
Clitoria ternatea Flower Petal Extract Inhibits Adipogenesis and Lipid Accumulation in 3T3-L1 Preadipocytes by Downregulating Adipogenic Gene Expression

 ,
, 
Abstract
:1. Introduction
2. Results
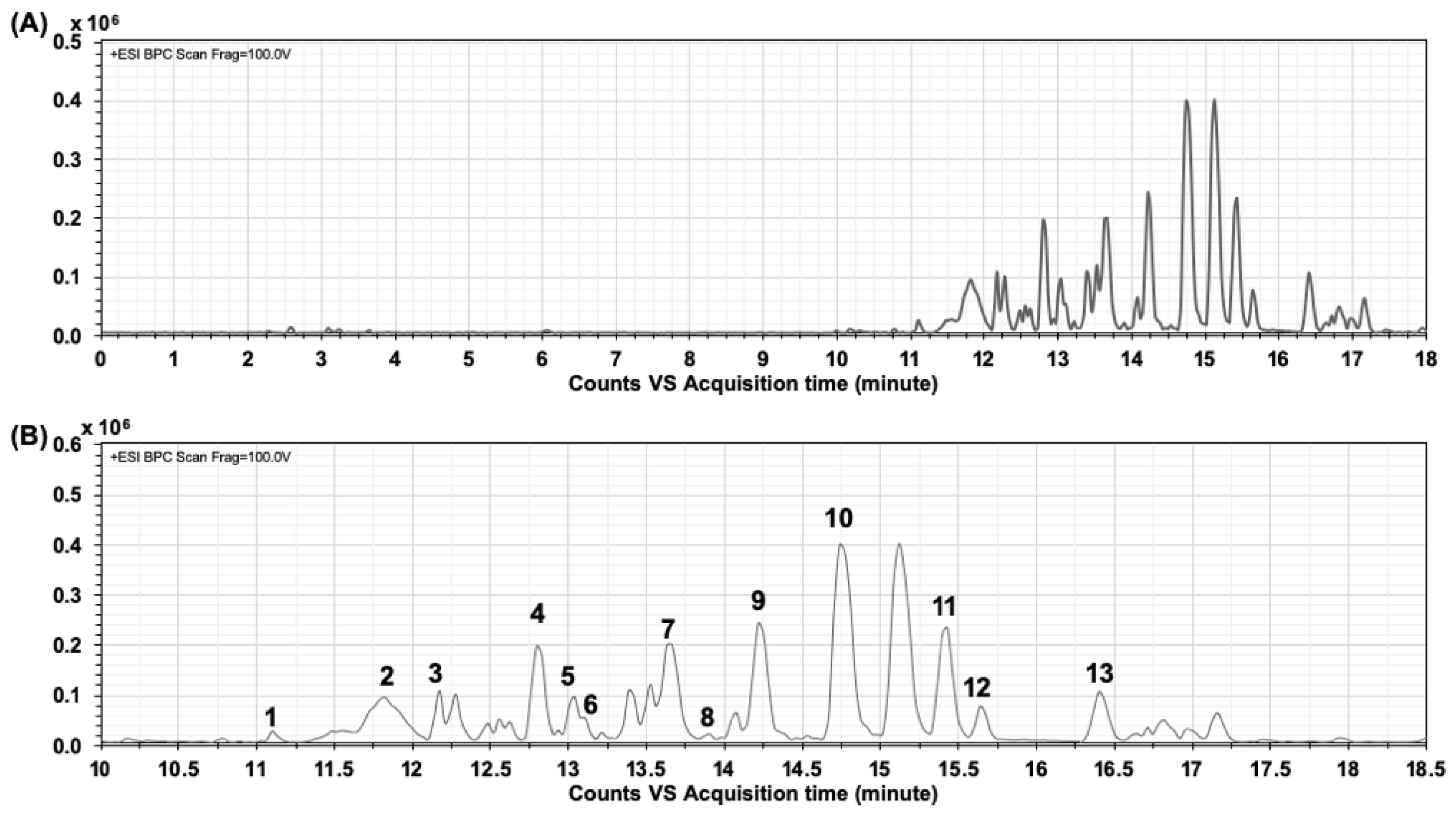
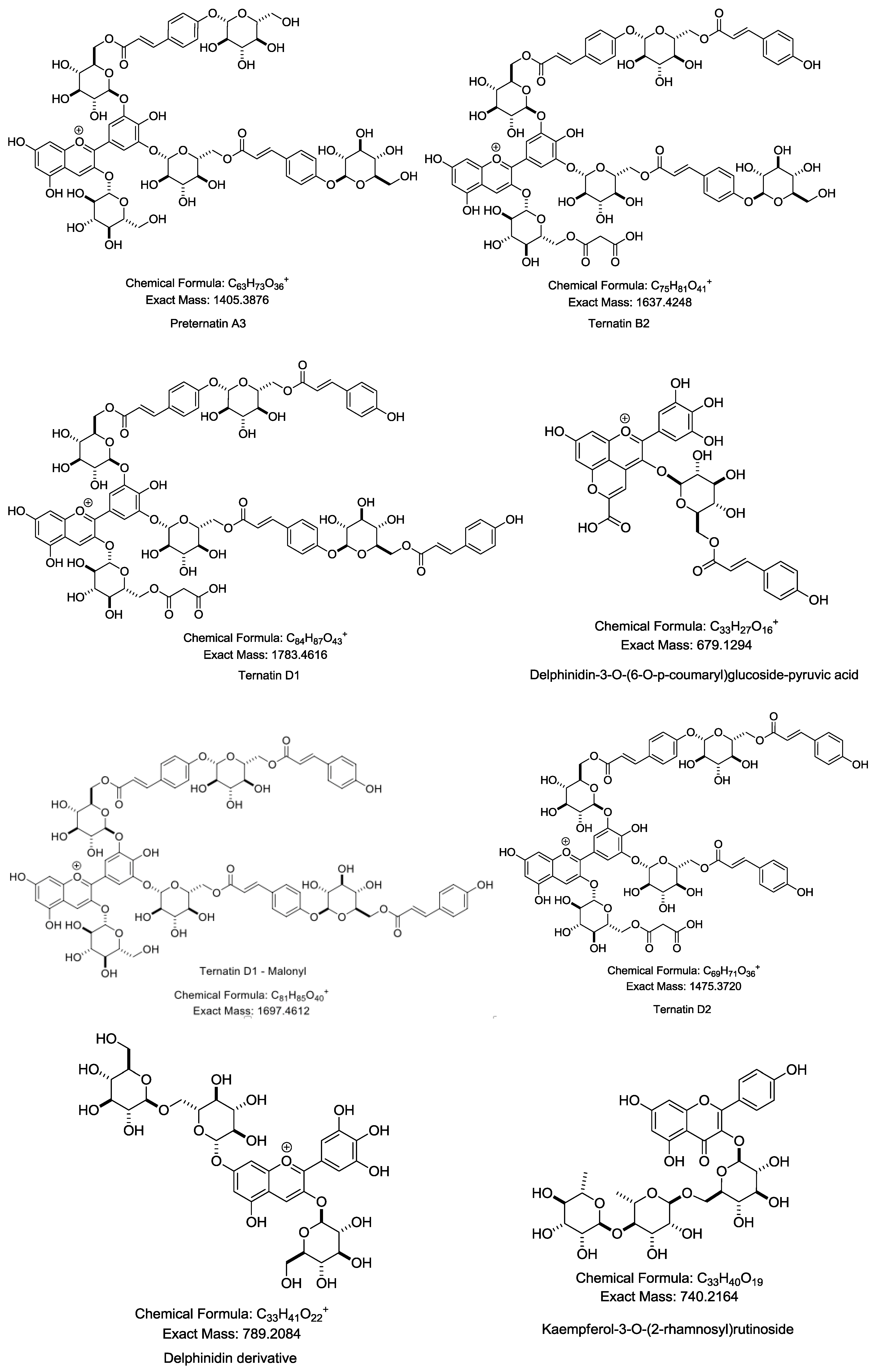
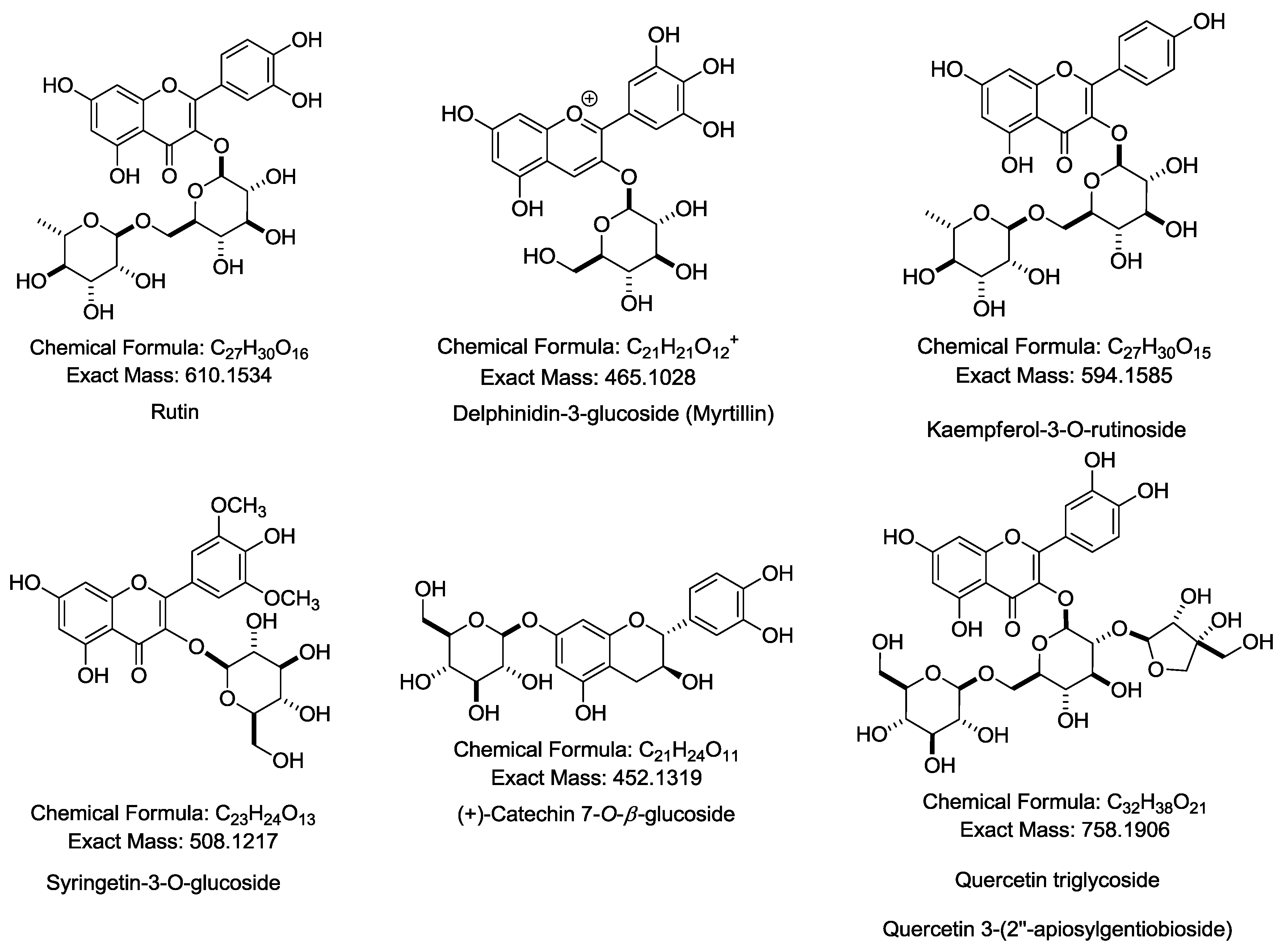
2.1. Characterization and Identification of Phenolic Compounds in CTE by LC-MS/MS
2.2. Effects of CTE on Cell Viability of 3T3-L1 Cells
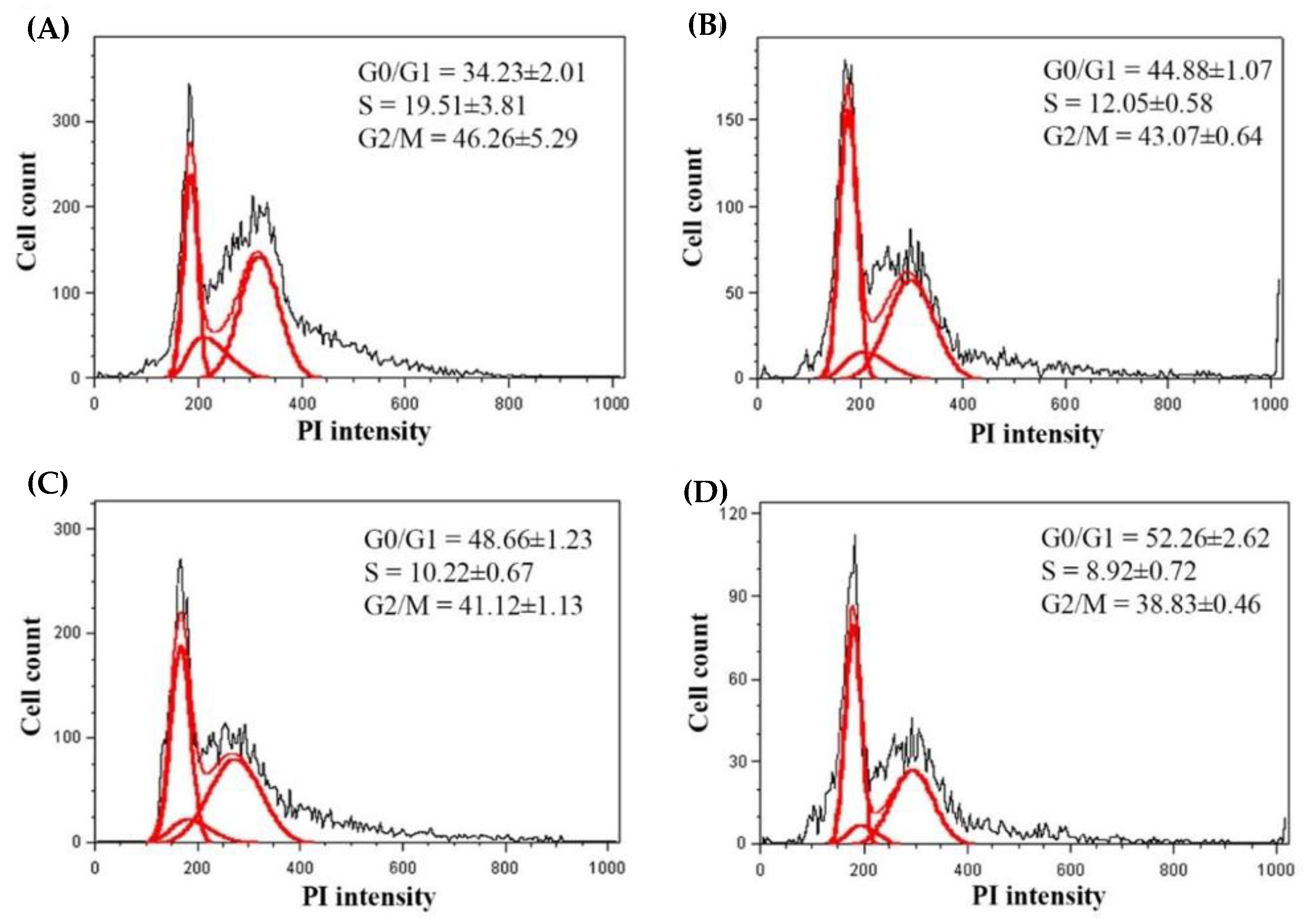
2.3. Effects of CTE on the Cell Cycle of 3T3-L1 Cells
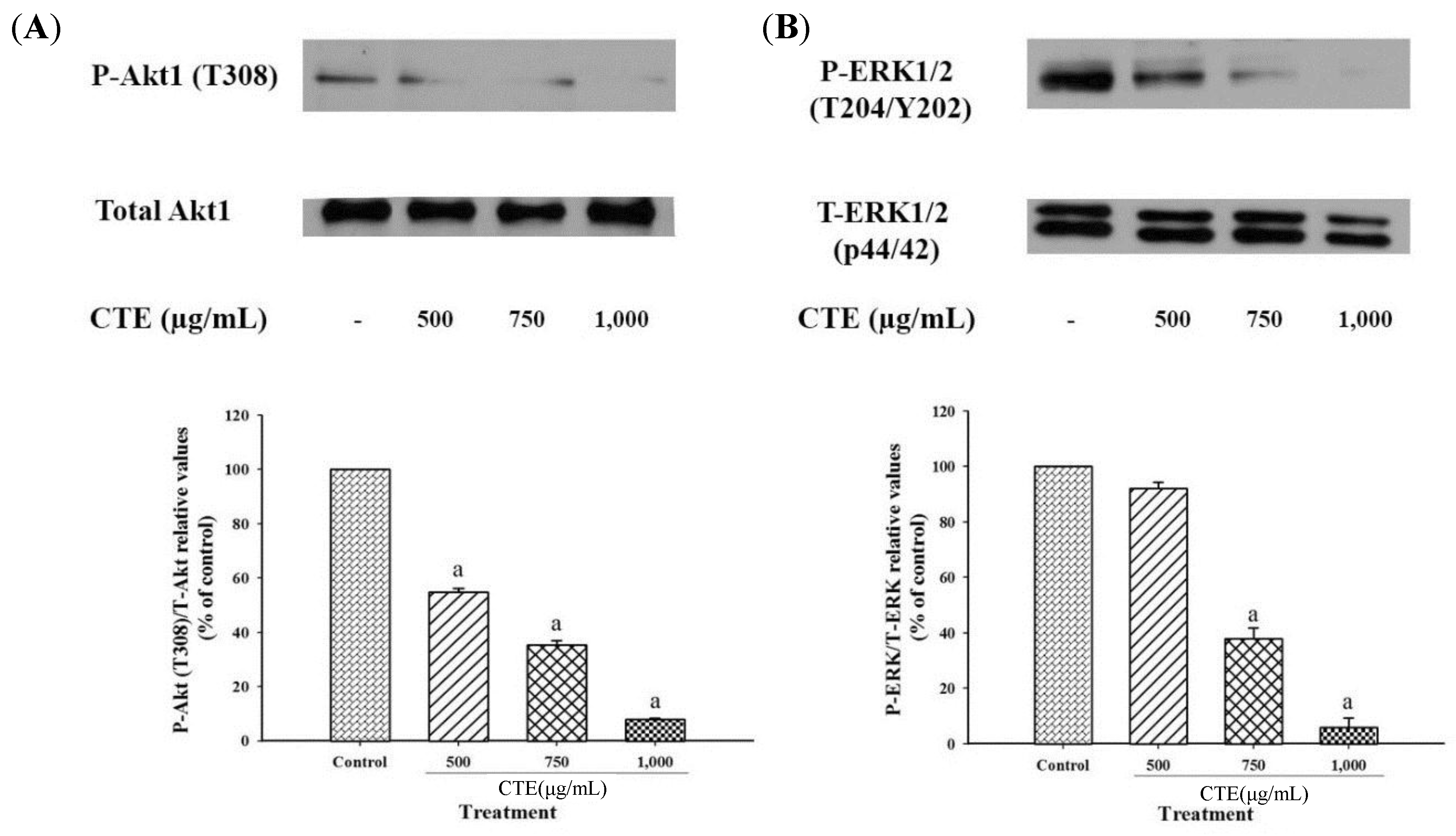
2.4. Effects of CTE on Akt1 and ERK1/2 Signaling Pathways in 3T3-L1 Cells
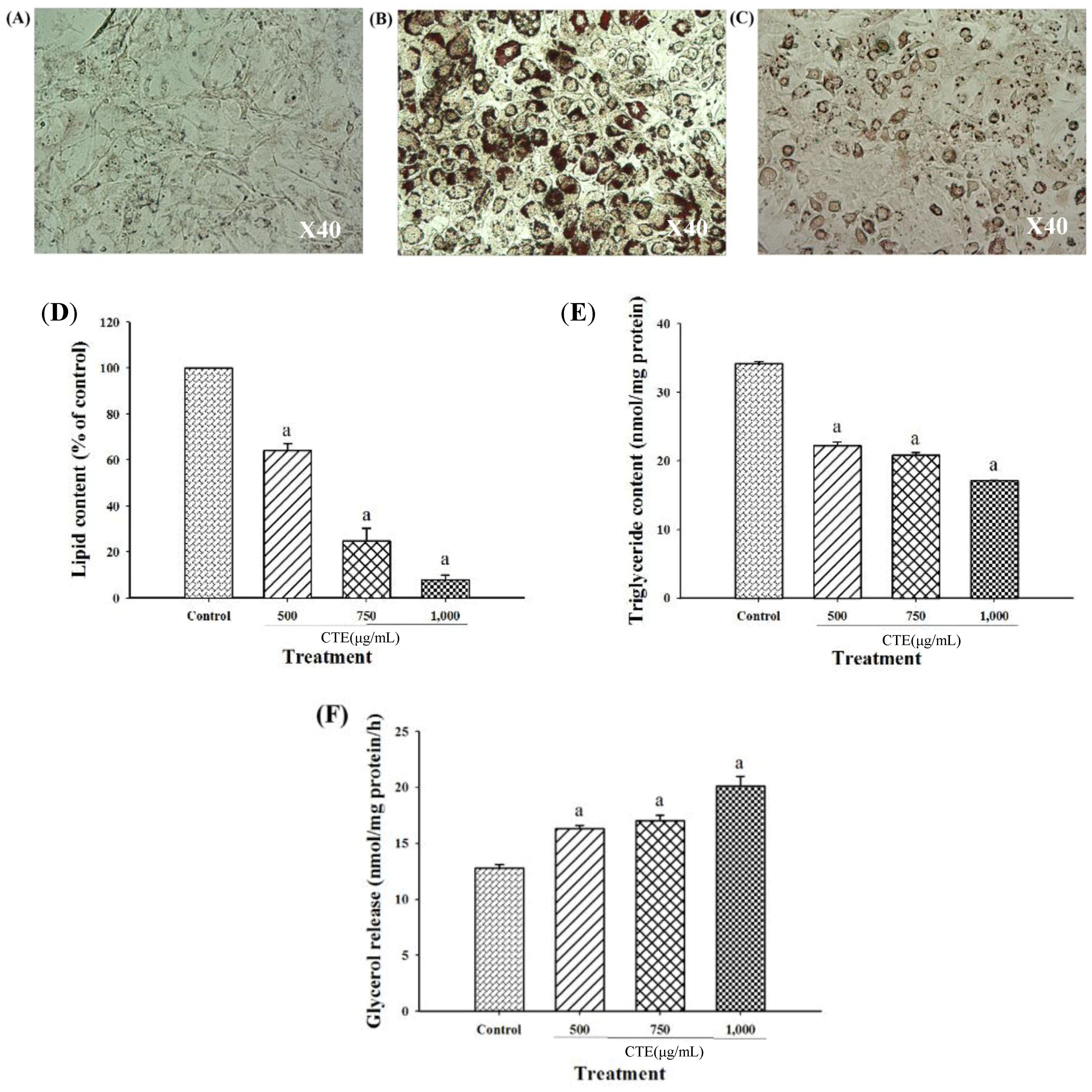
2.5. Effects of CTE on Lipid Accumulation in 3T3-L1 Cells
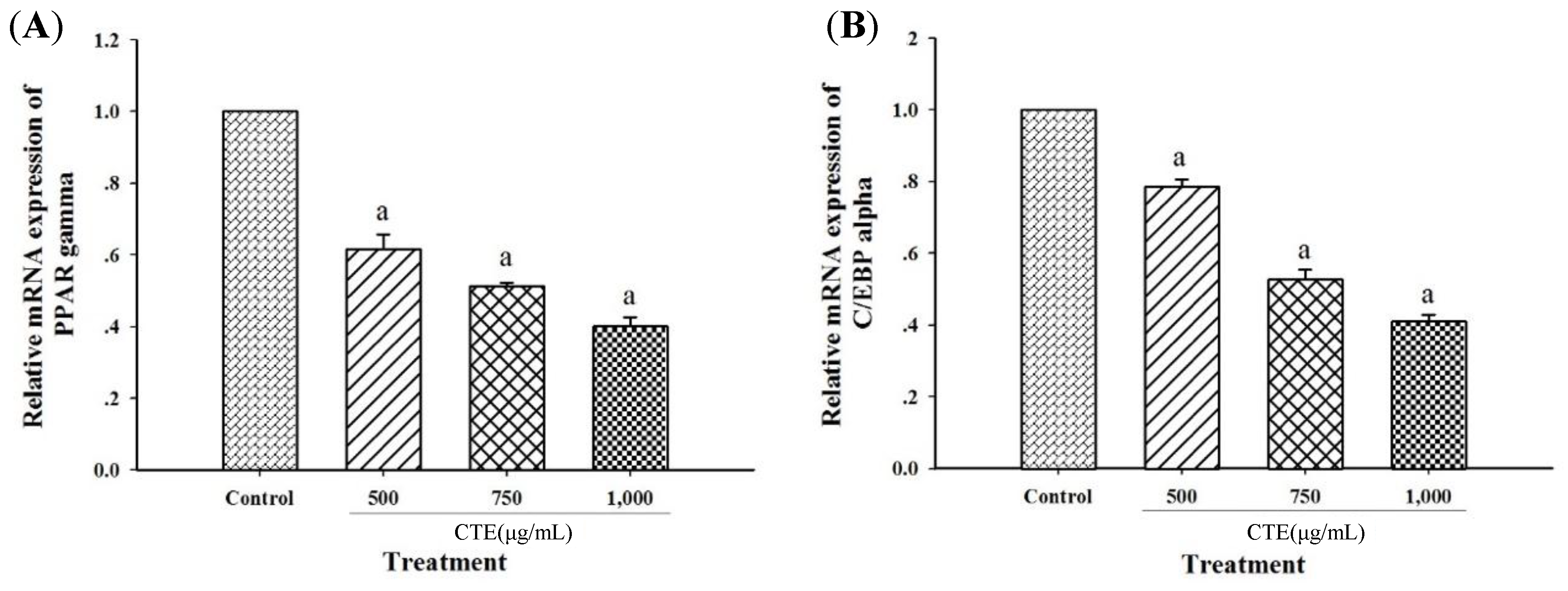
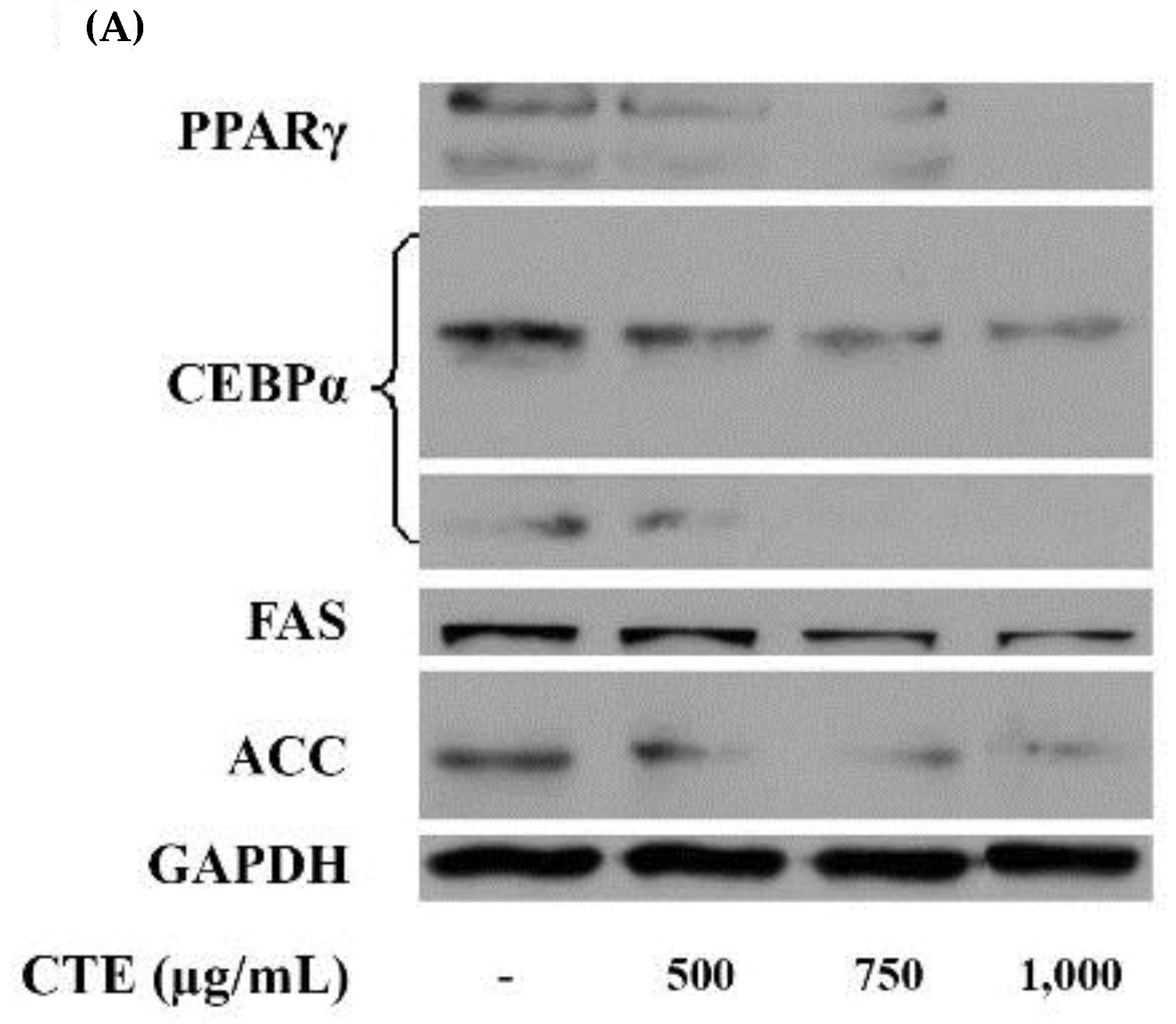
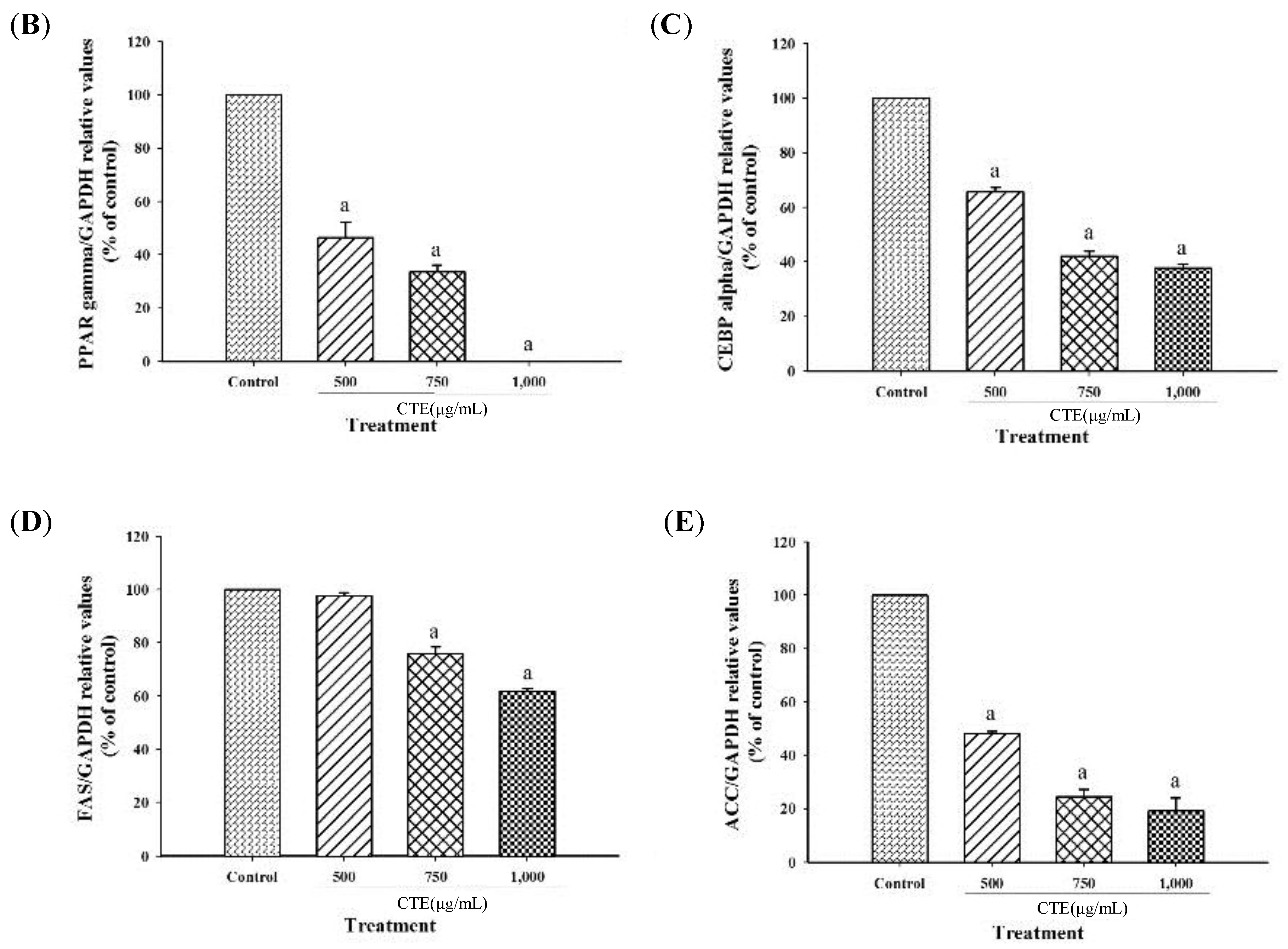
2.6. Effects of CTE on Adipogenic Transcription Factors and Lipogenic Enzymes
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Plant Materials
4.3. Sample Preparation for LC-MS/MS
4.4. Characterization of CTE
4.5. Culture and Differentiation of 3T3-L1 Cells
4.6. Cell Viability Assay
4.7. Oil Red O Assay
4.8. Cell Cycle by Flow Cytometry
4.9. Western Blot Analysis
4.10. Triglyceride Accumulation
4.11. Lipolysis Assay
4.12. Real-time PCR
4.13. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
Abbreviations
References
- Flatt, J.P. Differences in basal energy expenditure and obesity. Obesity 2007, 15, 2546–2548. [Google Scholar] [CrossRef] [PubMed]
- Redinger, R.N. Fat storage and the biology of energy expenditure. Transl. Res. 2009, 154, 52–60. [Google Scholar] [CrossRef]
- Greenberg, A.; McDaniel, M. Identifying the links between obesity, insulin resistance and β-cell function: potential role of adipocyte-derived cytokines in the pathogenesis of type 2 diabetes. Eur. J. Clin. Investig. 2002, 32, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Guilherme, A.; Virbasius, J.V.; Puri, V.; Czech, M.P. Adipocyte dysfunctions linking obesity to insulin resistance and type 2 diabetes. Nat. Rev. Mol. Cell Biol. 2008, 9, 367–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kahn, B.B.; Flier, J.S. Obesity and insulin resistance. J. Clin. Investig. 2000, 106, 473–481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno-Navarrete, J.M.; Fernández-Real, J.M. Adipocyte differentiation. In Adipose Tissue Biology; Springer: Basel, Switzerland, 2012; pp. 17–38. [Google Scholar]
- Bost, F.; Aouadi, M.; Caron, L.; Binetruy, B. The role of MAPKs in adipocyte differentiation and obesity. Biochimie 2005, 87, 51–56. [Google Scholar] [CrossRef] [Green Version]
- Chuang, C.C.; Yang, R.S.; Tsai, K.S.; Ho, F.M.; Liu, S.H. Hyperglycemia enhances adipogenic induction of lipid accumulation: Involvement of extracellular signal-regulated protein kinase 1/2, phosphoinositide 3-kinase/Akt, and peroxisome proliferator-activated receptor gamma signaling. Endocrinology 2007, 148, 4267–4275. [Google Scholar] [CrossRef] [PubMed]
- Feve, B. Adipogenesis: Cellular and molecular aspects. Best Pract. Res. Clin. Endocrinol. Metab. 2005, 19, 483–499. [Google Scholar] [CrossRef] [PubMed]
- Jia, X.; Chang, T.; Wilson, T.W.; Wu, L. Methylglyoxal mediates adipocyte proliferation by increasing phosphorylation of Akt1. PLoS ONE 2012, 7, e36610. [Google Scholar] [CrossRef]
- Ferguson, B.S.; Nam, H.; Morrison, R.F. Curcumin inhibits 3T3-L1 preadipocyte proliferation by mechanisms involving post-transcriptional p27 regulation. Biochem. Biophys. Rep. 2016, 5, 16–21. [Google Scholar] [CrossRef]
- Jaworski, K.; Sarkadi-Nagy, E.; Duncan, R.E.; Ahmadian, M.; Sul, H.S. Regulation of triglyceride metabolism. IV. Hormonal regulation of lipolysis in adipose tissue. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 293, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Prusty, D.; Park, B.-H.; Davis, K.E.; Farmer, S.R. Activation of MEK/ERK signaling promotes adipogenesis by enhancing peroxisome proliferator-activated receptor γ (PPARγ) and C/EBPα gene expression during the differentiation of 3T3-L1 preadipocytes. J. Biol. Chem. 2002, 277, 46226–46232. [Google Scholar] [CrossRef]
- Tang, Q.Q.; Otto, T.C.; Lane, M.D. Mitotic clonal expansion: A synchronous process required for adipogenesis. Proc. Natl. Acad. Sci. USA 2003, 100, 44–49. [Google Scholar] [CrossRef]
- Mohamed, G.A.; Ibrahim, S.R.; Elkhayat, E.S.; El Dine, R.S. Natural anti-obesity agents. Bull. Faculty Pharm. Cairo Univ. 2014, 52, 269–284. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, A.; Mukherjee, S.; Biswas, J.; Roy, M. Phytochemicals in obesity control. Int. J. Curr. Microbiol. App. Sci 2015, 4, 558–567. [Google Scholar]
- Yun, J.W. Possible anti-obesity therapeutics from nature—A review. Phytochemistry 2010, 71, 1625–1641. [Google Scholar] [CrossRef]
- Mukherjee, P.K.; Kumar, V.; Kumar, N.S.; Heinrich, M. The Ayurvedic medicine Clitoria ternatea--from traditional use to scientific assessment. J. Ethnopharmacol. 2008, 120, 291–301. [Google Scholar] [CrossRef]
- Terahara, N.; Oda, M.; Matsui, T.; Osajima, Y.; Saito, N.; Toki, K.; Honda, T. Five new anthocyanins, ternatins A3, B4, B3, B2, and D2, from Clitoria ternatea flowers. J. Nat. Prod. 1996, 59, 139–144. [Google Scholar] [CrossRef]
- Terahara, N.; Saito, N.; Honda, T.; Toki, K.; Osajima, Y. Structure of ternatin A1, the largest ternatin in the major blue anthocyanins from Clitoria ternatea flowers. Tetrahedron Lett. 1990, 31, 2921–2924. [Google Scholar] [CrossRef]
- Chayaratanasin, P.; Barbieri, M.A.; Suanpairintr, N.; Adisakwattana, S. Inhibitory effect of Clitoria ternatea flower petal extract on fructose-induced protein glycation and oxidation-dependent damages to albumin in vitro. BMC Complement. Altern. Med. 2015, 15, 27. [Google Scholar] [CrossRef] [PubMed]
- Gupta, G.K.; Chahal, J.; Bhatia, M. Clitoria ternatea (L.): Old and new aspects. J. Pharm. Res. 2010, 3, 2610–2614. [Google Scholar]
- Daisy, P.; Santosh, K.; Rajathi, M. Antihyperglycemic and antihyperlipidemic effects of Clitoria ternatea Linn. in alloxan-induced diabetic rats. Afr. J. Microbiol. Res. 2009, 3, 287–291. [Google Scholar]
- Daisy, P.; Rajathi, M. Hypoglycemic effects of Clitoria ternatea Linn.(Fabaceae) in alloxan-induced diabetes in rats. Trop. J. Pharm. Res. 2009, 8, 393–398. [Google Scholar] [CrossRef]
- Nair, V.; Bang, W.Y.; Schreckinger, E.; Andarwulan, N.; Cisneros-Zevallos, L. Protective role of ternatin anthocyanins and quercetin glycosides from Butterfly Pea (Clitoria ternatea Leguminosae) blue flower petals against lipopolysaccharide (LPS)-induced inflammation in macrophage Cells. J. Agric. Food Chem. 2015, 63, 6355–6365. [Google Scholar] [CrossRef]
- Shen, Y.; Du, L.; Zeng, H.; Zhang, X.; Prinyawiwatkul, W.; Alonso-Marenco, J.R.; Xu, Z. Butterfly pea (Clitoria ternatea) seed and petal extracts decreased HEp-2 carcinoma cell viability. Int. J. Food Sci. Technol. 2016, 51, 1860–1868. [Google Scholar] [CrossRef]
- Rayalam, S.; Della-Fera, M.A.; Baile, C.A. Phytochemicals and regulation of the adipocyte life cycle. J. Nutr. Biochem. 2008, 19, 717–726. [Google Scholar] [CrossRef]
- Patra, S.; Nithya, S.; Srinithya, B.; Meenakshi, S. Review of medicinal plants for anti-obesity activity. Transl. Biomed. 2015, 6, 21. [Google Scholar] [CrossRef]
- Kim, G.S.; Park, H.J.; Woo, J.H.; Kim, M.K.; Koh, P.O.; Min, W.; Ko, Y.G.; Kim, C.H.; Won, C.K.; Cho, J.H. Citrus aurantium flavonoids inhibit adipogenesis through the Akt signaling pathway in 3T3-L1 cells. BMC Complement. Altern. Med. 2012, 12, 31. [Google Scholar] [CrossRef]
- Song, Y.; Park, H.J.; Kang, S.N.; Jang, S.H.; Lee, S.J.; Ko, Y.G.; Kim, G.S.; Cho, J.H. Blueberry peel extracts inhibit adipogenesis in 3T3-L1 cells and reduce high-fat diet-induced obesity. PLoS ONE 2013, 8, e69925. [Google Scholar] [CrossRef]
- Xu, J.; Liao, K. Protein kinase B/AKT 1 plays a pivotal role in insulin-like growth factor-1 receptor signaling induced 3T3-L1 adipocyte differentiation. J. Biol. Chem. 2004, 279, 35914–35922. [Google Scholar] [CrossRef]
- Choi, K.M.; Lee, Y.S.; Sin, D.M.; Lee, S.; Lee, M.K.; Lee, Y.M.; Hong, J.T.; Yun, Y.P.; Yoo, H.S. Sulforaphane inhibits mitotic clonal expansion during adipogenesis through cell cycle arrest. Obesity 2012, 20, 1365–1371. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.L.; Lee, H.K.; Chin, T.Y.; Tu, S.C.; Kuo, M.H.; Kao, M.C.; Wu, Y.C. Inhibitory Effects of purple sweet potato leaf extract on the proliferation and lipogenesis of the 3T3-L1 Preadipocytes. Am. J. Chin. Med. 2015, 43, 915–925. [Google Scholar] [CrossRef] [PubMed]
- Fayard, E.; Xue, G.; Parcellier, A.; Bozulic, L.; Hemmings, B.A. Protein kinase B (PKB/Akt), a key mediator of the PI3K signaling pathway. Curr. Top. Microbiol. Immunol. 2010, 346, 31–56. [Google Scholar] [CrossRef]
- Tagaya, Y.; Miura, A.; Okada, S.; Ohshima, K.; Mori, M. Nucleobindin-2 is a positive modulator of EGF-dependent signals leading to enhancement of cell growth and suppression of adipocyte differentiation. Endocrinology 2012, 153, 3308–3319. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.S.; Kim, J.H.; Ali, M.Y.; Min, B.S.; Kim, G.D.; Jung, H.A. Coptis chinensis alkaloids exert anti- adipogenic activity on 3T3-L1 adipocytes by downregulating C/EBP-α and PPAR-γ. Fitoterapia 2014, 98, 199–208. [Google Scholar] [CrossRef]
- Lee, Y.J.; Choi, H.S.; Seo, M.J.; Jeon, H.J.; Kim, K.J.; Lee, B.Y. Kaempferol suppresses lipid accumulation by inhibiting early adipogenesis in 3T3-L1 cells and zebrafish. Food Funct. 2015, 6, 2824–2833. [Google Scholar] [CrossRef] [PubMed]
- Zebisch, K.; Voigt, V.; Wabitsch, M.; Brandsch, M. Protocol for effective differentiation of 3T3-L1 cells to adipocytes. Anal. Biochem. 2012, 425, 88–90. [Google Scholar] [CrossRef]
- Desvergne, B.; Michalik, L.; Wahli, W. Transcriptional regulation of metabolism. Physiol. Rev. 2006, 86, 465–514. [Google Scholar] [CrossRef]
- Schmid, B.; Rippmann, J.F.; Tadayyon, M.; Hamilton, B.S. Inhibition of fatty acid synthase prevents preadipocyte differentiation. Biochem. Biophys. Res. Commun. 2005, 328, 1073–1082. [Google Scholar] [CrossRef]
- Li, K.K.; Liu, C.L.; Shiu, H.T.; Wong, H.L.; Siu, W.S.; Zhang, C.; Han, X.Q.; Ye, C.X.; Leung, P.C.; Ko, C.H. Cocoa tea (Camellia ptilophylla) water extract inhibits adipocyte differentiation in mouse 3T3-L1 preadipocytes. Sci. Rep. 2016, 6, 20172. [Google Scholar] [CrossRef]
- Duncan, R.E.; Ahmadian, M.; Jaworski, K.; Sarkadi-Nagy, E.; Sul, H.S. Regulation of lipolysis in adipocytes. Annu. Rev. Nutr. 2007, 27, 79–101. [Google Scholar] [CrossRef]
- Lee, B.; Lee, M.; Lefevre, M.; Kim, H.R. Anthocyanins inhibit lipogenesis during adipocyte differentiation of 3T3-L1 preadipocytes. Plant. Foods Hum. Nutr. 2014, 69, 137–141. [Google Scholar] [CrossRef]
- Szkudelska, K.; Nogowski, L.; Szkudelski, T. Resveratrol, a naturally occurring diphenolic compound, affects lipogenesis, lipolysis and the antilipolytic action of insulin in isolated rat adipocytes. J. Steroid. Biochem. Mol. Biol. 2009, 113, 17–24. [Google Scholar] [CrossRef]
- Fernandez-Galilea, M.; Perez-Matute, P.; Prieto-Hontoria, P.L.; Martinez, J.A.; Moreno-Aliaga, M.J. Effects of lipoic acid on lipolysis in 3T3-L1 adipocytes. J. Lipid Res. 2012, 53, 2296–2306. [Google Scholar] [CrossRef]
- Kazuma, K.; Noda, N.; Suzuki, M. Flavonoid composition related to petal color in different lines of Clitoria ternatea. Phytochemistry 2003, 64, 1133–1139. [Google Scholar] [CrossRef]
- Bin, H.-S.; Choi, U.-K. Myricetin inhibits adipogenesis in human adipose tissue-derived mesenchymal stem cells. Food Sci. Biotechnol. 2012, 21, 1391–1396. [Google Scholar] [CrossRef]
- Wongs-Aree, C.; Giusti, M.; Schwartz, S. Anthocyanins derived only from delphinidin in the blue petals of Clitoria ternatea. In Proceedings of the IV International Conference on Managing Quality in Chains-The Integrated View on Fruits and Vegetables Quality, Bangkok, Thailand, 30 June 2006; pp. 437–442. [Google Scholar]
- Jang, Y.J.; Koo, H.J.; Sohn, E.H.; Kang, S.C.; Rhee, D.K.; Pyo, S. Theobromine inhibits differentiation of 3T3-L1 cells during the early stage of adipogenesis via AMPK and MAPK signaling pathways. Food Funct. 2015, 6, 2365–2374. [Google Scholar] [CrossRef]
- Galvis, A.; Marcano, A.; Stefancin, C.; Villaverde, N.; Priestap, H.A.; Tonn, C.E.; Lopez, L.A.; Barbieri, M.A. The effect of dehydroleucodine in adipocyte differentiation. Eur. J. Pharmacol. 2011, 671, 18–25. [Google Scholar] [CrossRef]
- Drira, R.; Chen, S.; Sakamoto, K. Oleuropein and hydroxytyrosol inhibit adipocyte differentiation in 3 T3-L1 cells. Life Sci. 2011, 89, 708–716. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.J.; Chen, C.Y.; Chang, G.D.; Wen, H.C.; Chen, C.Y.; Chang, S.C.; Liao, J.F.; Chang, C.H. Activation of Akt by advanced glycation end products (AGEs): Involvement of IGF-1 receptor and caveolin-1. PLoS ONE 2013, 8, e58100. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are available from the authors. |











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Rt (min) | Compound | Product Ion |
|---|---|---|---|
| 1 | 11.098 | Preternatin A3 | 1405.3310 [M + H]+, Calcd Mass: 1405.3876, MS/MS: 1329.2817, 1183.2193, 1021.2119, 876.1811, 739.1818, 627.1281, 493.3180, 471.3372 |
| 2 | 11.826 | Delphinidin derivatives | 788.4017 [M + H]+, Calcd Mass: 789.2084, MS/MS: 801.3825 [M + Na]+, [Quercetin 3-glucoside+dihexose] |
| 3 | 12.277 | Ternatin B2 | 1637.3591 [M + H]+, Calcd Mass: 1637.4248, MS/MS: 1389.3186, 757.1914, 611.1385, 465.0866, 303.0396 |
| 4 | 12.797 | Ternatin D2 | 1475.3163 [M + H]+, Calcd Mass: 1475.3720, MS/MS: 741.1967, 595.1441, 449.0916, 287.0456 |
| 5 | 13.040 | Quercetin-3-rutinoside (rutin) | 611.1384 [M + H]+, Calcd Mass: 610.1534, MS/MS: 465.0860, 303.0399 |
| 6 | 13.109 | Ternatin D1 | 1697.3959 [M + H − Malonyl]+, Calcd Mass: 1697.4612, MS/MS: 1389.3165, 1243.2461, 611.1379, 465.0855, 303.0396 |
| 7 | 13.664 | Kaemferol-3-O-(2-rhamnosyl)rutinoside | 741.1969 [M + H]+, Calcd Mass: 740.2164, MS/MS: 595.1443, 449.0924, 287.0457 |
| 8 | 14.080 | Delphinidin-3-glucoside (myrtillin) | 465.0856 [M + H]+, Calcd Mass: 465.1028, MS/MS: 487.0665 [M + Na]+, 303.0390 |
| 9 | 14.218 | Kaemferol-3-O-rutinoside | 595.1437 [M + H]+, Calcd Mass: 594.1585, MS/MS: 588.3886, 566.4074, 449.0917, 287.0454 |
| 10 | 14.738 | Delphinidin-3-O-(6-O-p-coumaryl)glucoside-pyruvic acid | 679.4869 [M + H]+, Calcd Mass: 679.1294, MS/MS: 701.4680 [M + Na]+, 595.1422, 340.2488 |
| 11 | 15.4311 | (+)-Catechin 7-O-β-glucoside | 453.3275 [M + H]+, Calcd Mass: 452.1319, MS/MS: 927.6276 [2M + Na]+, 905.6462 [2M + H]+, 814.5468 |
| 12 | 15.639 | Syringetin-3-O-glucoside | 509.8675 [M + H]+, Calcd Mass: 508.1217, MS/MS: 1040.7075 [2M + Na]+ |
| 13 | 16.402 | Quercetin triglycoside | 759.3755 [M + H]+, Calcd Mass: 758.1906, MS/MS: 781.3571 [M + Na]+ |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chayaratanasin, P.; Caobi, A.; Suparpprom, C.; Saenset, S.; Pasukamonset, P.; Suanpairintr, N.; Barbieri, M.A.; Adisakwattana, S. Clitoria ternatea Flower Petal Extract Inhibits Adipogenesis and Lipid Accumulation in 3T3-L1 Preadipocytes by Downregulating Adipogenic Gene Expression. Molecules 2019, 24, 1894. https://doi.org/10.3390/molecules24101894
Chayaratanasin P, Caobi A, Suparpprom C, Saenset S, Pasukamonset P, Suanpairintr N, Barbieri MA, Adisakwattana S. Clitoria ternatea Flower Petal Extract Inhibits Adipogenesis and Lipid Accumulation in 3T3-L1 Preadipocytes by Downregulating Adipogenic Gene Expression. Molecules. 2019; 24(10):1894. https://doi.org/10.3390/molecules24101894
Chicago/Turabian StyleChayaratanasin, Poramin, Allen Caobi, Chaturong Suparpprom, Sudarat Saenset, Porntip Pasukamonset, Nipattra Suanpairintr, Manuel Alejandro Barbieri, and Sirichai Adisakwattana. 2019. "Clitoria ternatea Flower Petal Extract Inhibits Adipogenesis and Lipid Accumulation in 3T3-L1 Preadipocytes by Downregulating Adipogenic Gene Expression" Molecules 24, no. 10: 1894. https://doi.org/10.3390/molecules24101894
APA StyleChayaratanasin, P., Caobi, A., Suparpprom, C., Saenset, S., Pasukamonset, P., Suanpairintr, N., Barbieri, M. A., & Adisakwattana, S. (2019). Clitoria ternatea Flower Petal Extract Inhibits Adipogenesis and Lipid Accumulation in 3T3-L1 Preadipocytes by Downregulating Adipogenic Gene Expression. Molecules, 24(10), 1894. https://doi.org/10.3390/molecules24101894