Plant Growth Modulates Metabolites and Biological Activities in Retama raetam (Forssk.) Webb




Abstract
:1. Introduction
2. Results and Discussion
2.1. Lipophilic Compounds
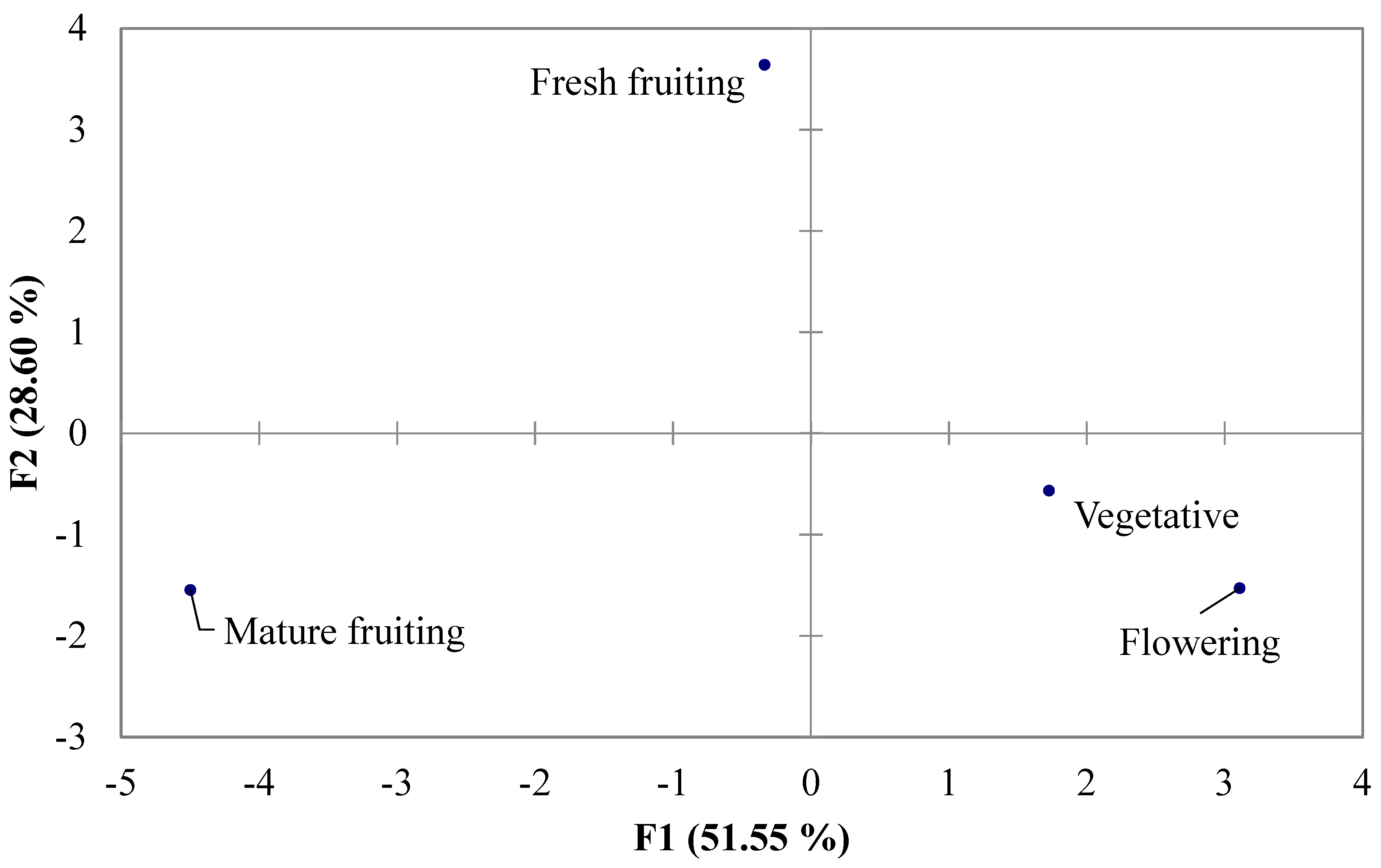
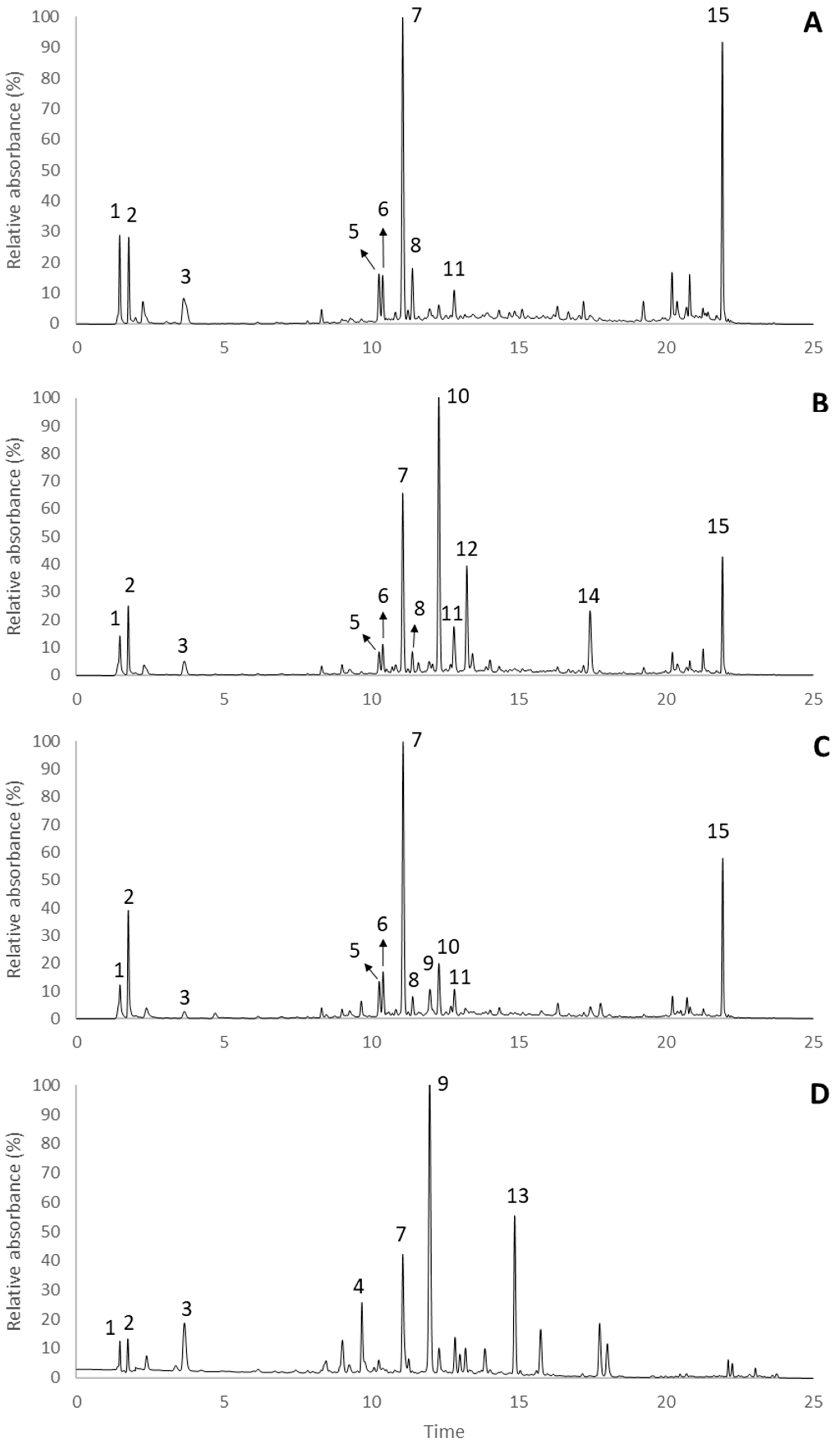
2.2. Hydrophylic Compounds
2.3. Antioxidant Activities
2.4. Antimicrobial Activity of Shoot Extracts
3. Materials and Methods
3.1. Plant Sampling and Extract Preparation
- 1-
- Vegetative stage: the plants, of about 2 m high, were collected in September 2010. The shrub had many long and velvety green twigs, jagged and covered with small silky white hairs (Figure 1a).
- 2-
- Flowering stage: the plants, of about 2 m high, were collected in February 2011. Their flowers (Figure 1b) were white, tiny and formed by 5 to 10 petals.
- 3-
- Fresh fruiting stage: the plants, of about 2 m high, were collected in April 2011. The green fruit (Figure 1c) was a small egg-shaped pod that ended with a beak.
- 4-
- Mature fruiting stage: the plants, of about 2 m high, were collected in collected in May 2011. The matured fruit (Figure 1d) contained only one kidney-shaped seed of a yellow ocher.
3.2. Chemical Reagents
3.3. Assessment of Lipophilic Compounds
3.3.1. Carotenoids
3.3.2. Fatty Acids
3.4. Assessment of Hydrophilic Compounds
3.4.1. Vitamin C
3.4.2. Proline
3.4.3. Phenolic Compounds
3.5. Evaluation of Antioxidant Activities
3.5.1. Total Antioxidant Capacity
3.5.2. DPPH• Scavenging Ability
3.5.3. ABTS•+ Scavenging Activity
3.5.4. Reducing Power
3.5.5. Ferrous Ion Chelating Activity
3.5.6. β-Carotene Bleaching Test
3.6. Evaluation of Antibacterial Activity
3.7. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Yanishlieva, N.V.; Marinova, E.; Pokorny, J. Natural antioxidants from herbs and spices. Eur. J. Lipid Sci. Technol. 2006, 108, 776–793. [Google Scholar] [CrossRef]
- Ângela, S.F.; Barros, F.L.; Carvalho, A.M.; Ferreira, I.C.F.R. Lipophilic and hydrophilic antioxidants, lipid peroxidation inhibition and radical scavenging activity of two Lamiaceae food plants. Eur. J. Lipid Sci. Technol. 2010, 112, 1115–1121. [Google Scholar]
- Hamana, N.; Longoa, E.; Schiraldib, A.; Scampicchioa, M. Radical scavenging activity of lipophilic antioxidants and extra-virgin olive oil by isothermal calorimetry. Thermochim. Acta 2017, 658, 1–6. [Google Scholar] [CrossRef]
- Correia, R.T.; Borges, K.C.; Medeiros, M.F.; Genovese, M.I. Bioactive compounds and phenolic-linked functionality of powdered tropical fruit residues. Food Sci. Technol. Int. 2012, 6, 539–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lü, J.M.; Lin, P.H.; Yao, Q.; Chen, C. Chemical and molecular mechanisms of antioxidants: Experimental approaches and model systems. J. Cell. Mol. Med. 2010, 14, 840–860. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.N.; JahanBristi, N.; Rafiquzzaman, M. Review on in vivo and in vitro methods evaluation of antioxidant activity. Saudi Pharm. J. 2013, 21, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Lisiewska, Z.; Kmiecik, W.; Korus, A. Content of vitamin C, carotenoids, chlorophylls and polyphenols in green parts of dill (Anethum graveolens L.) depending on plant height. J. Food Compos. Anal. 2006, 19, 134–140. [Google Scholar] [CrossRef]
- Liu, C.Z.; Guo, C.; Wang, Y.C.; Ouyang, F. Effect of light irradiation on hairy root growth and artemisinin biosynthesis of Artemisia annua. Process. Biochem. 2002, 38, 581–585. [Google Scholar] [CrossRef]
- Xie, B.D.; Wang, H.T. Effects of light spectrum and photoperiod on contents of flavonoid and terpene in leaves of Ginkgo biloba L. J. Nanjing Fore. Univ. Nat. Sci. Ed. 2006, 30, 51–54. [Google Scholar]
- Briskin, D.P.; Gawienowski, M.C. Differential effects of light and nitrogen on production of hypericins and leaf glands in Hypericum perforatum. Plant Physiol. Biochem. 2001, 39, 1075–1081. [Google Scholar] [CrossRef]
- Edziri, H.; Mastouri, M.; Mahjoub, M.A.; Mighri, Z.; Mahjoub, A.; Verschaeve, L. Antibacterial, Antifungal and Cytotoxic Activities of Two Flavonoids from Retama raetam Flowers. Molecules 2012, 17, 7284–7293. [Google Scholar] [CrossRef] [PubMed]
- Djeddi, S.; Karioti, A.; Yannakopoulou, E.; Papadopoulosn, K.; Chatter, R.; Skaltsa, H. Analgesic and Antioxidant Activities of Algerian Retama raetam (Forssk.) Webb & Berthel Extracts. Rec. Nat. Prod 2013, 7, 169–176. [Google Scholar]
- El Hamrouni, A. Conservation des zones humides littorales et des ecosystèmes côtiers du Cap-bon. Rapport de diagnostic des sites partie relative à la flore et la végétation. In Rapport De Diagnostic Des Sites, Partie Relative Aux Mammifères; Agence de Protection et d’Amenagement du Littoral: Tunisie, Tunisia, 2001; pp. 1–33. [Google Scholar]
- Maghrani, M. Acute diuretic effect of aqueous extract of Retama raetam in normal rats. J. Ethnopharmacol. 2005, 99, 31–35. [Google Scholar] [CrossRef] [PubMed]
- Awen, B.Z.; Unnithan, C.R.; Ravi, S.; Kermagy, A.; Sasikumar, J.M.; Khrbash, A.S.; Ekreem, W.L. Essential oils of Retama raetam from Libya: Chemical composition and antimicrobial activity. Nat. Prod. Res. 2011, 9, 927–933. [Google Scholar] [CrossRef] [PubMed]
- Edziri, H.; Mastouri, M.; Cherai, I.; Aouni, M. Chemical composition and antibacterial, antifungal and antioxidant activities of the flower oil of Retama raetam (Forssk.) Webb from Tunisia. Nat. Prod. Res. 2010, 9, 789–796. [Google Scholar] [CrossRef] [PubMed]
- Saada, M.; Falleh, H.; Jalleli, I.; Snoussi, M.; Ksouri, R. Phenolic profile, biological activities and fraction analysis of the medicinal halophyte Retama raetam. S. Afr. J. Bot. 2014, 94, 114–121. [Google Scholar]
- Howard, L.R.; Talcott, S.T.; Brenes, C.H.; Villalon, B. Changes in Phytochemical and Antioxidant Activity of Selected Pepper Cultivars (Capsicum Species) as influenced by maturity. J. Agric. Food Chem. 2000, 48, 1713−1720. [Google Scholar] [CrossRef]
- Hopkins, W.G.; Évrard, C.M. Physiologie Végétale; De Boeck Supérieur: Paris, France, 2003; pp. 1–532. [Google Scholar]
- Richard, D.; Kefi, K.; Barbe, U.; Bausero, P.; Visioli, F. Polyunsaturated fatty acids as antioxidants. Pharmacol. Res. 2008, 57, 451–455. [Google Scholar] [CrossRef] [PubMed]
- Nelson, M.M.; Phleger, C.F.; Nichols, P.D. Seasonal lipid composition in macroalgae of the northeastern Pacific Ocean. Bot. Mar. 2002, 45, 58–65. [Google Scholar] [CrossRef]
- Los, D.; Mironov, K.; Allakhverdiev, S. Regulatory role of membrane fluidity in gene expression and physiological functions. Photosynth. Res. 2013, 116, 489–509. [Google Scholar] [CrossRef] [PubMed]
- Gallie, D.R. L-Ascorbic Acid: A Multifunctional Molecule Supporting Plant Growth and Development. Scientifica 2012, 2013, 795964. [Google Scholar] [CrossRef] [PubMed]
- Smirnoff, N.; Wheeler, G.L. Ascorbic acid in plants: Biosynthesis and function. Crit. Rev. Biochem. Mol. Biol. 2000, 35, 291–314. [Google Scholar] [CrossRef] [PubMed]
- Gallie, D.R. The role of l-ascorbic acid recycling in responding to environmental stress and in promoting plant growth. J. Exp. Bot. 2013, 64, 433–443. [Google Scholar] [CrossRef] [PubMed]
- Mattioli, R.; Falasca, G.; Sabatini, S.; Altamura, M.M.; Costantino, P.; Trovato, M. The proline biosynthetic genes P5CS1 and P5CS2 play overlapping roles in Arabidopsis flower transition but not in embryo development. Physiol. Plant. 2009, 137, 72–85. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, S.; Funck, D.; Szabados, L.; Rentsch, D. Proline metabolism and transport in plant development. J. Amino Acids 2010, 39, 949–962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kale, V.S. Variable Rates of Primary and Secondary Metabolites during Different Seasons and Physiological Stages in Convolvulus, Datura and Withania. Asian J. Exp. Biol. Sci. 2010, 10, 50–53. [Google Scholar]
- Ksouri, R.; Megdiche, W.; Falleh, H.; Trabelsi, N.; Boulaaba, M.; Smaoui, A.; Abdelly, C. Influence of biological, environmental and technical factors on phenolic content and antioxidant activities of Tunisian halophytes. C. R. Biol. 2008, 331, 865–873. [Google Scholar] [CrossRef] [PubMed]
- Parr, A.J.; Bolwell, P.A.J. Phenols in the plant and in man: The potential for possible nutritional enhancement of the diet by modifying the phenols content or profile. J. Sci. Food Agric. 2000, 80, 985–1012. [Google Scholar] [CrossRef]
- Combris, P.; Amiot-Carlin, M.J.; Caillavet, F.; Causse, M.; Dallongeville, J.; Padilla, M.; Renard, C.; Soler, L.G. Les Fruits et Légumes dans l’Alimentation: Enjeux et Déterminants de la Consommation; Expertise Scientifique Collective INRA: Paris, France, 2007. [Google Scholar]
- Toor, R.K.; Savage, G.P.; Lister, C.E. Seasonal variations in the antioxidant compositions of greenhouse grown tomatoes. J. Food Compos. Anal. 2006, 19, 1–10. [Google Scholar] [CrossRef]
- Touati, R.; Santos, S.A.O.; Rocha, S.M.; Belhamel, K.; Silvestre, A.J.D. Phenolic composition and biological prospecting of grains and stems of Retama sphaerocarpa. Ind. Crops Prod. 2017, 95, 244–255. [Google Scholar] [CrossRef]
- León-González, A.J.; Navarro, I.; Acero, N.; Muñoz Mingarro, D.; Martín -Cordero, C. Genus Retama: A review on traditional uses, phytochemistry, and pharmacological activities. Phytochem. Rev. 2018, 17, 701–731. [Google Scholar] [CrossRef]
- González-Mauraza, N.H.; León-González, A.J.; Espartero, J.L.; Gallego-Fernández, J.B.; Sánchez-Hidalgo, M.; Martin-Cordero, C. Isolation and quantification of pinitol, a bioactive cyclitol, in Retama spp. Nat. Prod. Commun. 2016, 11, 405–406. [Google Scholar] [PubMed]
- Xu, W.H.; Adnan, J.A.R.; Muhammad, Y.; Mohammad, S.A.; Shabana, I.K.; Ikhlas, A.K. Two New Flavonoids from Retama raetam. Helv. Chim. Acta. 2015, 98, 561–568. [Google Scholar] [CrossRef]
- Meot-Duros, L.; Le Floch, G.; Magné, C. Radical scavenging, antioxidant and antimicrobial activities of halophytic species. J. Ethnopharmacol. 2008, 116, 258–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scalbert, A.; Johnson, I.T.; Saltmarsh, M. Polyphenols: Antioxidants and beyond. Am. J. Clin. Nutr. 2005, 81, 215S–217S. [Google Scholar] [CrossRef] [PubMed]
- Bensaci, H.; Sekhri, L.; Atmani, A.; Tabchouche, A. A qualitative phytochemical analysis and a comparative study of the antibactrial activity of Retama stalks (raetam). Orient. J. Chem. 2016, 32, 313–319. [Google Scholar] [CrossRef]
- Hammouche-Mokranea, N.J.; León-Gonzálezb, A.; Navarroc, I.; Boulilaa, F.; Benallaouad, S.; Martín-Corderob, C. Phytochemical Profile and Antibacterial Activity of Retama raetam and R. sphaerocarpa cladodes from Algeria. Nat. Prod. Commun. 2017, 12, 1857–1860. [Google Scholar]
- Nonier, M.F.; De Gaulejac, N.V.; Vivas, N.; Vitry, C. Characterization of carotenoids and their degradation products in oak wood. Incidence on the flavour of wood. C. R. Chim. 2004, 689–698. [Google Scholar] [CrossRef]
- Megdiche Ksouri, W.; Chaouachi, F.; M’Rabet, R.; Medini, F.; Zaouali, Y.; Trabelsi, N.; Ksouri, R.; Noumi, E.; Abdelly, C. Antioxidant and antimicrobial properties of Frankenia thymifolia Desf. fractions and their related biomolecules identification by gas chromatography/mass spectrometry (GC/MS) and high performance liquid chromatography (HPLC). J. Med. Plants Res. 2011, 5, 5754–5765. [Google Scholar]
- Kampfenkel, K.; Van Montau, M.; Inze, D. Extraction and determination of ascorbate and dehydrascorbate from plant tissue. Anal. Biochem. 1995, 225, 165–167. [Google Scholar] [CrossRef] [PubMed]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of the free proline in water stress studies. Plant Soil 1973, 39, 205–208. [Google Scholar] [CrossRef]
- Dewanto, V.; Wu, X.; Adom, K.K.; Liu, R.H. Thermal processing enhances the nutritional value of tomatoes by increasing total antioxidant activity. J. Agric. Food Chem. 2002, 50, 3010–3014. [Google Scholar] [CrossRef] [PubMed]
- Catarino, M.D.; Silva, A.M.S.; Saraiva, S.C.; Sobral, A.J.F.N.; Cardoso, S.M. Characterization of phenolic constituents and evaluation of antioxidant properties of leaves and stems of Eriocephalus africanus. Arab. J. Chem. 2018, 11, 62–69. [Google Scholar] [CrossRef]
- Perestrelo, R.; Lu, Y.; Santos, S.A.O.; Silvestre, A.J.D.; Neto, C.P.; Câmara, J.S.; Rocha, S.M. Phenolic profile of Sercial and Tinta Negra Vitis vinifera L. grape skins by HPLC-DAD-ESI-MSn: Novel phenolic compounds in Vitis vinifera L. grape. Food Chem. 2012, 135, 94–104. [Google Scholar] [CrossRef] [Green Version]
- Määttä-Riihinen, K.R.; Kamal-Eldin, A.; Törrönen, A.R. Identification and quantification of phenolic compounds in berries of Fragaria and Rubus species (family rosaceae). J. Agric. Food Chem. 2004, 52, 6178–6187. [Google Scholar] [CrossRef] [PubMed]
- Falleh, H.; Ksouri, R.; Chaieb, K.; Karray-Bouraoui, N.; Trabelsi, N.; Boulaaba, M.; Chedly, A. Phenolic composition of Cynara cardunculus L. organs and their biological activities. C. R. Biol. 2008, 331, 372–379. [Google Scholar] [CrossRef] [PubMed]
- Prieto, P.; Pineda, M.; Aguilar, M. Spectrophotometric quantitation of antioxidant capacity through the formation of a phosphomolybdenum complex: Specific application to the determination of vitamin E. Anal. Biochem. 1999, 269, 337–341. [Google Scholar] [CrossRef] [PubMed]
- Hanato, T.; Kagawa, H.; Yasuhara, T.; Okuda, T. Two new flavonoids and other constituents in licorice root their relative astringency and radical scavenging effect. Chem. Pharm. Bull. 1988, 36, 1090–1097. [Google Scholar]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-evans, C. Antioxidant activity applying an improved ABTS•+ radical decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Oyaizu, M. Studies on products of browning reaction: Antioxidative activity of products of browning reaction. Jpn. J. Nutr. Diet. 1986, 44, 307–315. [Google Scholar] [CrossRef]
- Zhao, H.; Dong, J.; Lu, J.; Chen, J.; Li, Y.; Shan, L.; Lin, Y.; Fan, W.; Gu, G. Effect of extraction solvent mixtures on antioxidant activity evaluation and their extraction capacity and selectivity for free phenolic compounds in Barely (Hordeum vulgare L.). J. Agric. Food. Chem. 2006, 54, 7277–7286. [Google Scholar] [CrossRef] [PubMed]
- Koleva, I.I.; Teris, A.B.; Jozef, P.H.; Linssen, A.G.; Lyuba, N.E. Screening of plant extracts for antioxidant activity: A comparative study on three testing methods. Phytochem. Anal. 2002, 13, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Bagamboula, M.; Uyttendaele, J.; Debevere, M. Inhibitory effect of thyme and basil essential oils, carvacrol, thymol, estragol, linalool and p-cymene towards Shigella sonnei and S. flexneri. Food Microbiol. 2003, 21, 33–42. [Google Scholar] [CrossRef]
- Statistica. In Windows (Computer Program Electronic Manual); StatSoft, Inc.: Tulsa, OK, USA, 1998.
Sample Availability: Samples of the compounds are available from Mariem Saada and Susana M. Cardoso authors. |




{kind=link}
{kind=link}
{kind=link}
| Vegetative Stage | Flowering Stage | Fresh Fruiting Stage | Mature Fruiting Stage | |
|---|---|---|---|---|
| Carotenoids (μg·g−1 FW) | 16.67 ± 0.02 b | 9.72 ± 0.03 c | 2.67 ± 0.02 d | 27.19 ± 0.06 a |
| Fatty acids (relative %) | ||||
| Saturated | ||||
| Lauric acid C12:0 | 3.68 ± 0.18 a | 2.36 ± 0.12 b | - | 0.04 ± 0.00 c |
| Myristic acid C14:0 | 1.51 ± 0.07 a | 1.13 ± 0.05 a | - | 0.16 ± 0.01 b |
| Pentadecylic acid C15:0 | - | - | 25.38 ± 1.27 a | 0.08 ± 0.00 b |
| Palmitic acid C16:0 | 19.10 ± 0.95 b | 20.52 ± 1.02 a | 12.2 ± 0.61 c | 12.23 ± 0.61 c |
| Margaric acid C17:0 | 0.52 ± 0.02 b | - | - | 1.28 ± 0.06 a |
| Stearic acid C18:0 | 3.66 ± 0.18 b | 4.63 ± 0.23 a | 2.39 ± 0.12 c | 4.14 ± 0.11 a |
| Arachidic acid C20:0 | - | 6.10 ± 0.30 a | - | 0.5 ± 0.21 b |
| Heneicosanoic acid C21:0 | - | 1 ± 0.05 | - | - |
| Behenic acid C22:0 | 2.24 ± 0.11 a | - | - | 0.18 ± 0.01 b |
| Unsaturated | ||||
| Oleic acid C18 :1 | - | - | - | 24.97 ± 1.25 |
| Linoleic acid C18:2 | 22.72 ± 1.13 b | 18.80 ± 0.94 c | 18.75 ± 0.94 c | 49.85 ± 2.49 a |
| α-linolenic acid C18:3 | 43.77 ± 2.18 b | 45.37 ± 2.26 a | 38.67 ± 1.93 c | 3.99 ± 0.19 d |
| SFA | 30.71 ± 1.53 c | 34.74 ±1.74 b | 39.97 ± 1.99 a | 18.61 ± 0.93 d |
| MUFA | - | - | - | 24 .97 ±1.25 |
| PUFA | 66.49 ± 3.32 a | 64.17 ± 3.02 b | 57.42 ± 2.87 c | 53.84 ± 2.69 d |
| Vegetative Stage | Flowering Stage | Fresh Fruiting Stage | Mature Fruiting Stage | |
|---|---|---|---|---|
| Vitamin C (mg·100g−1 FW) | ||||
| Total AA | 645.61 ± 0.15 a | 627 ± 0.18 b | 367 ± 0.17 c | 103.2 ± 0.00 d |
| Reduced AA | 173.2 ± 0.01 b | 184.5 ± 0.01 a | 130.6 ± 0.03 c | 101.2 ± 0.00 d |
| Oxidized AA | 472.45 ± 0.01 a | 442.51 ± 0.05 b | 236.27 ± 0.07 c | 2.01 ± 0.06 d |
| Proline (μmol·g−1 DW) | 12.1 ± 0.03 b | 25.4 ± 0.01 a | 11.23 ± 0.01 c | 1.56 ± 0.01 d |
| TPC (mg GAE·g−1 DW) | 23.93 ± 0.03 a | 20.75 ± 0.02 b | 18.23 ± 0.02 b | 15.17 ± 0.02 c |
| Peak | RT (min) | λmax (nm) | [M − H]− (m/z) | ESI-MSn Fragments | Proposed Compounds | Mean Content (μg/g Dry Plant Material) | |||
|---|---|---|---|---|---|---|---|---|---|
| Vegetative Stage | Flowering Stage | Fresh Fruiting Stage | Mature Fruiting Stage | ||||||
| 1 | 1.45 | 303 | 133 | MS2[133]: 115 | Malic acid | + | + | + | + |
| 2 | 1.74 | 227, 304 | 191 | MS2[191]: 111,173 | Citric acid | + | + | + | + |
| 3 | 3.63 | 223, 275 | 255 | MS2[255]: 165, 193, 179, 149 | Piscidic acid | + | + | + | + |
| 4 | 9.68 | 271, 334 | 593 | MS2[593]: 473, 503, 353, 383, 575 | Vicenin 2 | ND | ND | tc | + |
| 5 | 10.26 | 255 | 491* | MS2[491]: 283, 445; MS3[283]: 268 | Calycosin-O-hexoside | + | + | + | ND |
| 6 | 10. 39 | 261 | 563 | MS2[563]: 311, 283, 341, 269 | Genistein-C-hexoside-O-pentoside | + | + | + | ND |
| 7 | 11.07 | 261 | 431 | MS2[431]: 311, 269 | Genistein-8-C-hexoside | 888.53 ± 24.57 a | 571.74 ± 60.94 c | 628.19 ± 10.68 b | + |
| 8 | 11.93 | 262 | 461 | MS2[461]: 341, 371 | Tectorigenin-8-C-hexoside | + | + | + | ND |
| 9 | 11.98 | 289 | 303 | MS2[303]: 285, 177, 125; MS3[285]: 241, 175, 257, 199, 217 | Taxifolin | ND | ND | + | 224.92 ± 4.03 |
| 10 | 12.30 | 260, sh325 | 477* | MS2[477]: 269, 311 | Genistin | tc | 867.17 ± 44.21 | + | ND |
| 11 | 12.82 | 262, sh324 | 575 | MS2[575]: 431, 311, 341 | Genistein-3-hydroxy-3-methylglutaroyl | + | + | + | tc |
| 12 | 13.24 | 267, 336 | 431 | MS2[431]: 269 | Apigenin-7-O-glucoside | ND | + | ND | ND |
| 13 | 14.87 | 231, 307 | 805 | MS2[805]: 497, 351; MS3[497]: 145, 351, 333 | p-Coumaric acid derivative | ND | ND | ND | + |
| 14 | 17.43 | 261, sh331 | 269 | MS2[269]: 269, 225, 241 | Genistein | ND | + | ND | ND |
| 15 | 22.40 | 282 | 311 | MS2[311]: 249, 293 | Unkown | + | + | + | ND |
| Vegetative Stage | Flowering Stage | Fresh Fruiting Stage | Mature Fruiting Stage | |
|---|---|---|---|---|
| TAA (mg GAE·g−1 DW) | 55.6 ± 0.04 a | 32.3 ± 0.02 c | 26.02 ± 0.02 d | 43.73 ± 0.03 b |
| DPPH• (IC50 μg·mL−1) | 23 ± 0.01 a | 160 ± 0.01 d | 68 ± 0.00 c | 20.5 ± 0.00 b |
| ABTS•+ assay (IC50 μg·mL−1) | 380 ± 0.02 a | 940 ± 0.01 d | 780 ± 0.00 c | 540 ± 0.01 b |
| Reducing power (EC50 μg·mL−1) | 410 ± 0.00 a | 2500 ± 0.01 d | 2300 ± 0.01 c | 805 ± 0.00 b |
| Chelating power (EC50 μg·mL−1) | 1160 ± 0.01 a | 12,292 ± 0.03 d | 11,933 ± 0.00 c | 7600 ± 0.02 b |
| β-carotene bleaching (IC50 μg·mL−1) | 960 ± 0.01 a | 8500 ± 0.00 d | 7700 ± 0.00 c | 3100 ± 0.00 b |
| Bacterial Strains | Vegetative Stage | Flowering Stage | Fresh Fruiting Stage | Mature Fruiting Stage | |
|---|---|---|---|---|---|
| Gram-positive | |||||
| Bacillus cereus | ATCC 14579 | 12 ± 0.00 a | 12 ± 0.57 a | 9 ± 0.57 a | 10 ± 1.00 a |
| Staphylococcus aureus | ATCC 25923 | - | - | - | - |
| Staphylococcus epidermidis | CIP 106510 | - | - | - | - |
| Listeria monocytogenes | ATCC 19115 | 9 ± 0.57 d | 8 ± 0.57 c | - | 7 ± 0.00 d |
| Enterococcus faecalis | ATCC 29212 | 10 ± 0.57 c | 9 ± 1.15 b | 8 ± 0.00 b | - |
| Micrococcus luteus | NCIMB 8166 | 9 ± 0.57 d | 9 ± 0.57 b | 9 ± 0.57 a | 9 ± 0.57 b |
| Gram-negative | |||||
| Escherichia coli | ATCC 85218 | - | - | - | - |
| Pseudomonas aeruginosa | ATCC 27853 | - | - | - | - |
| Salmonella typhimurium | ATCC 1408 | - | - | - | - |
| Aeromonas hydrophila | ATCC 7566 | 9 ± 0.00 d | 8 ± 0.57 c | 9 ± 0.57 a | 9 ± 0.00 b |
| Vibrio vulnificus | ATCC 27962T | 11 ± 1.15 b | 9 ± 0.57 b | - | - |
| Vibrio alginolyticus | ATCC 33787 | 9 ± 0.57 d | 9 ± 0.57 b | 8 ± 0.57 b | 8 ± 0.00 c |
| Vibrio cholerae | non-O1 IPT | 9 ± 0.00 d | 8 ± 1.00 c | - | - |
| Vibrio parahaemolyticus | ATCC 17802 | - | - | - | - |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saada, M.; Falleh, H.; Catarino, M.D.; Cardoso, S.M.; Ksouri, R. Plant Growth Modulates Metabolites and Biological Activities in Retama raetam (Forssk.) Webb. Molecules 2018, 23, 2177. https://doi.org/10.3390/molecules23092177
Saada M, Falleh H, Catarino MD, Cardoso SM, Ksouri R. Plant Growth Modulates Metabolites and Biological Activities in Retama raetam (Forssk.) Webb. Molecules. 2018; 23(9):2177. https://doi.org/10.3390/molecules23092177
Chicago/Turabian StyleSaada, Mariem, Hanen Falleh, Marcelo D. Catarino, Susana M. Cardoso, and Riadh Ksouri. 2018. "Plant Growth Modulates Metabolites and Biological Activities in Retama raetam (Forssk.) Webb" Molecules 23, no. 9: 2177. https://doi.org/10.3390/molecules23092177
APA StyleSaada, M., Falleh, H., Catarino, M. D., Cardoso, S. M., & Ksouri, R. (2018). Plant Growth Modulates Metabolites and Biological Activities in Retama raetam (Forssk.) Webb. Molecules, 23(9), 2177. https://doi.org/10.3390/molecules23092177