Rapid Identification of Tanshinone IIA Metabolites in an Amyloid-β1-42 Induced Alzherimer’s Disease Rat Model using UHPLC-Q-Exactive Qrbitrap Mass Spectrometry

Abstract
:1. Introduction
2. Results
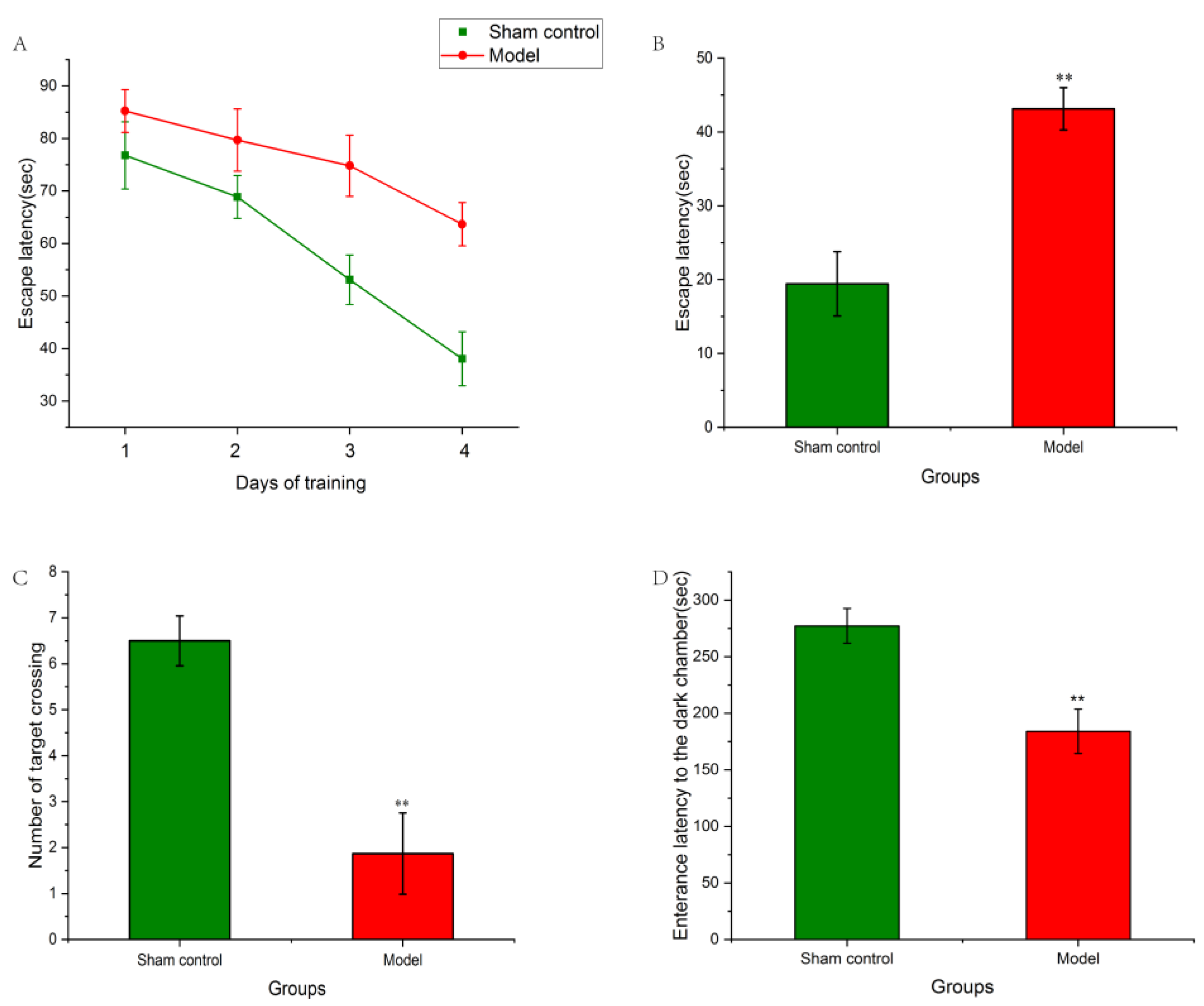
2.1. Effect of Aβ1–42 on Morris Water Maze Test of AD Rat Model
2.2. Effect of Aβ1–42 on Passive Avoidance Test AD Rat Model
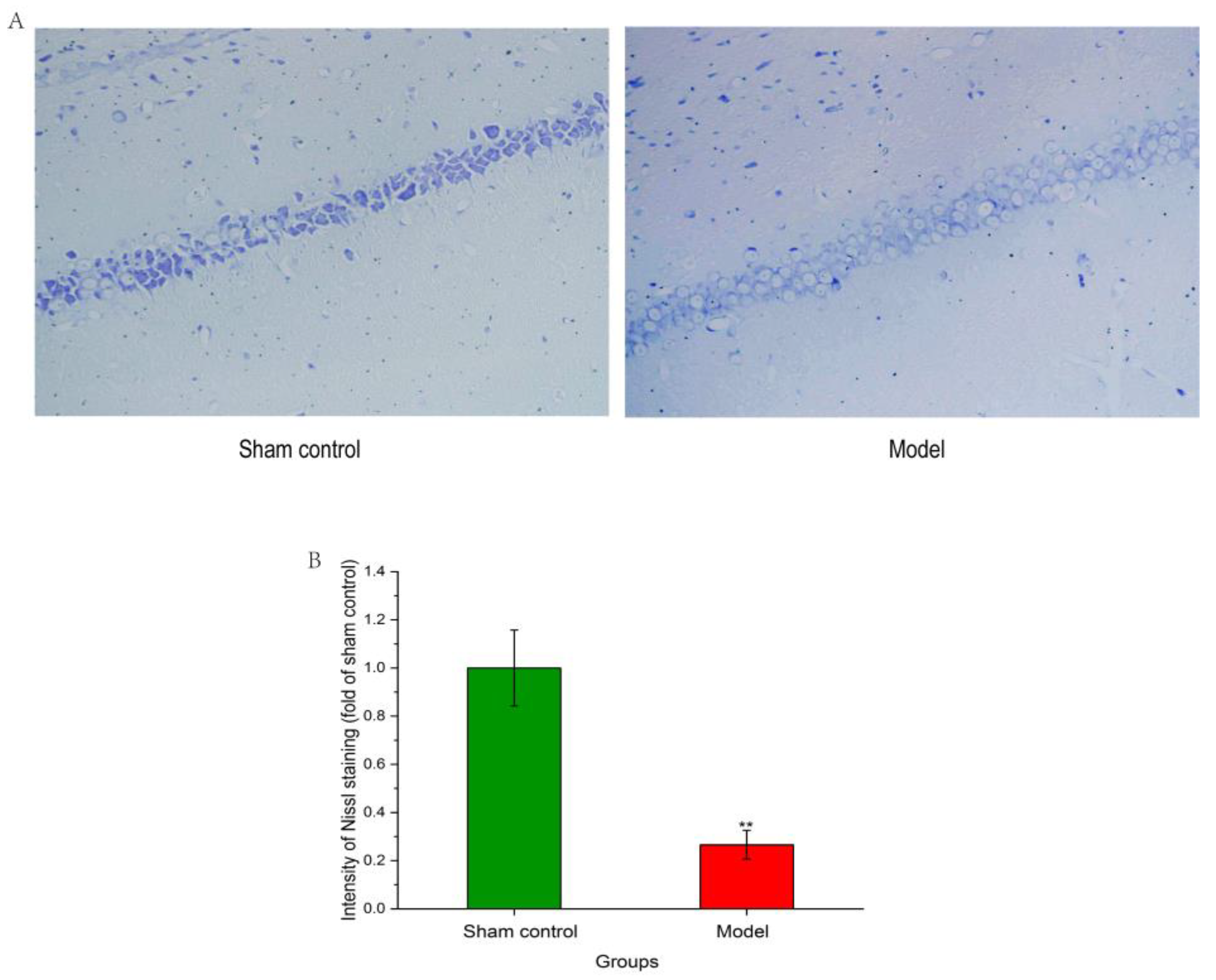
2.3. Effect of Aβ1–42 on Hippocampal Neuronal Structure of AD Rat Model
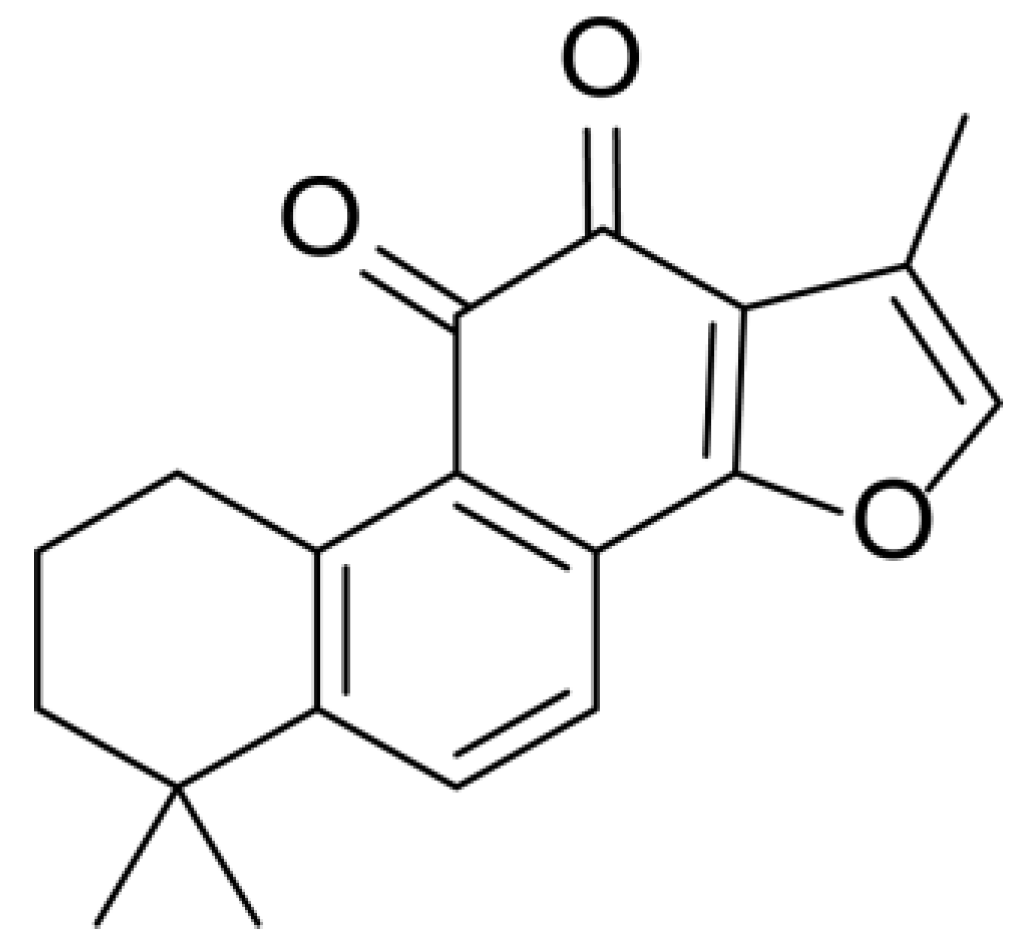
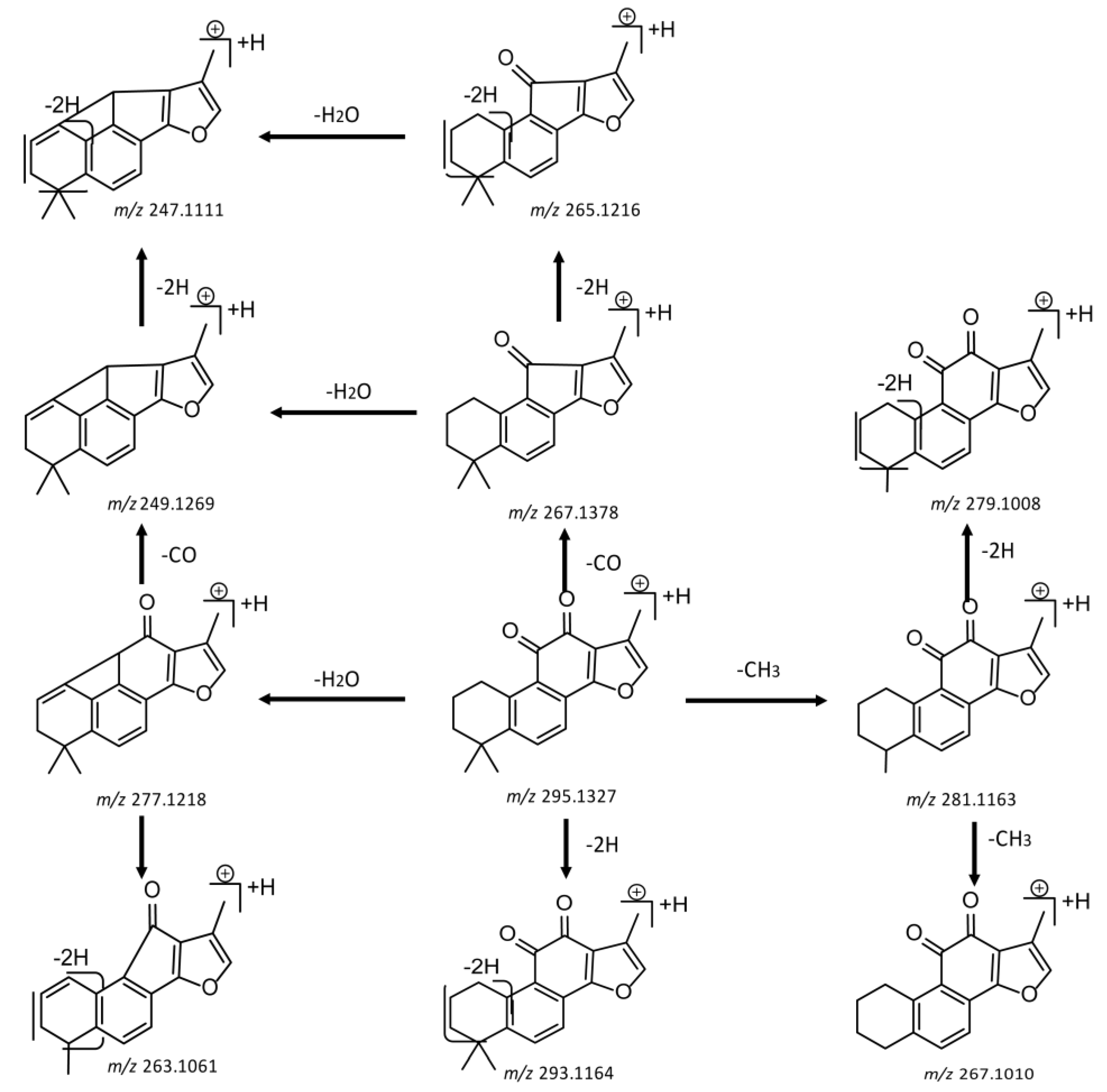
2.4. Mass Fragmentation Behavior Analyses of Tanshinone IIA
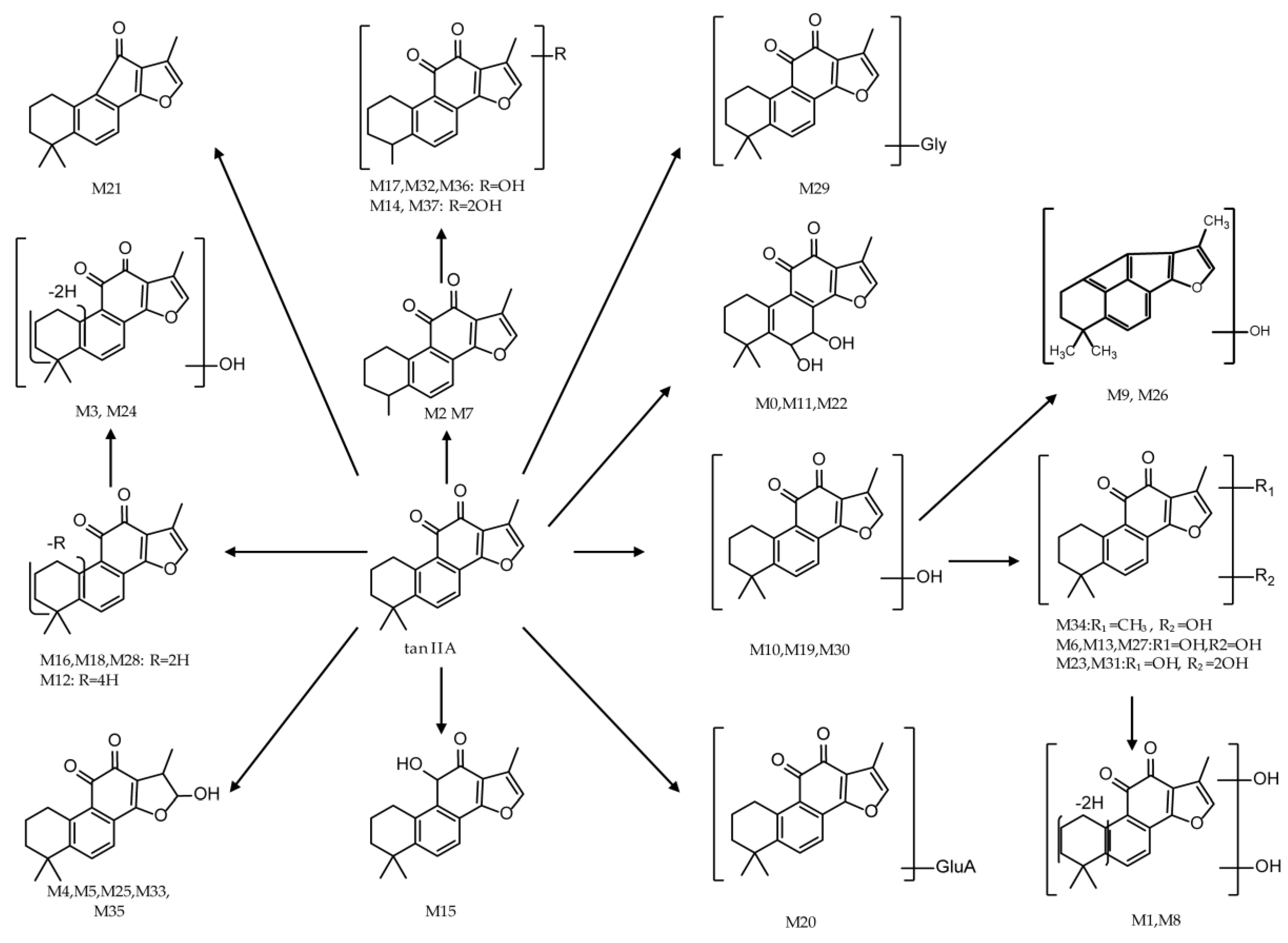
2.5. Identification of Metabolites of Tanshinone IIA in Rats
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Preparation of Aggregated Soluble Aβ1–42 Oligomers
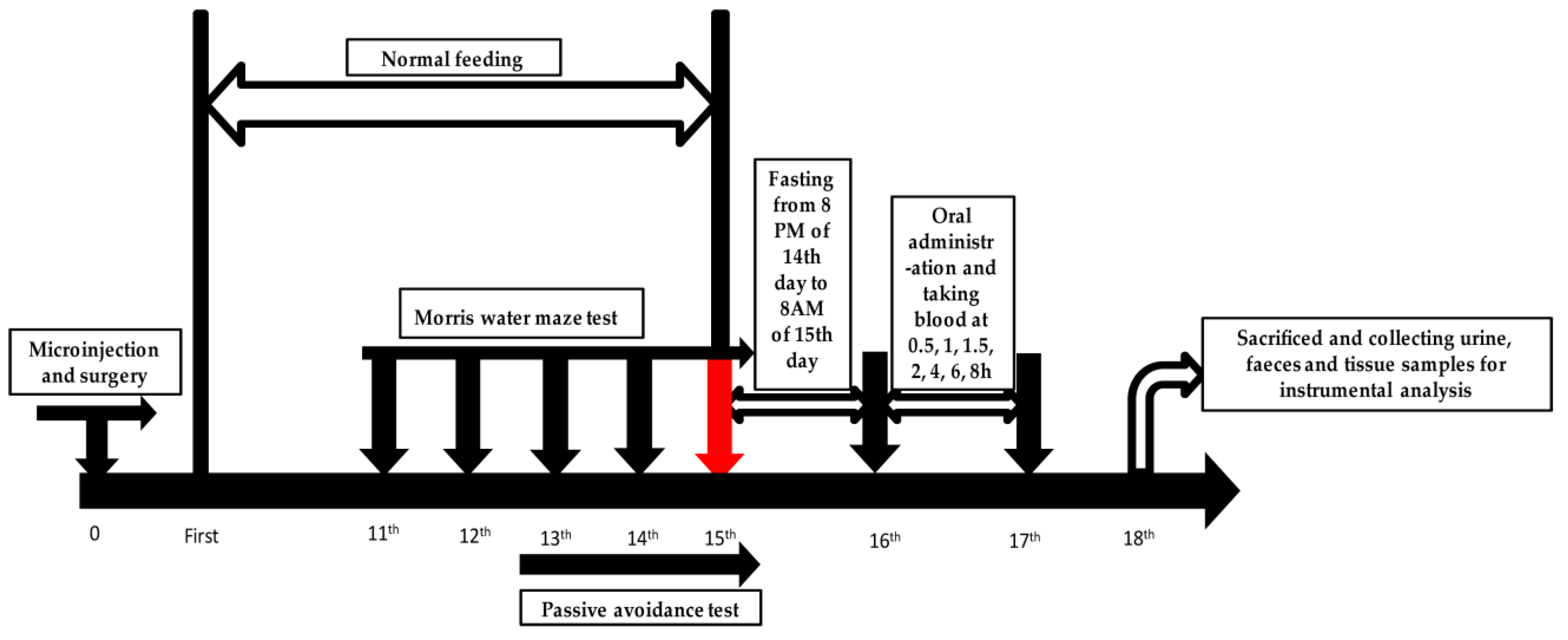
4.3. Animals and Drug Administration
4.4. Microinjection and Surgery
4.5. Morris Water Maze Test
4.6. Passive Avoidance Test
4.7. Sample Collection and Preparation
4.7.1. Plasma Sample Collection
4.7.2. Urine and Feces Sample Collection
4.7.3. Tissue Sample Collection
4.7.4. Sample Preparation
4.8. Nissl Staining
4.9. Instruments and Analytical Conditions
4.10. Data Processing and Statistical Analyses
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Koelsch, G. BACE1 Function and inhibition: Implications of intervention in the amyloid pathway of Alzheimer’s disease pathology. Molecules 2017, 22, 1723. [Google Scholar] [CrossRef] [PubMed]
- Yan, R. Stepping closer to treating Alzheimer’s disease patients with BACE1 inhibitor drugs. Transl. Neurodegener. 2016, 5, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, Q.Q.; Zhao, S.; Li, S.D.; Song, C. Application of chitosan, chitooligosaccharide, and their derivatives in the treatment of Alzheimer’s disease. Mar. Drugs 2017, 15, 322. [Google Scholar] [CrossRef] [PubMed]
- Baldacci, F.; Lista, S.; Cavedo, E.; Bonuccelli, U.; Hampel, H. Diagnostic function of the neuroinflammatory biomarker YKL-40 in Alzheimer’s disease and other neurodegenerative diseases. Expert Rev. Proteom. 2017, 14, 285–299. [Google Scholar] [CrossRef] [PubMed]
- Heneka, M.T.; Carson, M.J.; El Khoury, J.; Landreth, G.E.; Brosseron, F.; Feinstein, D.L.; Jacobs, A.H.; Wyss-Coray, T.; Vitorica, J.; Ransohoff, R.M.; et al. Neuroinflammation in Alzheimer’s disease. Lancet Neurol. 2015, 14, 388–405. [Google Scholar] [CrossRef]
- Monte, S.M.D.L.; Tong, M. Brain metabolic dysfunction at the core of Alzheimer’s disease. Biochem. Pharmacol. 2014, 88, 548–559. [Google Scholar] [CrossRef] [PubMed]
- Olsson, B.; Lautner, R.; Andreasson, U.; Öhrfelt, A.; Portelius, E.; Bjerke, M.; Höltt, M.; Rosén, C.; Olsson, C.; Strobel, G.; et al. CSF and blood biomarkers for the diagnosis of Alzheimer’s disease: A systematic review and meta-analysis. Lancet Neurol. 2016, 15, 673–684. [Google Scholar] [CrossRef]
- Lista, S.; Hampel, H. Synaptic degeneration and neurogranin in the pathophysiology of Alzheimer’s disease. Expert Rev. Neurother. 2017, 17, 47. [Google Scholar] [CrossRef]
- Iturria-Medina, Y.; Sotero, R.C.; Toussaint, P.J.; Mateos-Pérez, J.M.; Evans, A.C.; Weiner, M.W.; Aisen, P.; Petersen, R.; Jack, C.R.; Jagust, W. Early role of vascular dysregulation on late-onset Alzheimer’s disease based on multifactorial data-driven analysis. Nat. Commun. 2016, 7, 11934–11948. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Hou, J.; Ping, J.; Cai, D. Advances in developing novel therapeutic strategies for Alzheimer’s disease. Mol. Neurodegener. 2018, 13, 64–84. [Google Scholar] [CrossRef] [PubMed]
- Evin, G. Future Therapeutics in Alzheimer’s disease: Development status of BACE inhibitors. BioDrugs 2016, 30, 173–194. [Google Scholar] [CrossRef]
- Chen, X.Y.; Du, Y.F.; Chen, L. Neuropeptides exert neuroprotective effects in Alzheimer’s disease. Front. Mol. Neurosci. 2018, 11, 493–506. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Dominguez, R.; Garcia-Barrera, T.; Gomez-Ariza, J.L. Metabolomic study of lipids in serum for biomarker discovery in Alzheimer’s disease using direct infusion mass spectrometry. J. Pharm. Biomed. Anal. 2014, 98, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Zhang, W.; Chen, Z.; Shi, Z.; He, C.; Chen, M. Recent insights into the biological activities and drug delivery systems of tanshinones. Int. J. Nanomed. 2016, 11, 121–130. [Google Scholar] [CrossRef]
- Zhu, S.; Wei, W.; Liu, Z.; Yang, Y.; Jia, H. TanshinoneIIA attenuates the deleterious effects of oxidative stress in osteoporosis through the NFkappaB signaling pathway. Mol. Med. Rep. 2018, 17, 6969–6976. [Google Scholar] [CrossRef]
- Gu, Y.; Liang, Z.; Wang, H.; Jin, J.; Zhang, S.; Xue, S.; Chen, J.; He, H.; Duan, K.; Wang, J.; et al. Tanshinone IIA protects H9c2 cells from oxidative stress-induced cell death via microRNA-133 upregulation and Akt activation. Exp. Ther. Med. 2016, 12, 1147–1152. [Google Scholar] [CrossRef] [Green Version]
- Lv, C.; Zeng, H.W.; Wang, J.X.; Yuan, X.; Zhang, C.; Fang, T.; Yang, P.M.; Wu, T.; Zhou, Y.D.; Nagle, D.G.; et al. The antitumor natural product tanshinone IIA inhibits protein kinase C and acts synergistically with 17-AAG. Cell Death Dis. 2018, 9, 165–178. [Google Scholar] [CrossRef]
- Chiu, C.M.; Huang, S.Y.; Chang, S.F.; Liao, K.F.; Chiu, S.C. Synergistic antitumor effects of tanshinone IIA and sorafenib or its derivative SC-1 in hepatocellular carcinoma cells. Onco Targets Ther. 2018, 11, 1777–1785. [Google Scholar] [CrossRef]
- Gao, H.; Huang, L.; Ding, F.; Yang, K.; Feng, Y.; Tang, H.; Xu, Q.M.; Feng, J.; Yang, S. Simultaneous purification of dihydrotanshinone, tanshinone I, cryptotanshinone, and tanshinone IIA from Salvia miltiorrhiza and their anti-inflammatory activities investigation. Sci. Rep. 2018, 8, 8460–8473. [Google Scholar] [CrossRef]
- Tang, J.; Zhou, S.; Zhou, F.; Wen, X. Inhibitory effect of tanshinone IIA on inflammatory response in rheumatoid arthritis through regulating beta-arrestin 2. Exp. Ther. Med. 2019, 17, 3299–3306. [Google Scholar] [CrossRef]
- Li, H.; Han, W.; Wang, H.; Ding, F.; Xiao, L.; Shi, R.; Ai, L.; Huang, Z. Tanshinone IIA inhibits glutamate-induced oxidative toxicity through prevention of mitochondrial dysfunction and suppression of MAPK activation in SH-SY5Y human neuroblastoma cells. Oxid. Med. Cell. Longev. 2017, 2017, 4517486–4517499. [Google Scholar] [CrossRef] [PubMed]
- Qiuming, W.; Xiang, Y.; Kunal, P.; Rundong, H.; Steven, C.; Ge, Z.; Jie, Z. Tanshinones inhibit amyloid aggregation by amyloid-β peptide, disaggregate amyloid fibrils, and protect cultured cells. ACS Chem. Neurosci. 2013, 4, 1004–1015. [Google Scholar] [CrossRef]
- Shi, C.; Zhu, X.; Wang, J.; Long, D. Tanshinone IIA promotes non-amyloidogenic processing of amyloid precursor protein in platelets via estrogen receptor signaling to phosphatidylinositol 3-kinase/Akt. Biomed. Rep. 2014, 2, 500–504. [Google Scholar] [CrossRef]
- Lin, L.; Jadoon, S.S.; Liu, S.Z.; Zhang, R.Y.; Li, F.; Zhang, M.Y.; Ai-Hua, T.; You, Q.Y.; Wang, P. Tanshinone IIA ameliorates spatial learning and memory deficits by inhibiting the activity of ERK and GSK-3beta. J. Geriatr. Psychiatry Neurol. 2019, 32, 152–163. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Wang, G.J.; Li, J.; Hao, H.P.; Zheng, C.N. Identification of tanshinone IIA metabolites in rat liver microsomes by liquid chromatography–tandem mass spectrometry. J. Chromatogr. A 2006, 1104, 366–369. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.H.; Yang, M.; Han, J.; Wang, B.R.; Ma, X.C.; Xu, M.; Liu, P.; Guo, D. Profiling the metabolic difference of seven tanshinones using high-performance liquid chromatography/multi-stage mass spectrometry with data-dependent acquisition. Rapid Commun. Mass Spectrom. 2010, 21, 2211–2226. [Google Scholar] [CrossRef]
- Wei, Y.J.; Li, P.; Wang, C.M.; Peng, Y.R.; Shu, L.; Jia, X.B.; Ma, W.Q.; Wang, B. Metabolism of tanshinone IIA, cryptotanshinone and tanshinone I from Radix Salvia miltiorrhiza in zebrafish. Molecules 2012, 17, 8617–8632. [Google Scholar] [CrossRef]
- Li, J.; Kuang, G.; Chen, X.; Zeng, R. Identification of chemical composition of leaves and flowers from paeonia rockii by UHPLC-Q-Exactive Orbitrap HRMS. Molecules 2016, 21, 947. [Google Scholar] [CrossRef]
- Senyuva, H.Z.; Gökmen, V.; Sarikaya, E.A. Future perspectives in Orbitrap™-high-resolution mass spectrometry in food analysis: A review. Food Addit. Contam. 2015, 32, 1568–1606. [Google Scholar] [CrossRef]
- Li, L.; Wang, Y.; Xiu, Y.; Liu, S. Chemical differentiation and quantitative analysis of different types of panax genus stem-leaf based on a UPLC-Q-Exactive Orbitrap/MS combined with multivariate statistical analysis approach. J. Anal. Methods Chem. 2018, 2018, 9598672–9598688. [Google Scholar] [CrossRef]
- Michalski, A.; Damoc, E.; Hauschild, J.P.; Lange, O.; Wieghaus, A.; Makarov, A.; Nagaraj, N.; Cox, J.; Mann, M.; Horning, S. Mass spectrometry-based proteomics using Q Exactive, a high-performance benchtop quadrupole Orbitrap mass spectrometer. Mol. Cell. Proteom. 2011, 10, M111. [Google Scholar] [CrossRef]
- Li, Q.; Zhao, Y.; Zhu, D.; Pang, X.; Liu, Y.; Frew, R.; Chen, G. Lipidomics profiling of goat milk, soymilk and bovine milk by UPLC-Q-Exactive Orbitrap mass spectrometry. Food Chem. 2017, 224, 302–309. [Google Scholar] [CrossRef] [PubMed]
- Morris, R.G.M. Spatial localization does not require the presence of local cues. Learn. Motiv. 1981, 12, 239–260. [Google Scholar] [CrossRef]
- D’Hooge, R.; Deyn, P.P.D. Applications of the Morris water maze in the study of learning and memory. Brain Res. Brain Res. Rev. 2001, 36, 60–90. [Google Scholar] [CrossRef]
- Jiang, J.; Li, Z.; Kai, G.; Yuan, Z.; Han, X.; Zhanxun, P. Electro-acupuncture treatment improves learning-memory ability and brain glucose metabolism in a mouse model of Alzheimer’s disease:from MWM and m-PET. Evid. Based Complement. Altern. Med. 2015, 2015, 142129–142136. [Google Scholar] [CrossRef] [PubMed]
- Meilin, S.; Machicao, F.; Elmlinger, M. Treatment with Actovegin improves spatial learning and memory in rats following transient forebrain ischaemia. J. Cell. Mol. Med. 2014, 18, 1623–1630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Engen, J.R.; Hobbins, W.B. Ultra performance liquid chromatography (UPLC) further improves hydrogen/deuterium exchange mass spectrometry. J. Am. Soc. Mass Spectrom. 2006, 17, 163–167. [Google Scholar] [CrossRef] [Green Version]
- Jia, S.; Lu, Z.; Gao, Z.; An, J.; Wu, X.; Li, X.; Dai, X.; Zheng, Q.; Sun, Y. Chitosan oligosaccharides alleviate cognitive deficits in an amyloid-β 1–42 -induced rat model of Alzheimer’s disease. Int. J. Biol. Macromol. 2016, 83, 416–425. [Google Scholar] [CrossRef]
- Yu, Y.; Zhou, L.; Sun, M.; Zhou, T.; Zhong, K.; Wang, H.; Liu, Y.; Liu, X.; Xiao, R.; Ge, J.; et al. Xylocoside G reduces amyloid-beta induced neurotoxicity by inhibiting NF-kappaB signaling pathway in neuronal cells. J. Alzheimer Dis. 2012, 30, 263–275. [Google Scholar] [CrossRef]
- Ghosh, A.K.; Osswald, H.L. BACE1 (β-secretase) inhibitors for the treatment of Alzheimer’s disease. Chem. Soc. Rev. 2014, 43, 6765–6813. [Google Scholar] [CrossRef]
- Zhang, L.; Fang, Y.; Xu, Y.; Lian, Y.; Xie, N.; Wu, T.; Zhang, H.; Sun, L.; Zhang, R.; Wang, Z. Curcumin improves amyloid β-Peptide (1-42) induced spatial memory deficits through BDNF-ERK signaling pathway. PLoS ONE 2015, 10, e0131525–e0131542. [Google Scholar] [CrossRef] [PubMed]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 7th ed.; Academic Press: Cambridge, MA, USA, 2013; pp. 1–472. ISBN 978-0-12-391949-6. [Google Scholar]
- Vorhees, C.V.; Williams, M.T. Morris water maze: Procedures for assessing spatial and related forms of learning and memory. Nat. Protoc. 2006, 1, 848–858. [Google Scholar] [CrossRef] [PubMed]
- Shibani, F.; Sahamsizadeh, A.; Fatemi, I.; Allahtavakoli, M.; Hasanshahi, J.; Rahmani, M.; Azin, M.; Hassanipour, M.; Mozafari, N.; Kaeidi, A. Effect of oleuropein on morphine-induced hippocampus neurotoxicity and memory impairments in rats. Naunyn Schmiedeberg Arch. Pharmacol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Khorasani, A.; Abbasnejad, M.; Esmaeili-Mahani, S. Phytohormone abscisic acid ameliorates cognitive impairments in streptozotocin-induced rat model of Alzheimer’s disease through PPARbeta/delta and PKA signaling. Int. J. Neurosci. 2019. [Google Scholar] [CrossRef] [PubMed]
- Garabadu, D.; Verma, J. Exendin-4 attenuates brain mitochondrial toxicity through PI3K/Akt-dependent pathway in amyloid beta (1-42)-induced cognitive deficit rats. Neurochem. Int. 2019, 128, 39–49. [Google Scholar] [CrossRef]
- Mana, L.; Feng, H.; Dong, Y.; Wang, Y.; Shi, J.; Tian, J.; Wang, P. Effect of Chinese herbal compound GAPT on the early brain glucose metabolism of APP/PS1 transgenic mice. Int. J. Immunopathol. Pharmacol. 2019, 33, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Bromley-Brits, K.; Deng, Y.; Song, W. Morris water maze test for learning and memory deficits in Alzheimer’s disease model mice. J. Vis. Exp. 2011, 53, e2920–e2925. [Google Scholar] [CrossRef]
- Tian, F.Z.; Chang, H.S.; Liu, J.X.; Zheng, J.; Cheng, D.; Lu, Y. Cynomorium songaricum extract alleviates memory impairment through increasing CREB/BDNF via suppression of p38MAPK/ERK pathway in ovariectomized rats. Evid. Based Complement. Altern. Med. 2019, 2019, 9689325–9689335. [Google Scholar] [CrossRef]
- Staay, F.J.V.D.; Schuurman, T.; Reenen, C.G.V.; Korte, S.M. Emotional reactivity and cognitive performance in aversively motivated tasks: A comparison between four rat strains. Behav. Brain Funct. 2009, 5, 50–79. [Google Scholar] [CrossRef]
- Shiga, T.; Nakamura, T.J.; Komine, C.; Goto, Y.; Mizoguchi, Y.; Yoshida, M.; Kondo, Y.; Kawaguchi, M. A single neonatal injection of ethinyl estradiol impairs passive avoidance learning and reduces expression of estrogen receptor α in the hippocampus and cortex of adult female rats. PLoS ONE 2016, 11, e0146136–e0146147. [Google Scholar] [CrossRef]
- Ji, C.; Aisa, H.A.; Yang, N.; Li, Q.; Wang, T.; Zhang, L.; Qu, K.; Zhu, H.B.; Zuo, P.P. Gossypium herbaceam extracts inhibited NF-kappaB activation to attenuate spatial memory impairment and hippocampal neurodegeneration induced by amyloid-beta in rats. J. Alzheimer Dis. 2008, 14, 271–283. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak | tR/min | Formula [M + H]− | Theoretical Mass m/z | Experimental Mass m/z | Error (ppm) | MS/MS Fragment Ions | Identification/Reactions | Distribution | Sham Group | Model Group |
|---|---|---|---|---|---|---|---|---|---|---|
| M0 | 4.25 | C19H21O5 | 329.1383 | 329.13809 | −0.791 | MS2[329]:283.1321(100),311.1268(50),265.1219(15),267.1371(11) |  | 2 | + | + |
| M1 | 4.36 | C19H17O5 | 325.107 | 325.10681 | −0.739 | MS2[325]:156.0764(100),110.0715(59),307.0952(26),279.1008(23),168.1015(22) |  | 1 | + | + |
| M2 | 4.75 | C18H17O3 | 281.1172 | 281.11703 | −0.679 | MS2[281];133.0645(100),263.1061(18),282.18024(12),281.1166(10) |  | 1 | + | |
| M3 | 4.81 | C19H17O4 | 309.1121 | 309.11163 | −3.409 | MS2[309]:204.1378(100),267.1695(12) |  | 1 | + | |
| M4 | 5.71 | C19H21O4 | 313.1435 | 313.14383 | 1.259 | MS2[313]:85.0289(100),313.2163(18),267.1378(13),295.2051(12) |  | 1 | + | |
| M5 | 5.72 | C19H21O4 | 313.1435 | 313.14368 | 0.78 | MS2[313]:313.2150(100),159.1167(54), 295.2049(53.32), 277.1943(35),314.2175(26) |  | 7 | + | |
| M6 | 6.2 | C19H19O5 | 327.1227 | 327.1226 | −0.062 | MS2[327]:327.1219(100),309.1113(69),281.1166(61),257.1166(40),328.2262(26) |  | 1,2 | + | + |
| M7 | 6.22 | C18H17O3 | 281.1172 | 281.11716 | −0.217 | MS2[281]:255.1009(100),149.0230(72),224.2004(47),263.1636(42),207.1012(36),279.1578(36),95.0860(35),81.0705(31) |  | 1 | + | + |
| M8 | 6.6 | C19H17O5 | 325.107 | 325.10724 | 0.584 | MS2[325]:325.1060(100),281.1164(21) |  | 2 | + | + |
| M9 | 6.84 | C18H17O2 | 265.1223 | 265.12241 | 0.391 | MS2[265]:247.1685(100),187.1476(51),191.1059(27),229.1576(23) |  | 1 | + | + |
| M10 | 6.85 | C19H19O4 | 311.1278 | 311.12766 | −0.404 | MS2[311]:265.1216(100),283.1315(46),266.1237(20),223.0749(15),311.1257(13),223.1116(10) |  | 1 | + | |
| M11 | 7.45 | C19H21O5 | 329.1383 | 329.1385 | 0.607 | MS2[329]:85.0653(100),311.2002(57),330.2246(41),109.1014(44) |  | 1 | + | + |
| M12 | 8.33 | C19H15O3 | 291.1015 | 291.10138 | −2.539 | MS2[291]:290.1094(100),292.1068(43),249.1576(24),273.1848(14) |  | 1,2,3,4,5,6 | + | |
| M13 | 8.7 | C19H19O5 | 327.1227 | 327.12234 | −2.777 | MS2[327]:281.1163(100),309.1108(99),327.1212(96),273.1114(40),256.1088(29),268.1200(18) |  | 1 | + | + |
| M14 | 8.75 | C18H17O5 | 313.107 | 313.10617 | 1.477 | MS2[313]:285.1116(100),313.1061(63),267.1010(37),295.0950(22),163.1107(13) |  | 2 | + | + |
| M15 | 9.01 | C19H21O3 | 297.1486 | 297.14819 | −1.114 | MS2[297]:297.1117(100),279.1011(18),251.1056(13),269.1168(11) |  | 2 | + | |
| M16 | 9.13 | C19H17O3 | 293.1172 | 293.11688 | −1.163 | MS2[293]:293.11655(100),275.1055(87),247.1115(37) |  | 1 | + | + |
| M17 | 9.14 | C18H17O4 | 297.1121 | 297.11234 | 0.688 | MS2[297]:297.1117(100),95.0859(27),226.6350(20),251.1056(13),269.1168(10) |  | 2 | + | |
| M18 | 9.27 | C19H17O3 | 293.1172 | 293.11694 | −2.829 | MS2[293]:293.1160(100),226.89243(51),275.1056(32) |  | 1 | + | + |
| M19 | 9.27 | C19H19O4 | 311.1278 | 311.1275 | −0.918 | MS2[311]:275.1058(100),293.1164(72),196.1691(40),311.1268(39),265.1217(33),283.1163(12), |  | 1,2,3,4,5 | + | + |
| M20 | 9.46 | C25H27O9 | 471.165 | 471.16409 | −1.844 | MS2[471]:295.1321(100),277.1219(19) |  | 1 | + | |
| M21 | 9.6 | C18H19O2 | 267.138 | 267.13748 | −1.783 | MS2[267]:267.1369(100),225.1957(42),238.9165(18),211.06177(14) |  | 2 | + | + |
| M22 | 9.61 | C19H21O5 | 329.1383 | 329.13785 | −1.52 | MS2[329]:329.1373(100),299.0907(91),311.1269(39),283.1310(12),109.1012(12) |  | 1 | + | + |
| M23 | 9.61 | C19H19O6 | 343.1176 | 343.11746 | −0.451 | MS2[343]:279.1009(100),283.0957(63),325.1058(46),343.1167(31),297.1111(21),253.0853(17) |  | 1 | + | |
| M24 | 9.76 | C19H17O4 | 309.1121 | 309.1116 | −3.507 | MS2[309]:265.1217(100),309.1113(82) |  | 2 | + | |
| M25 | 9.87 | C19H21O4 | 313.1435 | 313.14352 | 0.269 | MS2[313]:84.9603(100),267.9286(58),313.1438(34),219.9089(14),238.9163(10) |  | 1,2 | + | + |
| M26 | 10.28 | C18H17O2 | 265.1223 | 265.12225 | −0.212 | MS2[265]:265.1215(100),201.0485(30),210.0539(20) |  | 2 | + | |
| M27 | 10.3 | C19H19O5 | 327.1227 | 327.12253 | −0.52 | MS2[327]:281.1165(100),309.1111(32),257.1163(14) |  | 2 | + | |
| M28 | 10.62 | C19H17O3 | 293.1172 | 293.11682 | −3.239 | MS2[293]:293.1165(100),275.1059(31),247.1111(25),278.0929(13) |  | 1 | + | + |
| M29 | 10.7 | C21H22O4N | 352.1544 | 352.15411 | −0.638 | MS2[352]:333.1089(100),352.1579(39),310.1425(38),210.1493(12),283.1340(10) |  | 1,2,5 | + | |
| M30 | 11.28 | C19H19O4 | 311.1278 | 311.12796 | 0.561 | MS2[311]:293.1163(100),311.1261(39),275.1052(25),247.1188(24),265.1222(17),286.6728(15),283.1318(10) |  | 2 | + | |
| M31 | 11.48 | C19H19O6 | 343.1176 | 343.11761 | -0.014 | MS2[343]:323.2511(100),341.2621(56),325.1058(44),343.1141(26),297.1100(20) |  | 1 | + | |
| M32 | 11.58 | C18H17O4 | 297.1121 | 297.11255 | 1.395 | MS2[297]:297.1112(100),81.0704(24),279.1004(19),97.1015(11) |  | 2 | + | + |
| M33 | 11.93 | C19H21O4 | 313.1435 | 313.14343 | −0.018 | MS2[313]:313.1421(100),271.1320(32),297.1496(28),267.1379(25),295.1332(17),107.0859(11) |  | 2 | + | + |
| M34 | 12.37 | C20H21O4 | 325.1434 | 325.14365 | 0.659 | MS2[325]:325.1423(100),265.1214(43),209.1167(41),279.1377(18) |  | 2 | + | |
| M35 | 13.09 | C19H21O4 | 313.1435 | 313.14322 | −0.689 | MS2[313]:313.1419(100),295.1322(39),267.1371(18),83.96021(11) |  | 2 | + | + |
| M36 | 13.53 | C18H17O4 | 297.1121 | 297.11194 | −2.504 | MS2[297]:269.1164(100),287.1276(21),297.1116(20),270.1204(19),109.1012(12) |  | 1 | + | + |
| M37 | 13.68 | C18H17O5 | 313.107 | 313.10715 | 0.319 | MS2[313]:285.1117(100),269.1169(85),313.1065(58),267.10065(41)295.0972(26) |  | 2 | + | + |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, S.; Wang, Z.; Yuan, J.; Zhang, J.; Dai, X.; Qin, F.; Zhang, J.; Sun, Y. Rapid Identification of Tanshinone IIA Metabolites in an Amyloid-β1-42 Induced Alzherimer’s Disease Rat Model using UHPLC-Q-Exactive Qrbitrap Mass Spectrometry. Molecules 2019, 24, 2584. https://doi.org/10.3390/molecules24142584
Liang S, Wang Z, Yuan J, Zhang J, Dai X, Qin F, Zhang J, Sun Y. Rapid Identification of Tanshinone IIA Metabolites in an Amyloid-β1-42 Induced Alzherimer’s Disease Rat Model using UHPLC-Q-Exactive Qrbitrap Mass Spectrometry. Molecules. 2019; 24(14):2584. https://doi.org/10.3390/molecules24142584
Chicago/Turabian StyleLiang, Shuang, Zijian Wang, Jiaqi Yuan, Jing Zhang, Xueling Dai, Fei Qin, Jiayu Zhang, and Yaxuan Sun. 2019. "Rapid Identification of Tanshinone IIA Metabolites in an Amyloid-β1-42 Induced Alzherimer’s Disease Rat Model using UHPLC-Q-Exactive Qrbitrap Mass Spectrometry" Molecules 24, no. 14: 2584. https://doi.org/10.3390/molecules24142584
APA StyleLiang, S., Wang, Z., Yuan, J., Zhang, J., Dai, X., Qin, F., Zhang, J., & Sun, Y. (2019). Rapid Identification of Tanshinone IIA Metabolites in an Amyloid-β1-42 Induced Alzherimer’s Disease Rat Model using UHPLC-Q-Exactive Qrbitrap Mass Spectrometry. Molecules, 24(14), 2584. https://doi.org/10.3390/molecules24142584