Effects of Germination on Protein, γ-Aminobutyric Acid, Phenolic Acids, and Antioxidant Capacity in Wheat

Abstract
:1. Introduction
2. Results and Discussion
2.1. Phenolic Acid Compositions of Germinated Wheat
2.2. γ-Aminobutyric Acid (GABA) Content of Germinated Wheat Grains
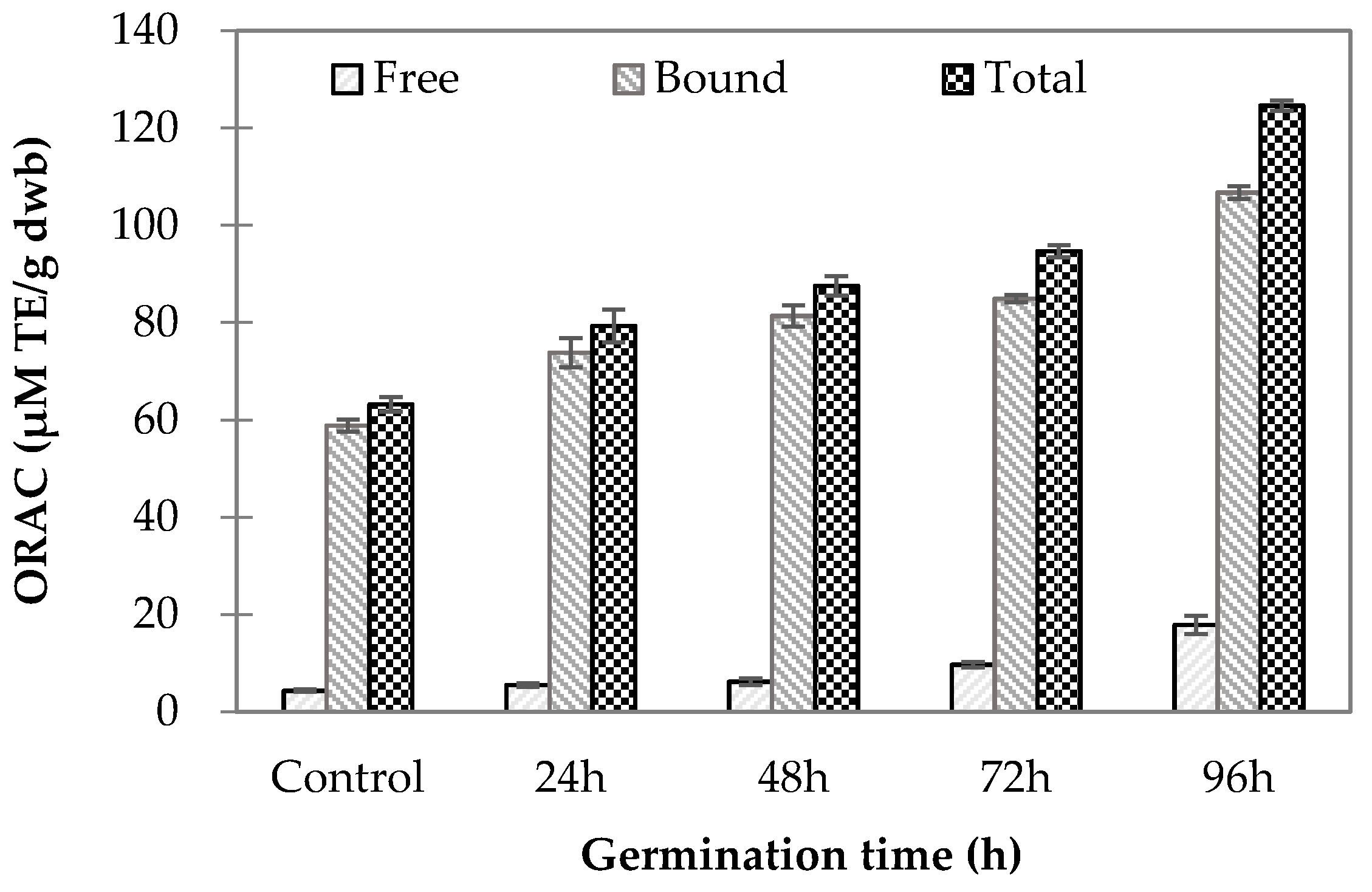
2.3. Oxygen Radical Absorbance Capacity (ORAC) of Germinated Wheat Grains
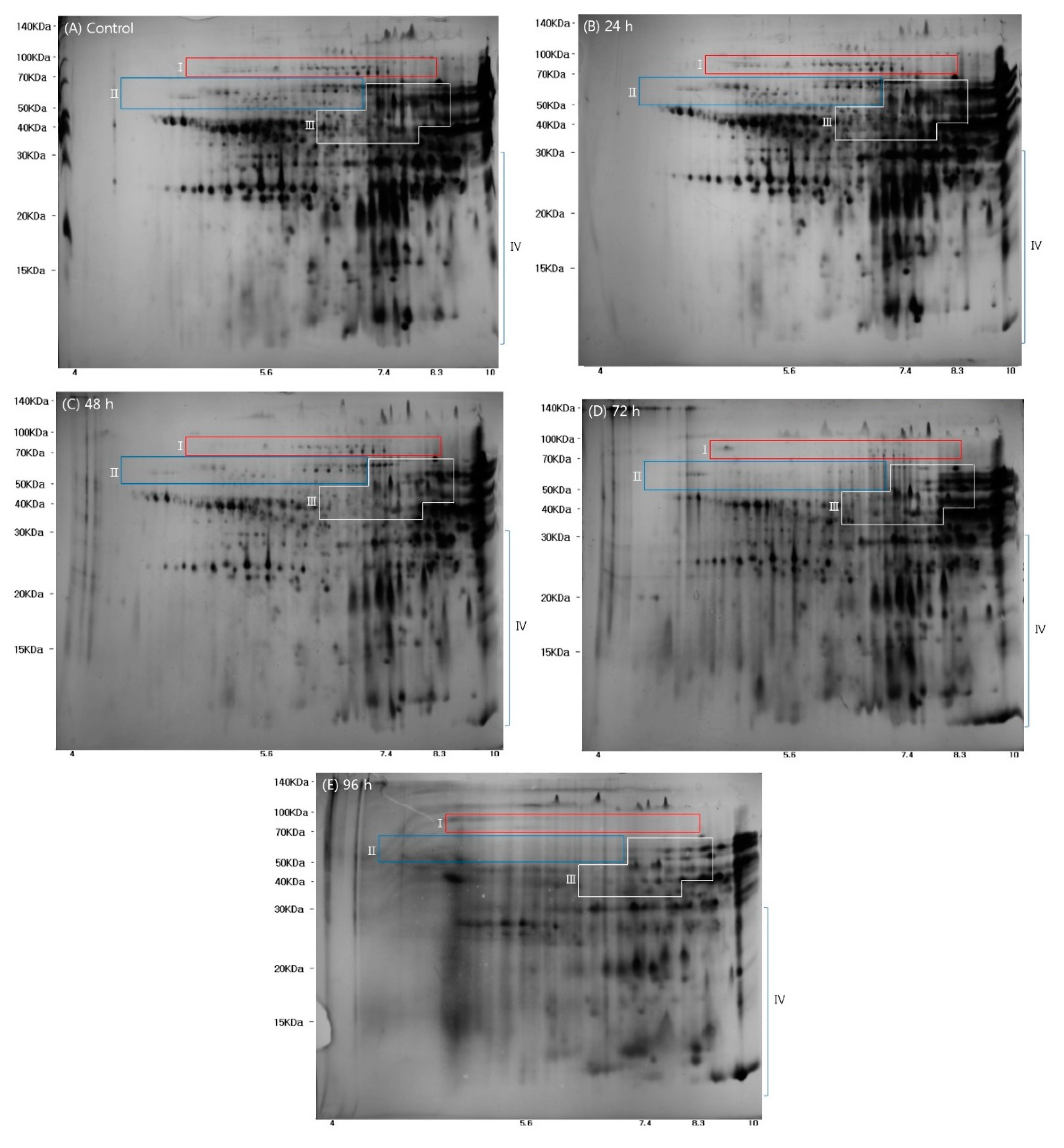
2.4. Wheat Grain 2-DE Protein Patterns during Germination
2.5. Correlation between Protein Spots and Antioxidant Properties in Germinated Wheat Samples
3. Materials and Methods
3.1. Materials
3.1.1. Wheat Sample and Germination Procedure
3.1.2. Chemicals and Reagents
3.2. Methods
3.2.1. Extraction of Free and Bound Phenolic Compounds
3.2.2. Oxygen Radical Absorbance Capacity (ORAC)
3.2.3. Phenolic Acid Compositions
3.2.4. Gamma-Aminobutyric Acid (GABA) Analysis
3.2.5. Two-Dimensional gel Electrophoresis (2-DE)
3.2.6. Peptide Mass Fingerprinting (PMF)
3.2.7. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Schaffer-Lequart, C.; Lehmann, U.; Ross, A.B.; Roger, O.; Eldridge, A.L.; Ananta, E.; Bietry, M.-F.; King, L.R.; Moroni, A.V.; Srichuwong, S. Whole grain in manufactured foods: Current use, challenges and the way forward. Crit. Rev. Food Sci. Nutr. 2017, 57, 1562–1568. [Google Scholar] [CrossRef] [PubMed]
- Van Hung, P.; Maeda, T.; Morita, N. Improvement of nutritional composition and antioxidant capacity of high-amylose wheat during germination. J. Food Sci. Technol. 2015, 52, 6756–6762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hung, P.V.; Maeda, T.; Yamamoto, S.; Morita, N. Effects of germination on nutritional composition of waxy wheat. J. Sci. Food Agric. 2012, 92, 667–672. [Google Scholar] [CrossRef] [PubMed]
- Ohm, J.B.; Lee, C.W.; Cho, K. Germinated wheat: Phytochemical composition and mixing characteristics. Cereal Chem. 2016, 93, 612–617. [Google Scholar] [CrossRef]
- Sharma, S.; Saxena, D.C.; Riar, C.S. Analysing the effect of germination on phenolics, dietary fibres, minerals and γ-amino butyric acid contents of barnyard millet (Echinochloa frumentaceae). Food Biosci. 2016, 13, 60–68. [Google Scholar] [CrossRef]
- Xu, L.; Wang, P.; Ali, B.; Yang, N.; Chen, Y.; Wu, F.; Xu, X. Changes of the phenolic compounds and antioxidant activities in germinated adlay seeds. J. Sci. Food Agric. 2017, 97, 4227–4234. [Google Scholar] [CrossRef] [PubMed]
- Nelson, K.; Stojanovska, L.; Vasiljevic, T.; Mathai, M. Germinated grains: A superior whole grain functional food? Can. J. Physiol. Pharmacol. 2013, 91, 429–441. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Yu, L.; Wang, X.; Gu, Z.; Beta, T. Changes of phenolic profiles and antioxidant activity in canaryseed (Phalaris canariensis L.) during germination. Food Chem. 2016, 194, 608–618. [Google Scholar] [CrossRef] [PubMed]
- Koehler, P.; Hartmann, G.; Wieser, H.; Rychlik, M. Changes of folates, dietary fiber, and proteins in wheat as affected by germination. J. Agric. Food Chem. 2007, 55, 4678–4683. [Google Scholar] [CrossRef] [PubMed]
- Donkor, O.N.; Stojanovska, L.; Ginn, P.; Ashton, J.; Vasiljevic, T. Germinated grains–sources of bioactive compounds. Food Chem. 2012, 135, 950–959. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Wang, P.; Weng, Y.; Ma, Y.; Gu, Z.; Yang, R. Comparison of phenolic profiles, antioxidant capacity and relevant enzyme activity of different chinese wheat varieties during germination. Food Biosci. 2017, 20, 159–167. [Google Scholar] [CrossRef]
- Van Hung, P.; Hatcher, D.W.; Barker, W. Phenolic acid composition of sprouted wheats by ultra-performance liquid chromatography (uplc) and their antioxidant activities. Food Chem. 2011, 126, 1896–1901. [Google Scholar] [CrossRef] [PubMed]
- Dong, K.; Zhen, S.M.; Cheng, Z.W.; Cao, H.; Ge, P.; Yan, Y.M. Proteomic analysis reveals key proteins and phosphoproteins upon seed germination of wheat (Triticum aestivum L.). Front. Plant Sci. 2015, 6, 14. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Zhu, C.; Dong, K.; Zhang, T.; Cheng, Z.; Li, J.; Yan, Y. Comparative proteome analysis of embryo and endosperm reveals central differential expression proteins involved in wheat seed germination. BMC Plant Biol. 2015, 15, 97. [Google Scholar] [CrossRef] [PubMed]
- Mandal, S.M.; Chakraborty, D.; Dey, S. Phenolic acids act as signaling molecules in plant-microbe symbioses. Plant Signal. Behav. 2010, 5, 359–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koo, S.C.; Kim, S.G.; Bae, D.-W.; Kim, H.Y.; Kim, H.T.; Lee, Y.H.; Kang, B.K.; Baek, S.-B.; Baek, I.Y.; Yun, H.T. Biochemical and proteomic analysis of soybean sprouts at different germination temperatures. J. Korean Soc. Appl. Biol. Chem. 2015, 58, 397–407. [Google Scholar] [CrossRef]
- Maksup, S.; Pongpakpian, S.; Roytrakul, S.; Cha-um, S.; Supaibulwatana, K. Comparative proteomics and protein profile related to phenolic compounds and antioxidant activity in germinated oryza sativa ‘kdml105’and thai brown rice ‘mali dang’for better nutritional value. J. Sci. Food Agric. 2017. [Google Scholar] [CrossRef] [PubMed]
- Hao, J.; Wu, T.; Li, H.; Wang, W.; Liu, H. Dual effects of slightly acidic electrolyzed water (saew) treatment on the accumulation of γ-aminobutyric acid (gaba) and rutin in germinated buckwheat. Food Chem. 2016, 201, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Baranzelli, J.; Kringel, D.H.; Colussi, R.; Paiva, F.F.; Aranha, B.C.; de Miranda, M.Z.; da Rosa Zavareze, E.; Dias, A.R.G. Changes in enzymatic activity, technological quality and gamma-aminobutyric acid (gaba) content of wheat flour as affected by germination. LWT-Food Sci. Technol. 2018, 90, 483–490. [Google Scholar] [CrossRef]
- Lamberts, L.; Joye, I.J.; Beliën, T.; Delcour, J.A. Dynamics of γ-aminobutyric acid in wheat flour bread making. Food Chem. 2012, 130, 896–901. [Google Scholar] [CrossRef]
- Sharma, S.; Saxena, D.C.; Riar, C.S. Changes in the gaba and polyphenols contents of foxtail millet on germination and their relationship with in vitro antioxidant activity. Food Chem. 2018, 245, 863–870. [Google Scholar] [CrossRef] [PubMed]
- Vensel, W.H.; Tanaka, C.K.; Altenbach, S.B. Protein composition of wheat gluten polymer fractions determined by quantitative two-dimensional gel electrophoresis and tandem mass spectrometry. Proteome Sci. 2014, 12, 8. [Google Scholar] [CrossRef] [PubMed]
- Hurkman, W.J.; Tanaka, C.K. Improved methods for separation of wheat endosperm proteins and analysis by two-dimensional gel electrophoresis. J. Cereal Sci. 2004, 40, 295–299. [Google Scholar] [CrossRef]
- Konopka, I.; Fornal, Ł.; Dziuba, M.; Czaplicki, S.; Nałȩcz, D. Composition of proteins in wheat grain streams obtained by sieve classification. J. Sci. Food Agric. 2007, 87, 2198–2206. [Google Scholar] [CrossRef]
- Mak, Y.; Willows, R.D.; Roberts, T.H.; Wrigley, C.W.; Sharp, P.J.; Copeland, L. Germination of wheat: A functional proteomics analysis of the embryo. Cereal Chem. J. 2009, 86, 281–289. [Google Scholar] [CrossRef]
- Okada, Y.; Iimure, T.; Takoi, K.; Kaneko, T.; Kihara, M.; Hayashi, K.; Ito, K.; Sato, K.; Takeda, K. The influence of barley malt protein modification on beer foam stability and their relationship to the barley dimeric α-amylase inhibitor-i (bdai-i) as a possible foam-promoting protein. J. Agric. Food Chem. 2008, 56, 1458–1464. [Google Scholar] [CrossRef] [PubMed]
- Bourne, L.C.; Rice-Evans, C. Bioavailability of ferulic acid. Biochem. Biophys. Res. Commun. 1998, 253, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Singhania, R.R.; Patel, A.K.; Sukumaran, R.K.; Larroche, C.; Pandey, A. Role and significance of beta-glucosidases in the hydrolysis of cellulose for bioethanol production. Bioresour. Technol. 2013, 127, 500–507. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Kim, S.S. Antioxidant and antiproliferative activities in immature and mature wheat kernels. Food Chem. 2016, 196, 638–645. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Kim, S.S. Utilisation of immature wheat flour as an alternative flour with antioxidant activity and consumer perception on its baked product. Food Chem. 2017, 232, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Oakley, B.R.; Kirsch, D.R.; Morris, N.R. A simplified ultrasensitive silver stain for detecting proteins in polyacrylamide gels. Anal. Biochem. 1980, 105, 361–363. [Google Scholar] [CrossRef]
- Fernandez, J.; Gharahdaghi, F.; Mische, S.M. Routine identification of proteins from sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-page) gels or polyvinyl difluoride membranes using matrix assisted laser desorption/ionization-time of flight-mass spectrometry (MALDI-TOF-MS). Electrophoresis 1998, 19, 1036–1045. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phenolic Acids | Germination Time (h) | |||||
|---|---|---|---|---|---|---|
| Control | 24 | 48 | 72 | 96 | ||
| Gallic acid (mg/100 g dwb 1) | Free *** | 0.18 ± 0.03 c | 0.25 ± 0.07 c | 0.31 ± 0.07 bc | 0.60 ± 0.23 b | 1.57 ± 0.29 a |
| Bound *** | 0.81 ± 0.09 c | 1.12 ± 0.12 b | 1.32 ± 0.09 b | 1.98 ± 0.20 a | 1.78 ± 0.21 a | |
| Total 2,*** | 0.99 ± 007 d | 1.36 ± 0.20 cd | 1.63 ± 0.02 c | 2.58 ± 0.35 b | 3.36 ± 0.35 a | |
| 4-Hydroxybenzoic acid (mg/100 g dwb 1) | Free *** | 0.19 ± 0.07 c | 0.38 ± 0.08 c | 0.54 ± 0.12 bc | 0.91 ± 0.36 b | 4.70 ± 0.36 a |
| Bound *** | 0.29 ± 0.03 c | 0.27 ± 0.07 c | 0.63 ± 0.08 b | 0.77 ± 0.11 ab | 0.85 ± 0.09 a | |
| Total 2,*** | 0.47 ± 0.04 d | 0.65 ± 0.15 cd | 1.17 ± 0.04 bc | 1.68 ± 0.45 b | 5.55 ± 0.45 a | |
| Vanillic acid (mg/100 g dwb 1) | Free *** | 0.16 ± 0.02 d | 0.32 ± 0.05 d | 0.77 ± 0.08 c | 2.79 ± 0.23 b | 3.72 ± 0.15 a |
| Bound *** | 1.51 ± 0.12 c | 1.71 ± 0.16 c | 1.64 ± 0.17 c | 16.27 ± 0.31 b | 19.43 ± 3.09 a | |
| Total 2,*** | 1.67 ± 0.13 c | 2.03 ± 0.18 c | 2.41 ± 0.25 c | 19.05 ± 0.26 b | 23.15 ± 3.24 a | |
| Caffeic acid (mg/100 g dwb 1) | Free *** | 0.82 ± 0.13 d | 14.20 ± 0.33 c | 13.84 ± 0.61 c | 15.85 ± 0.78 b | 17.61 ± 0.30 a |
| Bound *** | 0.11 ± 0.03 b | 0.09 ± 0.04 b | 0.10 ± 0.03 b | 0.07 ± 0.05 b | 0.38 ± 0.05 a | |
| Total 2,*** | 0.93 ± 0.11 d | 14.29 ± 0.35 c | 13.94 ± 0.58 c | 15.92 ± 0.77 b | 17.99 ± 0.29 a | |
| Syringic acid (mg/100 g dwb 1) | Free *** | 0.11 ± 0.07 b | 0.17 ± 0.04 b | 0.30 ± 0.09 b | 0.29 ± 0.06 b | 1.56 ± 0.20 a |
| Bound *** | 0.34 ± 0.06 c | 0.39 ± 0.03 c | 0.38 ± 0.02 c | 0.51 ± 0.03 b | 0.67 ± 0.07 a | |
| Total2,*** | 0.45 ± 0.13 c | 0.57 ± 0.05 c | 0.68 ± 0.10 bc | 0.80 ± 0.09 b | 2.22 ± 0.19 a | |
| Ferulic acid (mg/100 g dwb 1) | Free *** | 1.15 ± 0.17 c | 1.04 ± 0.11 c | 1.13 ± 0.08 c | 1.62 ± 0.13 b | 1.51 ± 0.09 a |
| Bound *** | 40.19 ± 1.70 d | 49.76 ± 4.46 c | 50.57 ± 1.22 b | 61.01 ± 2.56 b | 63.13 ± 0.77 a | |
| Total 2,*** | 41.35 ± 1.60 e | 50.81 ± 4.39 d | 51.69 ± 1.14 c | 62.63 ± 2.54 b | 64.64 ± 0.69 a | |
| p-Coumaric acid (mg/100 g dwb 1) | Free *** | 0.49 ± 0.06 b | 0.45 ± 0.06 b | 0.48 ± 0.04 b | 0.65 ± 0.05 a | 0.79 ± 0.09 a |
| Bound *** | 1.48 ± 0.05 c | 2.61 ± 0.21 b | 3.57 ± 0.26 b | 3.87 ± 0.17 a | 4.22 ± 0.09 a | |
| Total 2,*** | 1.97 ± 0.10 c | 3.06 ± 0.17 b | 4.06 ± 0.28 b | 4.52 ± 0.15 a | 5.01 ± 0.08 a | |
| GABA (mg/100 g dwb 1) | 4.55 ± 0.28 a | 12.93 ± 0.48 b | 7.82 ± 0.06 b | 4.51 ± 0.26 d | 39.98 ± 1.77 a | |
| Spot | Identified Proteins | Accession | Score 2 | Mw | PI | Relative Expression of Proteins | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| Control | 24 h | 48 h | 72 h | 96 h | ||||||
| 3104 | Cytosolic malate dehydrogenase | AAP7009.1 | 98 | 22 | 6.0 | 3313 | 3459 | 2553 | 2355 | 1601 |
| 4011 | Dimeric alpha-amylase inhibitor | ABI54569.1 | 88 | 14 | 6.6 | 215 | 176 | 87 | 77 | ND |
| 4101 | Hypothetical protein CL3131Contig 1 | AFG48690.1 | 69 | 24 | 6.3 | 339 | 204 | 188 | 191 | ND |
| 4010 | Alpha-amylase inhibitor | CAI84642.1 | 96 | 14 | 6.6 | 279 | 202 | 183 | 187 | ND |
| 4008 | Alpha-amylase/trypsin inhibitor | AQT26482.1 | 94 | 12 | 6.6 | 322 | 213 | 265 | 214 | ND |
| 5401 | Phospholipase SGR2isoform | XP_022027330.1 | 86 | 38 | 6.8 | 505 | 499 | 361 | 430 | 262 |
| 1603 | NI 1 | - | 56 | 5.3 | 139 | 153 | 122 | 85 | 85 | |
| 2813 | NI 1 | - | 88 | 5.6 | 753 | 1009 | 1926 | 1854 | 1854 | |
| 3708 | NI 1 | - | 63 | 5.7 | 191 | 192 | 99 | 142 | 537 | |
| 3405 | Granule bound starch synthase | AF75531.1 | 399 | 41 | 6.0 | 2189 | 2363 | 2362 | 2362 | 12,768 |
| 8009 | Dimeric alpha-amylase inhibitor | AAV39517.1 | 153 | 11 | 8.2 | 380 | 1823 | 1367 | 1780 | 2599 |
| 9004 | Glutathione S-transferase | XP_02255327.1 | 77 | 11 | 9.9 | 2581 | 5723 | 6173 | 6058 | 9776 |
| 4201 | unnamed protein product | CDJ26374.1 | 91 | 27 | 6.4 | 414 | 672 | 794 | 586 | 954 |
| 8013 | NI 1 | - | 11 | 8.9 | ND | 1939 | 2940 | 1884 | 3846 | |
| 1801 | Beta-glucosidase | XP_010066854.1 | 63 | 102 | 5.0 | 585 | 895 | 782 | 626 | 442 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, M.J.; Kwak, H.S.; Kim, S.S. Effects of Germination on Protein, γ-Aminobutyric Acid, Phenolic Acids, and Antioxidant Capacity in Wheat. Molecules 2018, 23, 2244. https://doi.org/10.3390/molecules23092244
Kim MJ, Kwak HS, Kim SS. Effects of Germination on Protein, γ-Aminobutyric Acid, Phenolic Acids, and Antioxidant Capacity in Wheat. Molecules. 2018; 23(9):2244. https://doi.org/10.3390/molecules23092244
Chicago/Turabian StyleKim, Mi Jeong, Han Sub Kwak, and Sang Sook Kim. 2018. "Effects of Germination on Protein, γ-Aminobutyric Acid, Phenolic Acids, and Antioxidant Capacity in Wheat" Molecules 23, no. 9: 2244. https://doi.org/10.3390/molecules23092244
APA StyleKim, M. J., Kwak, H. S., & Kim, S. S. (2018). Effects of Germination on Protein, γ-Aminobutyric Acid, Phenolic Acids, and Antioxidant Capacity in Wheat. Molecules, 23(9), 2244. https://doi.org/10.3390/molecules23092244