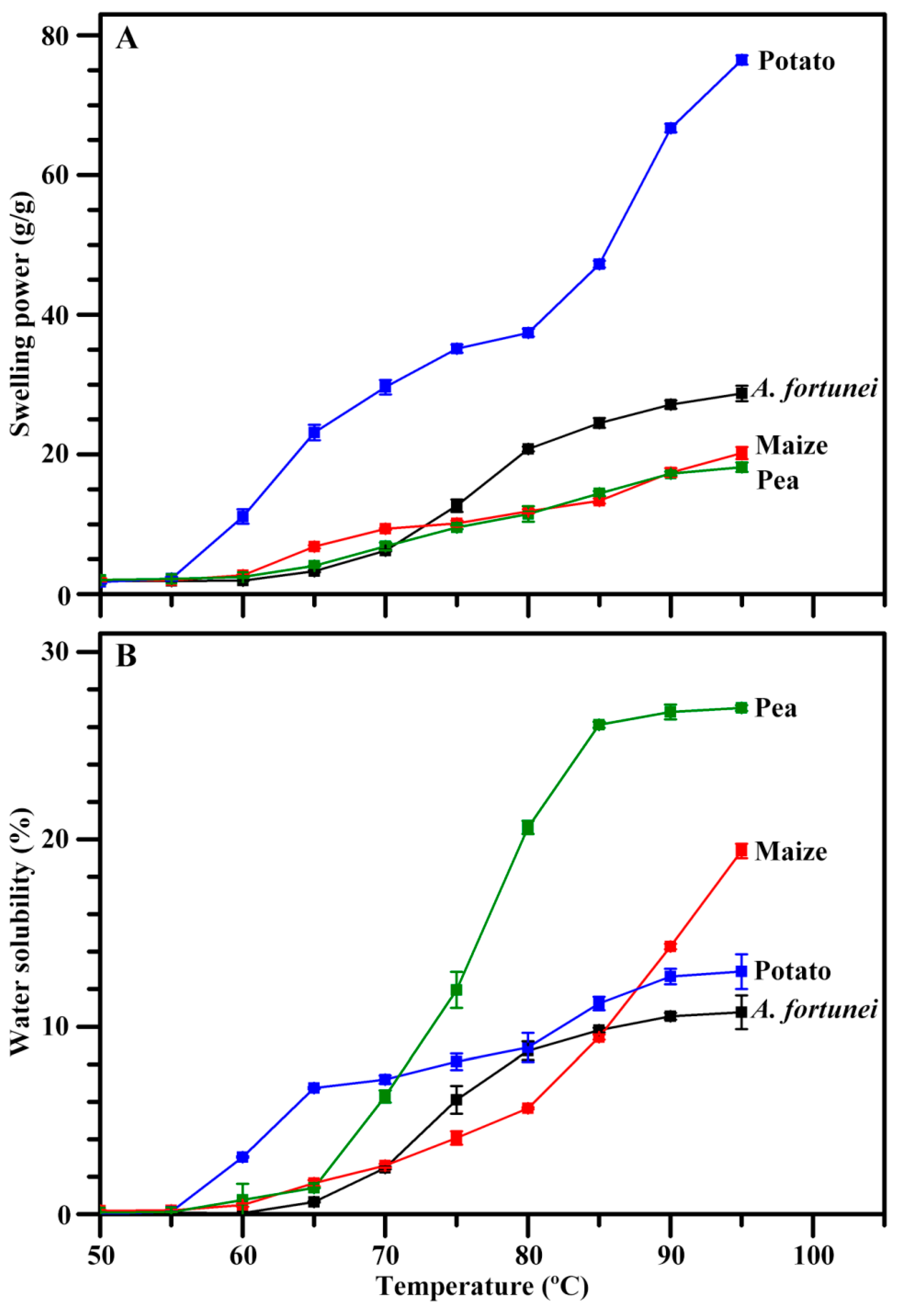
2.8. Swelling Power and Water Solubility of Starch
2.9. Pasting Properties of Starch
2.10. Digestion Properties of Starch
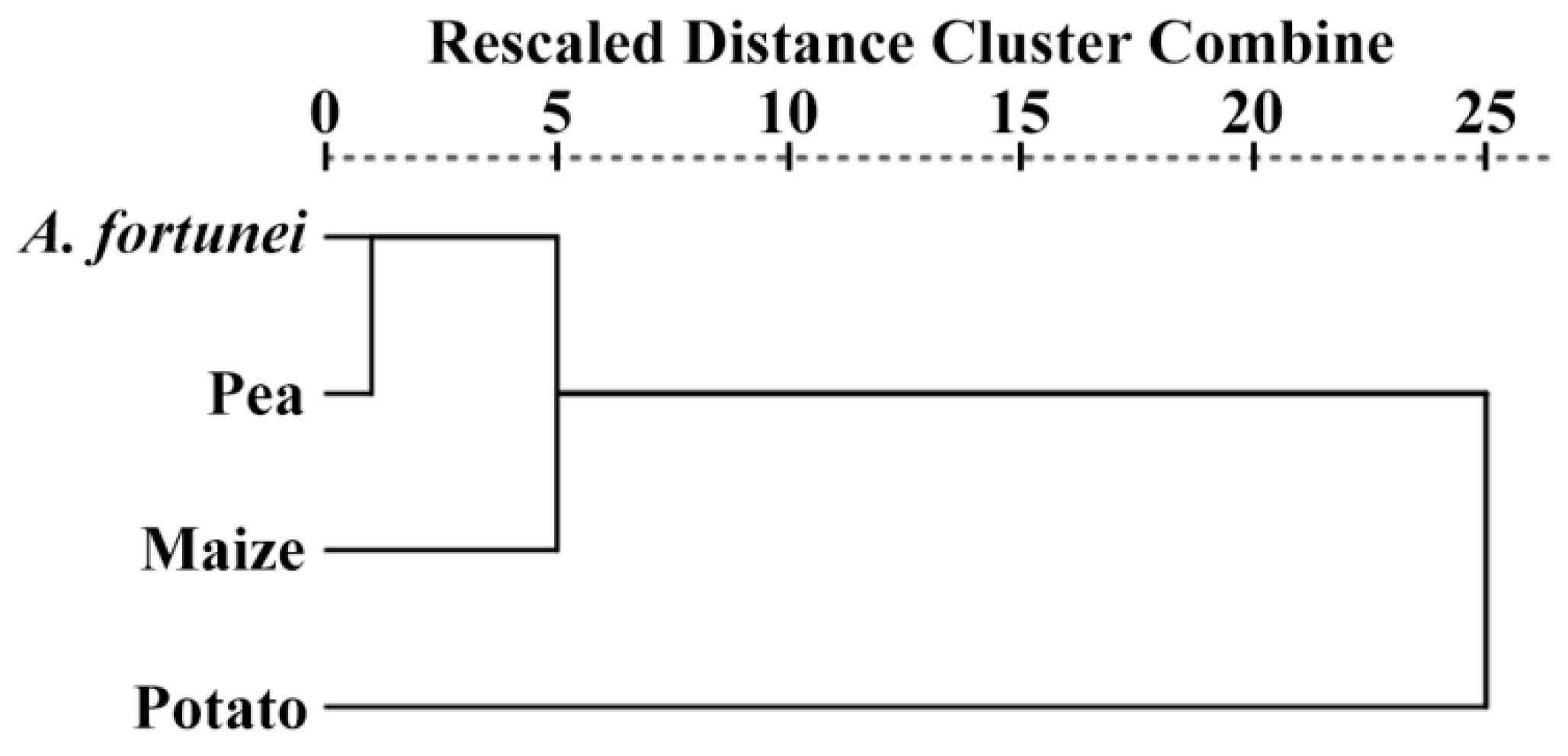
2.11. Cluster Analysis of Starch
3. Materials and Methods
3.1. Plant. Materials
3.2. Determination of Starch and Soluble Sugar Content in Root Tuber
3.3. Starch Isolation
3.4. Morphology Observation and Granule Size Analysis
3.5. Measurements of Iodine Absorption Spectrum and AAC
3.6. Crystalline Structure Analysis
3.7. Short-Range Ordered Structure Analysis
3.8. Lamellar Structure Analysis
3.9. Measurement of Swelling Power and Water Solubility
3.10. Measurement of Thermal Properties
3.11. Measurement of Pasting Properties
3.12. Measurement of Digestion Properties
3.13. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Cai, J.; Cai, C.; Man, J.; Zhou, W.; Wei, C. Structural and functional properties of C-type starches. Carbohydr. Polym. 2014, 101, 289–300. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Zhao, L.; Man, J.; Wang, J.; Zhou, W.; Huai, H.; Wei, C. Comparison of physicochemical properties of B-type nontraditional starches from different sources. Int. J. Biol. Macromol. 2015, 78, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Guo, K.; Lin, L.; Fan, X.; Zhang, L.; Wei, C. Comparison of structural and functional properties of starches from five fruit kernels. Food Chem. 2018, 257, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhao, L.; Bian, X.; Guo, K.; Zhou, L.; Wei, C. Characterization and comparative study of starches from seven purple sweet potatoes. Food Hydrocoll. 2018, 80, 168–176. [Google Scholar] [CrossRef]
- Hoover, R. Composition, molecular structure, and physicochemical properties of tuber and root starches: A review. Carbohydr. Polym. 2001, 45, 253–267. [Google Scholar] [CrossRef]
- Hoover, R.; Hughes, T.; Chung, H.J.; Liu, Q. Composition, molecular structure, properties, and modification of pulse starches: A review. Food Res. Int. 2010, 43, 399–413. [Google Scholar] [CrossRef]
- Lin, L.; Guo, D.; Zhao, L.; Zhang, X.; Wang, J.; Zhang, F.; Wei, C. Comparative structure of starches from high-amylose maize inbred lines and their hybrids. Food Hydrocoll. 2016, 52, 19–28. [Google Scholar] [CrossRef]
- Moorthy, S.N. Physicochemical and functional properties of tropical tuber starches: A review. Starch 2002, 54, 559–592. [Google Scholar] [CrossRef]
- Wani, I.A.; Sogi, D.S.; Hamdani, A.M.; Gani, A.; Bhat, N.A.; Shah, A. Isolation, composition, and physicochemical properties of starch from legumes: A review. Starch 2016, 68, 1–12. [Google Scholar] [CrossRef]
- Fan, X.; Zhang, S.; Lin, L.; Zhao, L.; Liu, A.; Wei, C. Properties of new starches from tubers of Arisaema elephas, yunnanense and erubescens. Food Hydrocoll. 2016, 61, 183–190. [Google Scholar] [CrossRef]
- Gao, H.; Cai, J.; Han, W.; Huai, H.; Chen, Y.; Wei, C. Comparison of starches isolated from three different Trapa species. Food Hydrocoll. 2014, 37, 174–181. [Google Scholar] [CrossRef]
- Huang, J.; Zhao, L.; Huai, H.; Li, E.; Zhang, F.; Wei, C. Structural and functional properties of starches from wild Trapa quadrispinosa, japonica, mammillifera and incise. Food Hydrocoll. 2015, 48, 117–126. [Google Scholar] [CrossRef]
- Blazek, J.; Gilbert, E.P. Application of small-angle X-ray and neutron scattering techniques to the characterisation of starch structure: A review. Carbohydr. Polym. 2011, 85, 281–293. [Google Scholar] [CrossRef]
- Cheetham, N.W.H.; Tao, L. Variation in crystalline type with amylose content in maize starch granules: An X-ray powder diffraction study. Carbohydr. Polym. 1998, 36, 277–284. [Google Scholar] [CrossRef]
- He, W.; Wei, C. Progress in C-type starches from different plant sources. Food Hydrocoll. 2017, 73, 162–175. [Google Scholar] [CrossRef]
- Bogracheva, T.Y.; Morris, V.J.; Ring, S.G.; Hedley, C.L. The granular structure of C-type pea starch and its role in gelatinization. Biopolym. 1998, 45, 323–332. [Google Scholar] [CrossRef]
- Buléon, A.; Gerard, C.; Riekel, C.; Vuong, R.; Chanzy, H. Details of the crystalline ultrastructure of C-starch granules revealed by synchrotron microfocus mapping. Macromolecules 1998, 31, 6605–6610. [Google Scholar] [CrossRef]
- Wei, C.; Qin, F.; Zhou, W.; Yu, H.; Xu, B.; Chen, C.; Zhu, L.; Wang, Y.; Gu, M.; Liu, Q. Granule structure and distribution of allomorphs in C-type high-amylose rice starch granule modified by antisense RNA inhibition of starch branching enzyme. J. Agric. Food Chem. 2010, 58, 11946–11954. [Google Scholar] [CrossRef] [PubMed]
- Cai, C.; Cai, J.; Man, J.; Yang, Y.; Wang, Z.; Wei, C. Allomorph distribution and granule structure of lotus rhizome C-type starch during gelatinization. Food Chem. 2014, 142, 408–415. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Cai, C.; Man, J.; Yang, Y.; Zhang, F.; Wei, C. Crystalline and structural properties of acid-modified lotus rhizome C-type starch. Carbohydr. Polym. 2014, 102, 799–807. [Google Scholar] [CrossRef] [PubMed]
- Cai, C.; Lin, L.; Man, J.; Zhao, L.; Wang, Z.; Wei, C. Different structural properties of high-amylose maize starch fractions varying in granule size. J. Agric. Food Chem. 2014, 62, 11711–11721. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Yu, J.; Zhu, Q.; Yu, J.; Jin, F. Granular structure and allomorph position in C-type Chinese yam starch granule revealed by SEM, 13C CP/MAS NMR and XRD. Food Hydrocoll. 2009, 23, 426–433. [Google Scholar] [CrossRef]
- Fan, X.; Zhao, L.; Zhang, L.; Xu, B.; Wei, C. A new allomorph distribution of C-type starch root tuber of Apios fortunei. Food Hydrocoll. 2017, 66, 334–342. [Google Scholar] [CrossRef]
- Li, N.; Zhang, S.; Zhao, Y.; Li, B.; Zhang, J. Over-expression of AGPase genes enhances seed weight and starch content in transgenic maize. Planta 2011, 233, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Ogasawara, Y.; Hidano, Y.; Kato, Y. Study on carbohydrate composition of Apios (Apios americana Medikus) flowers and tubers. J. Jpn. Soc. Food Sci. Technol. 2006, 53, 40–46. [Google Scholar] [CrossRef]
- Zhang, S.; Fan, X.; Lin, L.; Zhao, L.; Liu, A.; Wei, C. Properties of starch from root tuber of Stephania epigaea in comparison with potato and maize starches. Int. J. Food Prop. 2017, 20, 1740–1750. [Google Scholar] [CrossRef]
- Sandhu, K.S.; Singh, N.; Kaur, M. Characteristics of the different corn types and their grain fractions: Physicochemical, thermal, morphological and rheological properties of starches. J. Food Eng. 2004, 64, 119–127. [Google Scholar] [CrossRef]
- Lin, L.; Zhang, Q.; Zhang, L.; Wei, C. Evaluation of the molecular structural parameters of normal rice starch and their relationships with its thermal and digestion properties. Molecules 2017, 22, 1526. [Google Scholar] [CrossRef] [PubMed]
- Yangcheng, H.; Belamkar, V.; Cannon, S.B.; Jane, J.L. Characterization and development mechanism of Apios americana tuber starch. Carbohydr. Polym. 2016, 151, 198–205. [Google Scholar] [CrossRef] [PubMed]
- Genkina, N.K.; Takahiro, N.; Koltisheva, G.I.; Wasserman, L.A.; Tester, R.F.; Yuryev, V.P. Effects of growth temperature on some structural properties of crystalline lamellae in starches extracted from sweet potatoes (Sunnyred and Ayamurasaki). Starch 2003, 55, 350–357. [Google Scholar] [CrossRef]
- Sevenou, O.; Hill, S.E.; Farhat, I.A.; Mitchell, J.R. Organisation of the external region of the starch granule as determined by infrared spectroscopy. Int. J. Biol. Macromol. 2002, 31, 79–85. [Google Scholar] [CrossRef]
- Cai, J.; Man, J.; Huang, J.; Liu, Q.; Wei, W.; Wei, C. Relationship between structure and functional properties of normal rice starches with different amylose contents. Carbohydr. Polym. 2015, 124, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Sanderson, J.S.; Daniels, R.D.; Donald, A.M.; Blennow, A.; Engelsen, S.B. Exploratory SAXS and HPAEC-PAD studies of starches from diverse plant genotypes. Carbohydr. Polym. 2006, 64, 433–443. [Google Scholar] [CrossRef]
- Yuryev, V.P.; Krivandin, A.V.; Kiseleva, V.I.; Wasserman, L.A.; Genkina, N.K.; Fornal, J.; Blaszczak, W.; Schiraldi, A. Structural parameters of amylopectin clusters and semi-crystalline growth rings in wheat starches with different amylose content. Carbohydr. Res. 2004, 339, 2683–2691. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Fan, X.; Wang, Z.; Wei, C. Application of quantitative graphical method based on small angle X-ray scattering spectrum in crop starch study. Acta Agron. Sin. 2017, 43, 1827–1834. [Google Scholar] [CrossRef]
- Abegunde, O.K.; Mu, T.H.; Chen, J.W.; Deng, F.M. Physicochemical characterization of sweet potato starches popularly used in Chinese starch industry. Food Hydrocoll. 2013, 33, 169–177. [Google Scholar] [CrossRef]
- Singh, N.; Kaur, L.; Ezekiel, R.; Guraya, H.S. Microstructural, cooking and textural characteristics of potato (Solanum tuberosum L.) tubers in relation to physicochemical and functional properties of their flours. J. Sci. Food Agric. 2005, 85, 1275–1284. [Google Scholar] [CrossRef]
- Englyst, H.N.; Kingman, S.M.; Cummings, J.H. Classification and measurement of nutritionally important starch fractions. Eur. J. Clin. Nutr. 1992, 45, S33–S50. [Google Scholar]
- Wang, S.; Copeland, L. Molecular disassembly of starch granules during gelatinization and its effect on starch digestibility: A review. Food Funct. 2013, 4, 1564–1580. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.J.; Lim, H.S.; Lim, S.T. Effect of partial gelatinization and retrogradation on the enzymatic digestion of waxy rice starch. J. Cereal Sci. 2006, 43, 353–359. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are available from the authors. |









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Starches | d(0.1) (μm) | d(0.5) (μm) | d(0.9) (μm) | D[4,3] (μm) |
|---|---|---|---|---|
| A. fortunei | 5.403 ± 0.002 a | 8.947 ± 0.002 a | 14.760 ± 0.004 a | 9.584 ± 0.002 a |
| Maize | 7.519 ± 0.031 b | 13.767 ± 0.078 b | 21.305 ± 0.173 b | 13.895 ± 0.086 b |
| Potato | 16.317 ± 0.010 c | 34.420 ± 0.004 d | 62.406 ± 0.036 d | 36.778 ± 0.006 d |
| Pea | 16.255 ± 0.078 c | 27.518 ± 0.169 c | 41.917 ± 0.359 c | 27.922 ± 0.204 c |
| Starches | λmax (nm) | BV | AAC (%) | RC (%) |
|---|---|---|---|---|
| A. fortunei | 597.3 ± 0.6 b | 0.355 ± 0.007 b | 35.0 ± 0.6 b | 25.9 ± 0.1 c |
| Maize | 594.3 ± 0.6 a | 0.328 ± 0.003 a | 30.9 ± 0.3 a | 20.0 ± 0.9 b |
| Potato | 598.3 ± 0.6 b | 0.403 ± 0.008 c | 41.0 ± 0.4 c | 20.0 ± 0.6 b |
| Pea | 613.3 ± 0.6 c | 0.460 ± 0.006 d | 47.0 ± 0.3 d | 16.9 ± 0.1 a |
| Starches | Ordered Structure Parameters | Lamellar Structure Parameters | |||
|---|---|---|---|---|---|
| 1045/1022 cm−1 | 1022/995 cm−1 | Imax (Counts) | Smax (Å−1) | D (nm) | |
| A. fortunei | 0.648 ± 0.004 b | 0.883 ± 0.018 c | 215.3 ± 0.7 c | 0.066 ± 0.001 b | 9.60 ± 0.14 a |
| Maize | 0.623 ± 0.009 a | 0.837 ± 0.016 b | 217.7 ± 6.5 c | 0.063 ± 0.001 a | 10.05 ± 0.07 b |
| Potato | 0.816 ± 0.010 c | 0.763 ± 0.011 a | 170.1 ± 0.4 b | 0.067 ± 0.001 b | 9.40 ± 0.01 a |
| Pea | 0.623 ± 0.008 a | 0.781 ± 0.005 a | 154.6 ± 3.8 a | 0.061 ± 0.001 a | 10.30 ± 0.01 b |
| Starches | To (°C) | Tp1 (°C) | Tp2 (°C) | Tc (°C) | ΔT (°C) | ΔH (J/g) |
|---|---|---|---|---|---|---|
| A. fortunei | 61.8 ± 0.3 b | 68.0 ± 0.3 b | 75.6 ± 0.1 c | 83.9 ± 0.3 d | 22.1 ± 0.4 d | 10.2 ± 0.9 a |
| Maize | 65.8 ± 0.2 d | ND | 70.0 ± 0.1 b | 74.7 ± 0.1 b | 9.0 ± 0.1 a | 10.5 ± 0.4 a |
| Potato | 61.0 ± 0.1 a | 64.8 ± 0.1 a | ND | 70.7 ± 0.3 a | 9.6 ± 0.3 b | 14.9 ± 0.3 b |
| Pea | 63.6 ± 0.1 c | ND | 69.3 ± 0.1 a | 76.6 ± 0.2 c | 13.0 ± 0.2 c | 9.5 ± 0.6 a |
| Starches | PV (mPa s) | HV (mPa s) | BV (mPa s) | FV (mPa s) | SV (mPa s) | PTime (min) | PTemp (°C) |
|---|---|---|---|---|---|---|---|
| A. fortunei | 1689 ± 16 c | 1420 ± 29 c | 269 ± 15 b | 2103 ± 37 c | 683 ± 8 d | 5.04 ± 0.04 b | 80.80 ± 0.48 b |
| Maize | 954 ± 15 b | 770 ± 9 b | 184 ± 6 a | 887 ± 9 b | 117 ± 1 a | 5.73 ± 0.01 c | 91.88 ± 0.03 c |
| Potato | 7860 ± 30 d | 3285 ± 32 d | 4574 ± 50 c | 3682 ± 18 d | 396 ± 14 c | 3.89 ± 0.04 a | 68.67 ± 0.46 a |
| Pea | 580 ± 23 a | 438 ± 21 a | 141 ± 4 a | 795 ± 21 a | 357 ± 9 b | 7.00 ± 0.01 d | 94.85 ± 0.76 d |
| Starches | Components | A. fortunei | Maize | Potato | Pea |
|---|---|---|---|---|---|
| Native starch | RDS (%) | 6.04 ± 0.62 b | 6.45 ± 0.03 b | 2.27 ± 0.01 a | 6.20 ± 0.13 b |
| SDS (%) | 10.96 ± 0.41 b | 22.98 ± 0.65 d | 6.33 ± 0.23 a | 20.23 ± 0.29 c | |
| RS (%) | 83.00 ± 0.22 c | 70.56 ± 0.68 a | 91.40 ± 0.24 d | 73.57 ± 0.42 b | |
| Gelatinized starch | RDS (%) | 83.16 ± 1.33 a | 81.59 ± 0.57 a | 84.14 ± 0.62 a | 83.66 ± 1.23 a |
| SDS (%) | 15.23 ± 0.12 d | 7.97 ± 0.23 a | 8.49 ± 0.14 b | 14.55 ± 0.33 c | |
| RS (%) | 1.61 ± 1.28 a | 10.44 ± 0.44 c | 7.37 ± 0.73 b | 1.79 ± 1.41 a | |
| Retrograded starch | RDS (%) | 78.13 ± 3.90 a | 78.72 ± 0.72 a | 80.15 ± 0.45 a | 78.49 ± 1.26 a |
| SDS (%) | 17.88 ± 2.27 b | 7.99 ± 0.48 a | 8.29 ± 0.21 a | 17.80 ± 0.38 b | |
| RS (%) | 3.99 ± 1.64 a | 13.29 ± 0.26 b | 11.57 ± 0.27 b | 3.72 ± 0.96 a |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Guo, K.; Fan, X.; Feng, G.; Wei, C. Physicochemical Properties of C-Type Starch from Root Tuber of Apios fortunei in Comparison with Maize, Potato, and Pea Starches. Molecules 2018, 23, 2132. https://doi.org/10.3390/molecules23092132
Wang J, Guo K, Fan X, Feng G, Wei C. Physicochemical Properties of C-Type Starch from Root Tuber of Apios fortunei in Comparison with Maize, Potato, and Pea Starches. Molecules. 2018; 23(9):2132. https://doi.org/10.3390/molecules23092132
Chicago/Turabian StyleWang, Juan, Ke Guo, Xiaoxu Fan, Gongneng Feng, and Cunxu Wei. 2018. "Physicochemical Properties of C-Type Starch from Root Tuber of Apios fortunei in Comparison with Maize, Potato, and Pea Starches" Molecules 23, no. 9: 2132. https://doi.org/10.3390/molecules23092132
APA StyleWang, J., Guo, K., Fan, X., Feng, G., & Wei, C. (2018). Physicochemical Properties of C-Type Starch from Root Tuber of Apios fortunei in Comparison with Maize, Potato, and Pea Starches. Molecules, 23(9), 2132. https://doi.org/10.3390/molecules23092132