
Depsidone Derivatives and a Cyclopeptide Produced by Marine Fungus Aspergillus unguis under Chemical Induction and by Its Plasma Induced Mutant

 ,
,
Abstract
:1. Introduction
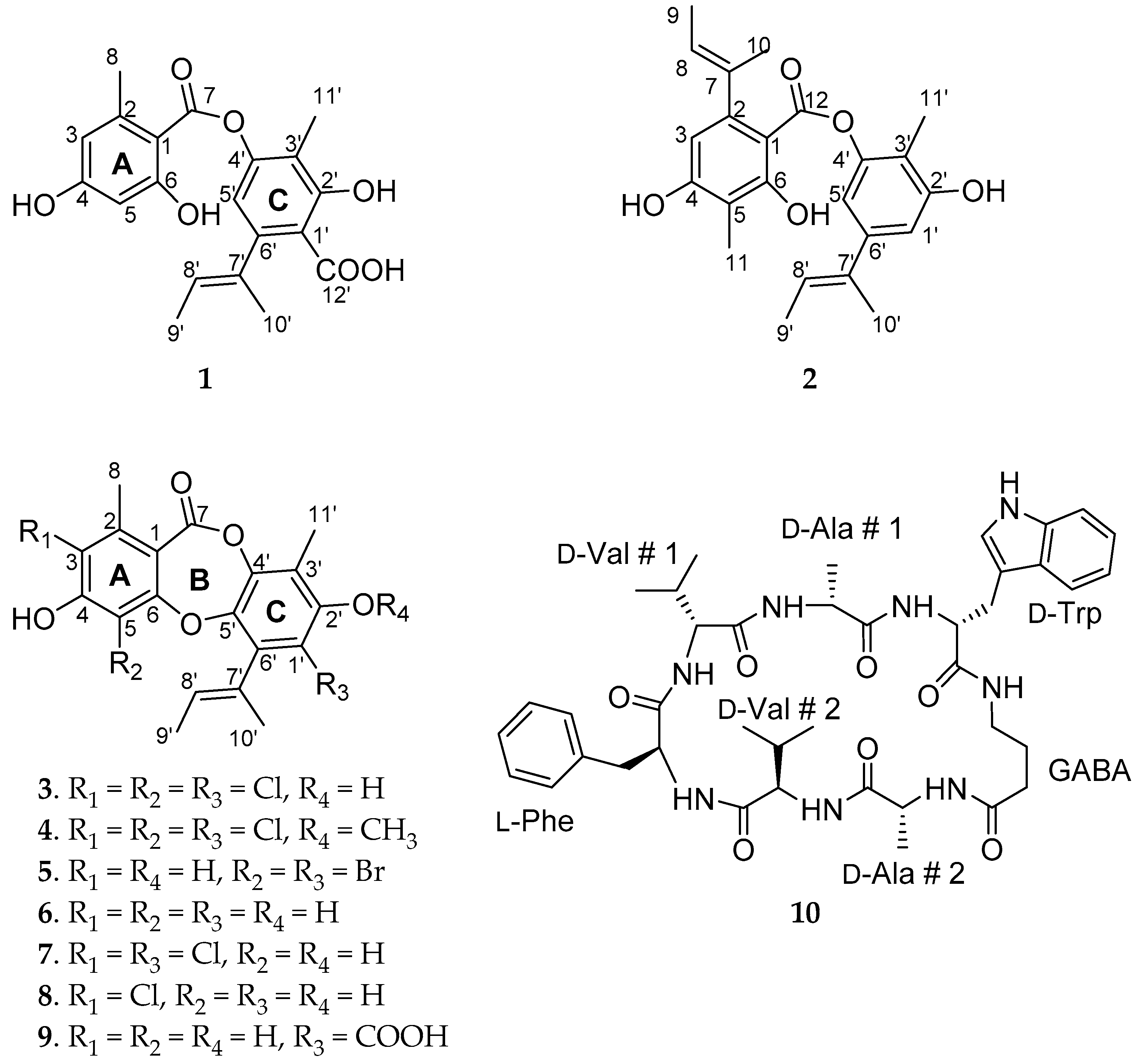
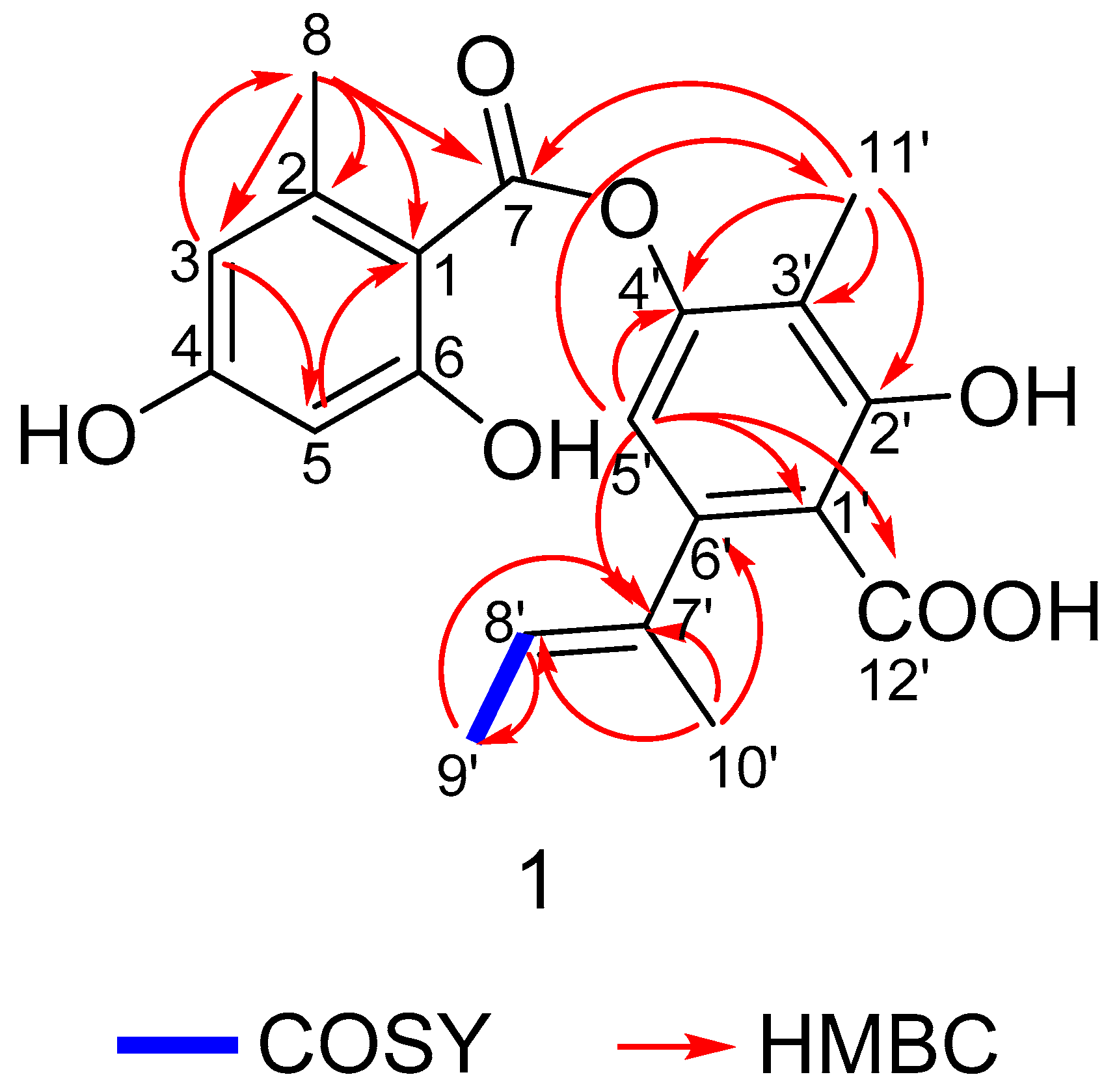
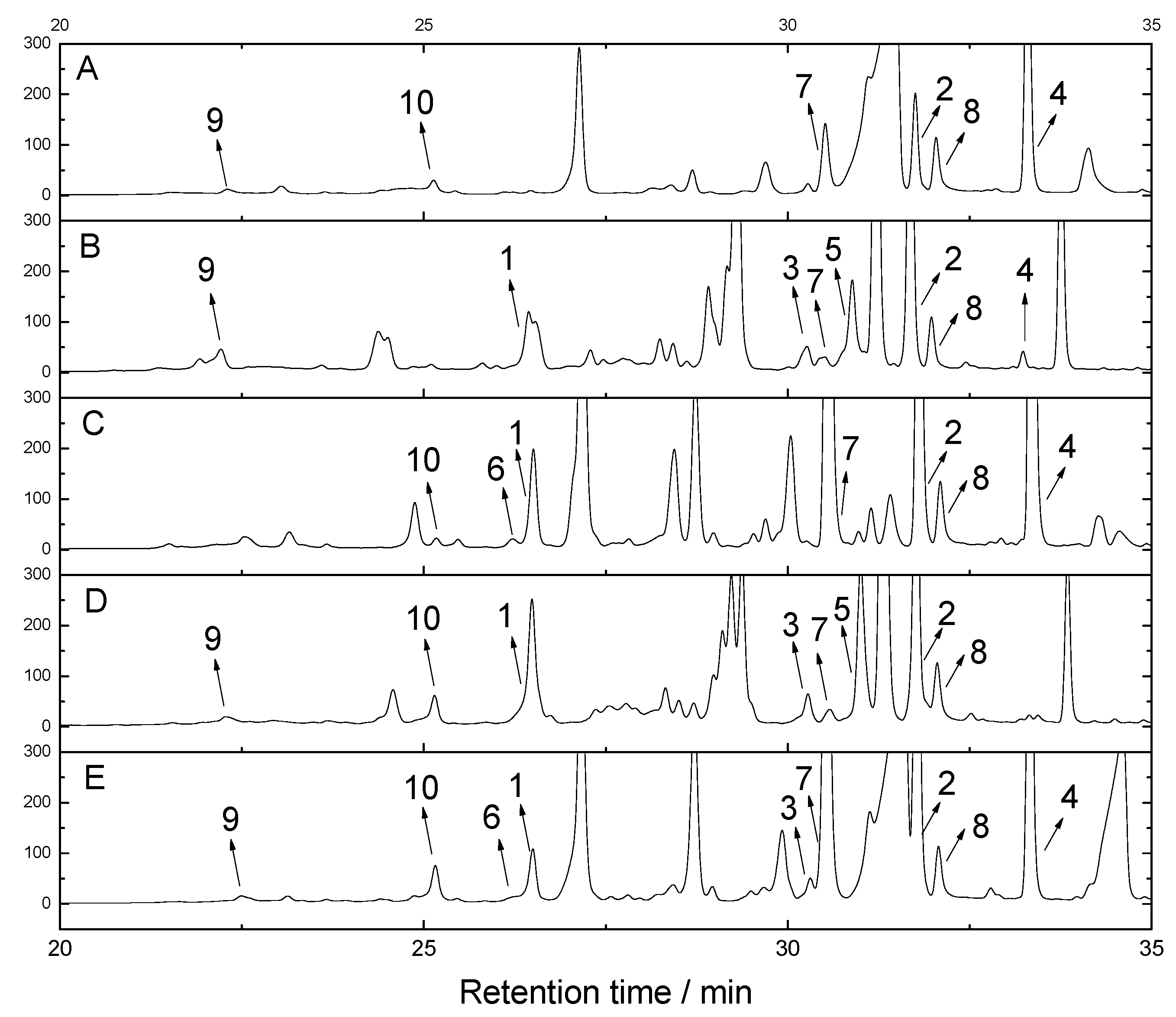
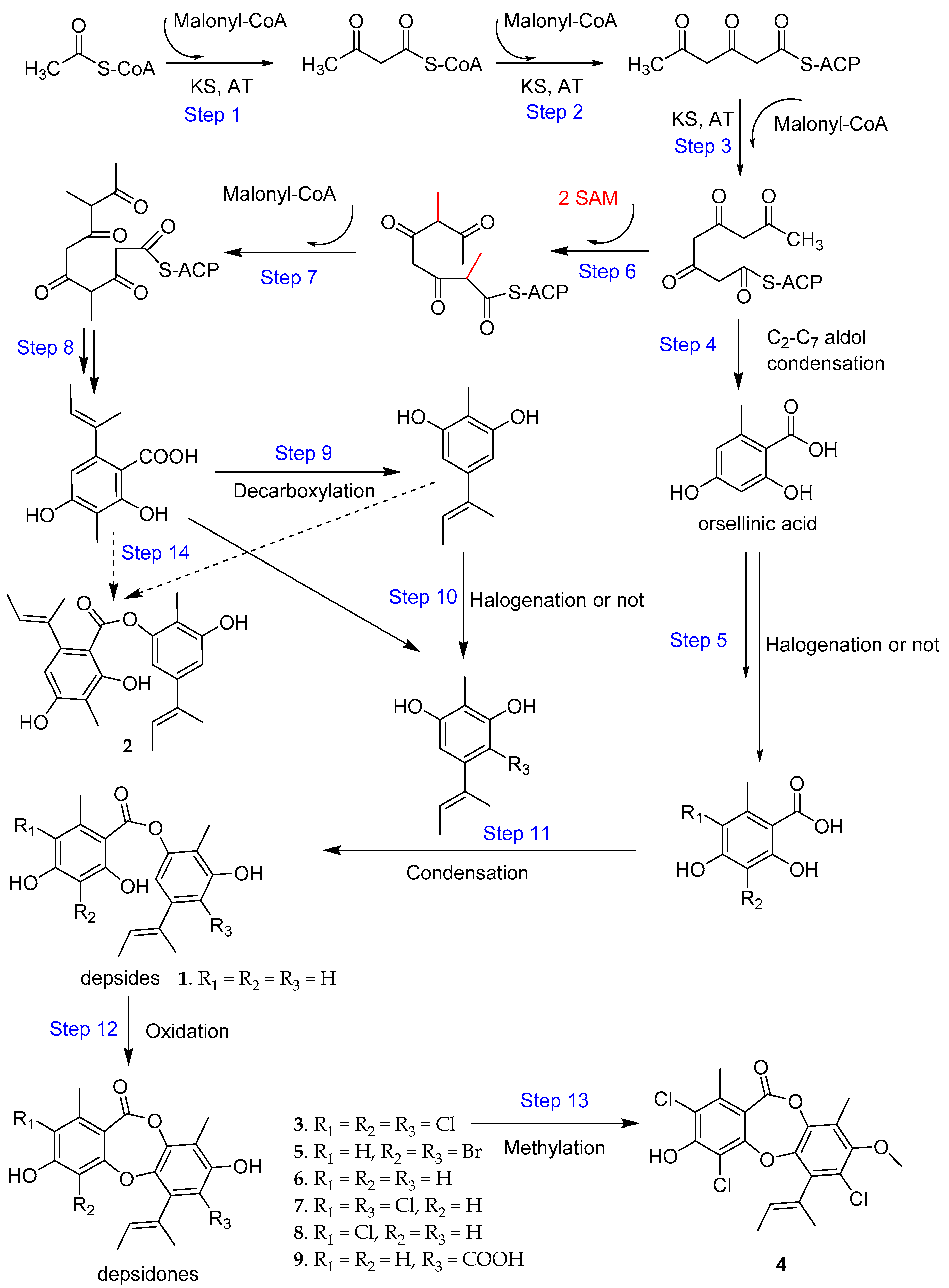
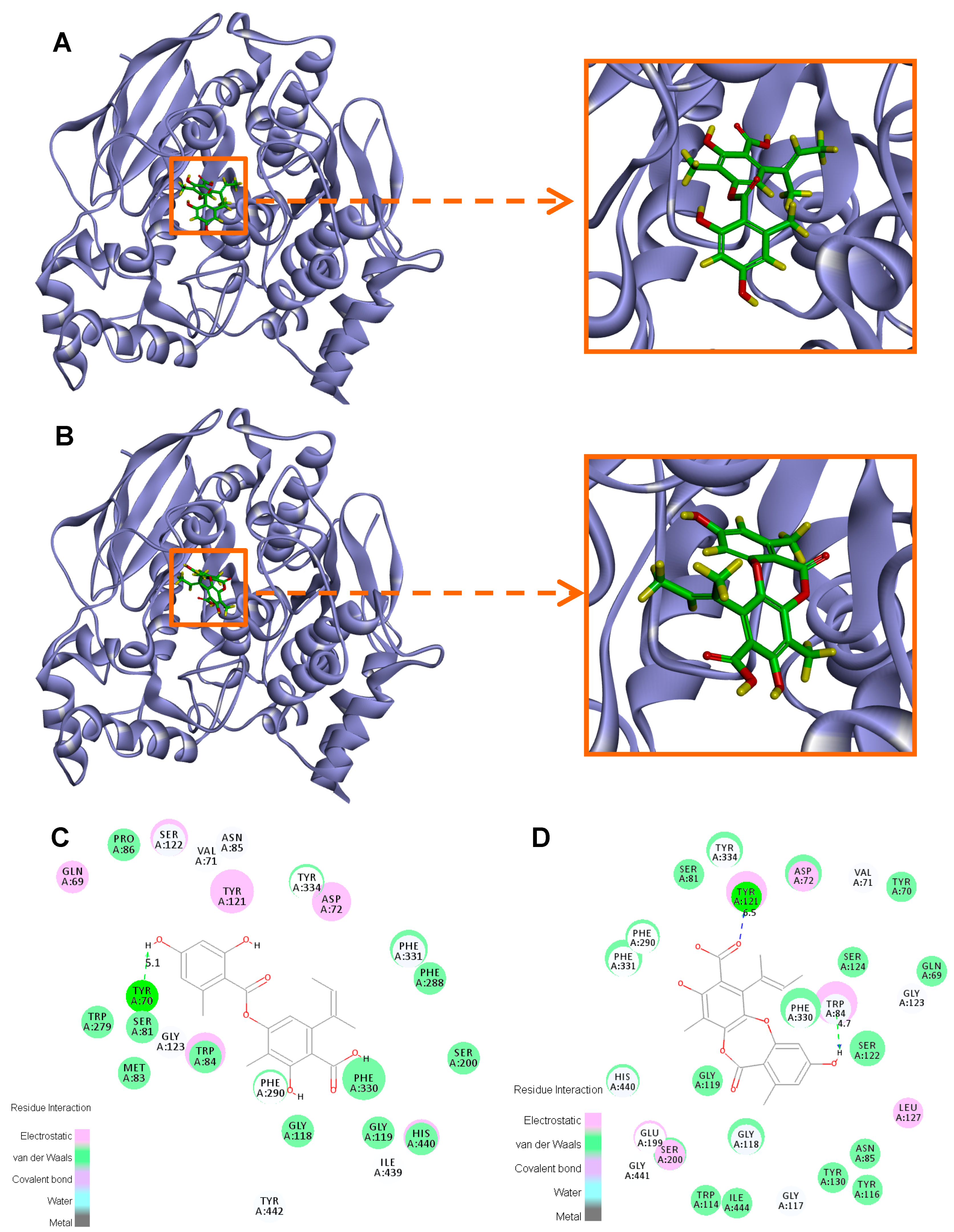
2. Results and Discussion
3. Materials and Methods
3.1. General Experimental Procedures
3.2. Fungal Material and Fermentation
3.3. Extraction and Isolation
3.4. HPLC Fingerprinting
3.5. Antimicrobial Assay
3.6. AChE Inhibition Assay
3.7. DPPH Free Radical Scavenging Assay
3.8. Brine Shrimp Larva Lethality Test
3.9. Molecular Docking Studies
4. Conclusions
5. Patents
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Deshmukh, S.K.; Prakash, V.; Ranjan, N. Marine fungi: A source of potential anticancer compounds. Front. Microbiol. 2018, 8, 2536–2560. [Google Scholar] [CrossRef] [PubMed]
- Fouillaud, M.; Venkatachalam, M.; Girard-Valenciennes, E.; Caro, Y.; Dufossé, L. Anthraquinones and derivatives from marine-derived fungi: Structural diversity and selected biological activities. Mar. Drugs 2016, 14, 64. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Quan, C.; Hou, X.; Fan, S.D. Potential pharmacological resources: Natural bioactive compounds from marine-derived fungi. Mar. Drugs 2016, 14, 76. [Google Scholar] [CrossRef] [PubMed]
- Overy, D.P.; Bayman, P.; Kerr, R.G.; Bills, G.F. An assessment of natural product discovery from marine (Sensu strictu) and marine-derived fungi. Mycology 2014, 5, 145–167. [Google Scholar] [CrossRef] [PubMed]
- Williams, R.B.; Henrikson, J.C.; Hoover, A.R.; Lee, A.E.; Cichewicz, R.H. Epigenetic remodeling of the fungal secondary metabolome. Org. Biomol. Chem. 2008, 6, 1895–1897. [Google Scholar] [CrossRef] [PubMed]
- Beau, J.; Mahid, N.; Burda, W.N.; Harrington, L.; Shaw, L.N.; Mutka, T.; Kyle, D.E.; Barisic, B.; Olphen, A.V.; Baker, B.J. Epigenetic tailoring for the production of anti-infective cytosporones from the marine fungus Leucostoma persoonii. Mar. Drugs 2012, 10, 762–774. [Google Scholar] [CrossRef] [PubMed]
- Laroussi, M.; Richardson, J.P.; Dobbs, F.C. Effects of nonequilibrium atmospheric pressure plasmas on the heterotrophic pathways of bacteria and on their cell morphology. Appl. Phys. Lett. 2002, 81, 772–774. [Google Scholar] [CrossRef]
- Bachmann, B.O.; Lanen, S.G.V.; Baltz, R.H. Microbial genome mining for accelerated natural products discovery: Is a renaissance in the making? J. Ind. Microbiol. Biotechnol. 2014, 41, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Brakhage, A.A.; Schroeckh, V. Fungal secondary metabolites-strategies to activate silent gene clusters. Fungal Genet. Biol. 2011, 48, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Henrikson, J.C.; Hoover, A.R.; Joyner, P.M.; Cichewicz, R.H. A chemical epigenetics approach for engineering the in situ biosynthesis of a cryptic natural product from Aspergillus niger. Org. Biomol. Chem. 2009, 7, 435–438. [Google Scholar] [CrossRef] [PubMed]
- Shwab, E.K.; Bok, J.W.; Tribus, M.; Galehr, J.; Graessle, S.; Keller, N.P. Histone deacetylase activity regulates chemical diversity in Aspergillus. Eukaryot. Cell 2007, 6, 1656–1664. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Mu, J.; Dong, X.W.; Feng, Y.; Mao, R.T.; Xiu, Z.L. Preliminary research on mutagenic breeding of marine fungal strain AP2T1 by dielectric barrier discharge plasma. J. Dalian Jiaotong Univ. 2012, 33, 75–79. [Google Scholar]
- Chen, H.; Chen, Z.J.; Wu, M.B.; Deng, S.X. Screening the Fusarium graminearum, inhibitory mutant strain from Bacillus subtilis, by atmospheric-pressure plasma jet. J. Appl. Microbiol. 2010, 108, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.Y.; Li, S.; Hou, Y.M.; Xiu, Z.L. Mutagensis of strain Klebsiella pneumoniae producing 1,3-propanediol by cold plasma at atmospheric pressure. Chin. J. Process. Eng. 2008, 8, 555–560. [Google Scholar]
- Zhang, Y.; Mu, J.; Feng, Y.; Wen, L.X.; Han, J.Y. Four chlorinated depsidones from a seaweed-derived strain of Aspergillus unguis and their new biological activities. Nat. Prod. Res. 2014, 28, 503–506. [Google Scholar] [CrossRef] [PubMed]
- Lang, G.; Cole, A.L.; Blunt, J.W.; Robinson, W.T.; Munro, M.H. Excelsione, a depsidone from an endophytic fungus isolated from the New Zealand endemic tree Knightia excelsa. J. Nat. Prod. 2007, 70, 310–311. [Google Scholar] [CrossRef] [PubMed]
- Chomcheon, P.; Wiyakrutta, S.; Sriubolmas, N.; Ngamrojanavanich, N.; Kengtong, S.; Mahidol, C.; Ruchirawat, S.; Kittakoop, P. Aromatase inhibitory, radical scavenging, and antioxidant activities of depsidones and diaryl ethers from the endophytic fungus Corynespora cassiicola L36. Phytochemistry 2009, 70, 407–413. [Google Scholar] [CrossRef] [PubMed]
- Beach, W.F.; Richards, J.H. The nature of the alkyl groups in nidulin. J. Org. Chem. 1961, 26, 3011–3012. [Google Scholar] [CrossRef]
- Sureram, S.; Wiyakrutta, S.; Ngamrojanavanich, N.; Mahidol, C.; Ruchirawat, S.; Kittakoop, P. Depsidones, aromatase inhibitors and radical scavenging agents from the marine-derived fungus Aspergillus unguis CRI282-03. Planta Med. 2012, 78, 582–588. [Google Scholar] [CrossRef] [PubMed]
- Kamal, A.H.; Yasmin, K.Y.; Khan, Y.A.; Qureshi, I.H.; Qureshi, A.A. Biochemistry of microorganisms. X. isolation, structure, and stereochemistry of yasimin and other metabolic products of Aspergillus unguis. Pak. J. Sci. Ind. Res. 1970, 13, 244–250. [Google Scholar]
- Sureram, S.; Kesornpun, C.; Mahidol, C.; Ruchirawat, S.; Kittakoop, P. Directed biosynthesis through biohalogenation of secondary metabolites of the marine-derived fungus Aspergillus unguis. RSC Adv. 2013, 3, 1781–1788. [Google Scholar] [CrossRef]
- Kawahara, N.; Nakajima, S.; Satoh, Y.; Yamazaki, M.; Kawai, K.I. Studies on fungal products. XVIII: Isolation and structures of a new fungal depsidone related to nidulin and a new phthalide from Emericella unguis. Chem. Pharm. Bull. 1988, 36, 1970–1975. [Google Scholar] [CrossRef]
- Malmstrom, J. Unguisins A and B: New cyclic peptides from the marine-derived fungus Emericella unguis. J. Nat. Prod. 1999, 62, 787–789. [Google Scholar] [CrossRef] [PubMed]
- Hunter, L.; Chung, J.H. Total synthesis of unguisin A. J. Org. Chem. 2011, 76, 5502–5505. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.G.; Lee, A.S.Y.; Huang, Y.C.; Flotow, H.; Ng, S.; Butler, M.S.; Buss, A.D. Agonodepsides A and B: Two new depsides from a filamentous fungus F7524. J. Nat. Prod. 2002, 65, 1037–1038. [Google Scholar] [CrossRef] [PubMed]
- Sierankiewicz, J.; Gatenbeck, S. The biosynthesis of nidulin and trisdechloronornidulin. Acta Chem. Scand. 1973, 27, 2710–2716. [Google Scholar] [CrossRef] [PubMed]
- Schümann, J.; Hertweck, C. Advances in cloning, functional analysis and heterologous expression of fungal polyketide synthase genes. J. Biotechnol. 2006, 124, 690–703. [Google Scholar] [CrossRef] [PubMed]
- Okoye, F.B.C.; Lu, S.; Nworu, C.S.; Esimone, C.O.; Proksch, P.; Chadli, A.; Debbab, A. Depsidone and diaryl ether derivatives from the fungus Corynespora cassiicola, an endophyte of Gongronema latifolium. Tetrahedron Lett. 2014, 44, 4210–4214. [Google Scholar]
- Ece, A.; Pejin, B. A computational insight into acetylcholinesterase inhibitory activity of a new lichen depsidone. J. Enzym. Inhib. Med. Chem. 2015, 30, 528–532. [Google Scholar] [CrossRef] [PubMed]
- Carlier, P.R.; Du, D.M.; Han, Y.F.; Liu, J.; Perola, E.; Williams, I.D.; Pang, Y.P. Dimerization of an inactive fragment of huperzine a produces a drug with twice the potency of the natural product. Angew. Chem. Int. Ed. 2010, 39, 1775–1777. [Google Scholar] [CrossRef]
- Sippl, W.; Contreras, J.M.; Parrot, I.; Rival, Y.M.; Wermuth, C.G. Structure-based 3D QSAR and design of novel acetylcholinesterase inhibitors. J. Comput. Aided. Mol. Des. 2001, 15, 395–410. [Google Scholar] [CrossRef] [PubMed]
- Contreras, J.M.; Parrot, I.; Sippl, W.; Rival, Y.M.; Wermuth, C.G. Design, synthesis, and structure−activity relationships of a series of 3-[2-(1-benzylpiperidin-4-yl)ethylamino]pyridazine derivatives as acetylcholinesterase inhibitors. J. Med. Chem. 2001, 44, 2707–2718. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, S.R.M.; Mohamed, G.A.; Haidari, R.A.A.; El-Kholy, A.A.; Zayed, M.F.; Khayat, M.T. Biologically active fungal depsidones: Chemistry, biosynthesis, structural characterization, and bioactivities. Fitoterapia 2018. [Google Scholar] [CrossRef] [PubMed]
- Neamati, N.; Hong, H.X.; Mazumder, A.; Wang, S.M.; Sunder, S.; Nicklaus, M.C.; Milne, G.W.A.; Proksa, B.; Pommier, Y. Depsides and depsidones as inhibitors of HIV-1 integrase: Discovery of novel inhibitors through 3D database searching. J. Med. Chem. 1997, 40, 942–951. [Google Scholar] [CrossRef] [PubMed]
- Pejin, B.; Tommonaro, G.; Iodice, C.; Tesevic, V.; Vajs, V.; Rosa, S.D. A new depsidone of Lobaria pulmonaria with acetylcholinesterase inhibition activity. J. Enzym. Inhib. Med. Chem. 2013, 28, 876–878. [Google Scholar] [CrossRef] [PubMed]
- Reddy, R.G.; Veeraval, L.; Maitra, S.; Chollet-Krugler, M.; Tomasi, S.; Dévéhat, F.L.; Boustie, J.; Chakravarty, S. Lichen-derived compounds show potential for central nervous system therapeutics. Phytomedicine 2016, 23, 1527–1534. [Google Scholar] [CrossRef] [PubMed]
- Wen, L.X.; Zhang, Y.; Mu, J.; Han, J.Y.; Xiu, Z.L. Screening plasma-induced bioactive mutants of sporogenous marine fungus using a modified method. J. Dalian Jiaotong Univ. 2014, 35, 87–92. [Google Scholar]
- Santhosh, S.K.; Venugopal, A.; Radhakrishnan, M.C. Study on the phytochemical, antibacterial and antioxidant activities of Simarouba glauca. Indian J. Biol. Sci. 2016, 2, 119–124. [Google Scholar] [CrossRef]
- Fothergill, A.W. Antifungal susceptibility testing: Clinical Laboratory and Standards Institute (CLSI) methods. In Interactions of Yeasts, Moulds, and Antifungal Agents: How to Detect Resistance; Hall, G.S., Ed.; Humana Press: New York, NY, USA, 2012; Volume 3, pp. 65–74. ISBN 979-1-58829-847-8. [Google Scholar]
- Weinstein, M.P. Broth and agar dilution antimicrobial susceptibility testing process. In M07, Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically, 11th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018; pp. 15–52. ISBN 1-56238-837-1. [Google Scholar]
- Ellman, G.L.; Courtney, K.D.; Andres, V.; Featherstone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef] [Green Version]
- Sharma, O.P.; Bhat, T.K. DPPH antioxidant assay revisited. Food Chem. 2009, 113, 1202–1205. [Google Scholar] [CrossRef]
- Hisem, D.; Hrouzek, P.; Tomek, P.; Tomšíčková, J.; Zapomělová, E.; Skácelová, K.; Lukešová, A.; Kopecký, J. Cyanobacterial cytotoxicity versus toxicity to brine shrimp Artemia salina. Toxicon 2011, 57, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Robertson, D.H.; Brooks, C.L.; Vieth, M. Detailed analysis of grid-based molecular docking: A case study of CDOCKER A CHARMm-based MD docking algorithm. J. Comput. Chem. 2003, 24, 1549–1562. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | δc a, Type | δHb, Mult (J in Hz) | Position | Δc a, Type | δHb, Mult (J in Hz) |
|---|---|---|---|---|---|
| 1 | 104.5, C | 3′ | 116.9, C | ||
| 2 | 144.7, C | 4′ | 161.8, C | ||
| 3 | 113.5, CH | 6.28, dd, (2.0, 0.5) | 5′ | 114.3, CH | 6.25, s |
| 4 | 166.9, C | 6′ | 148.7, C | ||
| 5 | 102.2, CH | 6.19, d, (2.4) | 7′ | 141.1, C | |
| 6 | 165.9, C | 8′ | 120.6, CH | 5.34, dq, (6.8, 1.3) | |
| 7 | 171.3, C | 9′ | 14.0, CH3 | 1.70, dd, (6.8, 0.8) | |
| 8 | 24.6, CH3 | 2.59, s | 10′ | 18.8, CH3 | 1.92, s |
| 1′ | 116.7, C | 11′ | 9.6, CH3 | 2.01, s | |
| 2′ | 151.3, C | 12′ | 175.2, C |
| Strains | Zone of Inhibition (mm) # | ||||
|---|---|---|---|---|---|
| Compounds | P. aeruginosa | MRSA | V. parahemolyticus * | C. albicans | |
| 1 | 7.7 ± 0.6 | 12.0 ± 1.0 | 9.3 ± 0.6 | 8.3 ± 0.6 | |
| 2 | - | - | 8.0 ± 0.0 | 6.0 ± 0.5 | |
| 3 | 12.2 ± 0.3 | - | - | 12.1 ± 0.2 | |
| 4 | 9.5 ± 0.6 | 9.0 ± 0.0 | - | 9.0 ± 0.0 | |
| 5 | 11.9 ± 0.2 | 14.5 ± 0.6 | 6.3 ± 0.6 | 7.5 ± 0.6 | |
| 6 | 8.7 ± 2.1 | 7.0 ± 0.0 | 7.0 ± 0.0 | 8.0 ± 0.5 | |
| 7 | 10.0 ± 0.0 | 13.7 ± 2.3 | 9.7 ± 0.6 | 7.9 ± 0.2 | |
| 8 | 11.1 ± 0.2 | 17.0 ± 0.0 | 8.3 ± 0.6 | 10.0 ± 0.0 | |
| 9 | 16.0 ± 0.0 | 17.7 ± 0.6 | 8.3 ± 0.6 | 15.5 ± 2.2 | |
| 10 | - | - | 9.0 ± 0.0 | 6.0 ± 0.5 | |
| Ampicillin | 12.0 ± 1.4 | 14.0 ± 0.0 | 9.0 ± 0.0 | - | |
| Ketoconazole | - | - | - | 22.0 ± 0.0 | |
| Strains | MIC (μM) | ||||
|---|---|---|---|---|---|
| Compounds | P. aeruginosa | MRSA | V. parahemolyticus | C. albicans | |
| 1 | 102.4 | 51.2 | >102.4 | 12.8 | |
| 2 | - | - | >102.4 | - | |
| 3 | >102.4 | - | - | >102.4 | |
| 4 | >102.4 | 102.4 | - | >102.4 | |
| 5 | 6.4 | 25.6 | >102.4 | >102.4 | |
| 6 | 102.4 | >102.4 | >102.4 | >102.4 | |
| 7 | 25.6 | >102.4 | >102.4 | >102.4 | |
| 8 | 51.2 | 25.6 | >102.4 | >102.4 | |
| 9 | 102.4 | 12.8 | >102.4 | 6.4 | |
| 10 | - | - | >102.4 | - | |
| Ampicillin | 1.6 | 6.4 | 102.4 | - | |
| Ketoconazole | - | - | - | 1.9 | |
| Compounds | AChE Inhibitory Activity (IC50, μM) | DPPH Scavenging Activity (EC50, μM) | Larvicidal Activity (LC50, μM) |
|---|---|---|---|
| 1 | >102.4 | >102.4 | >102.4 |
| 2 | >102.4 | >102.4 | >102.4 |
| 3 | >102.4 | >102.4 | 12.8 |
| 4 | >102.4 | >102.4 | 4.5 |
| 5 | >102.4 | >102.4 | 12.8 |
| 6 | >102.4 | >102.4 | >102.4 |
| 7 | >102.4 | >102.4 | 72 |
| 8 | >102.4 | >102.4 | 75.9 |
| 9 | 56.8 | >102.4 | 51.2 |
| 10 | >102.4 | >102.4 | >102.4 |
| Donepezil | 0.3 | - | - |
| Vitamin C | - | 31.8 | - |
| Hg(NO3)2 | - | - | 77.0 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, W.-C.; Bao, H.-Y.; Liu, Y.-Y.; Nie, Y.-Y.; Yang, J.-M.; Hong, P.-Z.; Zhang, Y. Depsidone Derivatives and a Cyclopeptide Produced by Marine Fungus Aspergillus unguis under Chemical Induction and by Its Plasma Induced Mutant. Molecules 2018, 23, 2245. https://doi.org/10.3390/molecules23092245
Yang W-C, Bao H-Y, Liu Y-Y, Nie Y-Y, Yang J-M, Hong P-Z, Zhang Y. Depsidone Derivatives and a Cyclopeptide Produced by Marine Fungus Aspergillus unguis under Chemical Induction and by Its Plasma Induced Mutant. Molecules. 2018; 23(9):2245. https://doi.org/10.3390/molecules23092245
Chicago/Turabian StyleYang, Wen-Cong, Hai-Yan Bao, Ya-Yue Liu, Ying-Ying Nie, Jing-Ming Yang, Peng-Zhi Hong, and Yi Zhang. 2018. "Depsidone Derivatives and a Cyclopeptide Produced by Marine Fungus Aspergillus unguis under Chemical Induction and by Its Plasma Induced Mutant" Molecules 23, no. 9: 2245. https://doi.org/10.3390/molecules23092245
APA StyleYang, W. -C., Bao, H. -Y., Liu, Y. -Y., Nie, Y. -Y., Yang, J. -M., Hong, P. -Z., & Zhang, Y. (2018). Depsidone Derivatives and a Cyclopeptide Produced by Marine Fungus Aspergillus unguis under Chemical Induction and by Its Plasma Induced Mutant. Molecules, 23(9), 2245. https://doi.org/10.3390/molecules23092245